

ipatov1997_fitotsenologija
.pdfРис. 6.37. Зависимость обилия видов от глубины снежного покрова (Л) (по: Александрова. 1961) и степень проникновения через последний солнечной радиации (Б) (по: Кузьмин, 1957).
По оси абсцисс — глубина снежного покрова, по оси ординат (Б) — степень проникновения через снег солнечной радиации для снега с ко-
эффицентом прозрачности: а — 0.65, 6 — 0.75, в — 0.80, г — 0.85, д — 0.90. 1
— Potentilla emarginata, 2— Alopecurus alpinus, 3— Papaver polare, 4 — Draba. pohlei, 5— Ranunculus nivalis, 6— Cerastium bialynickii, 7 — Salix polaris, 8 — Deschampsia brevifolia, 9— Androsace triflora, 10 — Juncus biglumis, 11— Phipps%a algida, 12— Ranunculus pygmaeus.
ванности среды, естественно, это может быть одной из причин квантованности растительного покрова.
На переломные пункты в изменении прямо действующих экологических режимов как источник дискретности указала В. Д. Александрова (1965), при этом она опиралась на пример влияния снежного покрова на растительность арктической тундры (рис.6.37). На Новосибирских островах при увеличении толщины снежного покрова более 12-13 см наблюдается смена полигональной тундры с преобладанием подушковидных форм разнотравья кочковато-пятнистой с господством Salix polaris, Alopecurus alpmus и др. При глубине снега менее указанной мощности солнечная радиация нагревает подстилающую поверхность, вызывая образование проталин задолго до того, как начинает освобождаться от льда остальная поверхность тундры. Если глубина снега превышает 12-13 см, этот эффект не наблюдается. Таким образом, при постепенном изменении косвенно действующий фактор достигает критических значений, при кото-
229
рых происходит скачкообразное изменение прямодействующих факторов. Снежный покров выступает как фактор формирования экотопа, т.е. само явление вызвано первой, указанной ранее причиной квантованности.
Следующая причина квантованности— специфическая трансформация среды растениями. Этот вопрос уже рассмотрен ранее, подчеркнем только некоторые аспекты роли эдификаторов в дифференциации растительного покрова. Эдификаторная роль растений как причина дискретности признается практически всеми исследователями, независимо от их позиции в отношении континуума растительности. Растения разных жизненных форм неодинаковы в отношении эдификаторной способности. Видимо, все или почти все деревья и кустарники могут выступать в роли эдификаторов, причем как на уровне особей, так и коллективов. Эдификаторная роль травянистых растений изучена довольно слабо. Может быть, поэтому распространено мнение, что среди трав эдификаторами выступают только некоторые виды и преимущественно на уровне коллективов. Размеры квантов, создаваемых эдификаторами, соответствуют размерам растений (их совокупностей) и, естественно, в травянистых типах много меньше. Конечно, размеры квантов зависят и от того, кто является эдификатором— особь или группы особей разных размеров. Эдификаторы могут создавать квантованность на уровнях как растительного покрова в целом, так и ценоэлементов. Равномерно представленный на некоторой территории эдификатор, выравнивая среду, может приводить к снижению квантованности подчиненных элементов в пределах этой территории. Вместе с тем, формируя растительный покров, отличный от покрова соседних участков, он одновременно создает квантованность на более высоком уровне. В случае неравномерного распределения эдификаторов (ясно выраженной контагиозности), напротив, возникает мелкая квантованность в пределах территории, занятой эдификатором, но благодаря отличию этого гетерогенного участка от соседних одновременно создается квантованность более крупная.
Катастрофические воздействия, разрушающие растительный покров, например пожары, вызывают при восстановлении формирование пятнистого покрова. Этот фактор может по праву рассматриваться среди причин квантованности растительного покрова.
Воздействия животных на растительный покров непосредственно или через среду обитания многообразны. Однако в данном контексте для нас важен один аспект: в зоне интенсивной деятельности животных формируются своеобразные, отличные от соседних, участки растительного покрова. Масштабы таких квантов могут быть самыми разными— от кочки до участков значительных размеров. Яркими примерами здесь могут служить растительность на пороях кабанов, птичьих базаров, песцовые "сады" и др.
Общепризнанным фактором дискретности является деятельность человека, но она обычно остается в стороне, так как, по-видимому, считается очевидной и не относящейся к природным процессам. Но этот факт заслуживает серьезного внимания наряду с другими хотя бы потому, что растительный покров на огромных территориях находится под мощным влиянием хозяйственной деятельности человека, да и геоботаники большей частью исследуют уже измененный человеком растительный покров. С другой стороны, многие механизмы антропогенного воздействия на покров (непосредственно и через изменение среды) принципиально не отличаются от воздействия природных факторов (например, изменение режимов увлажнения и т.п.). Конечно, имеются и специфические воздей-
230
ствия (многие мелиоративные мероприятия, современные концентрированные рубки леса, создание лесных культур и иных форм растительного покрова). По-видимому, было бы неверным утверждать, что влияние человека всегда и везде приводит к квантованности растительного покрова. Так, доброкачественные лесные культуры нередко дифференцированы существенно меньше, чем растительный покров на той же территории до создания культур. Такое же выравнивание, видимо, часто происходит и при мелиорации территорий, приводящей к мезофитизации растительности. Однако и в лесных культурах по мере их роста деревья дифференцируются на группы, в связи с чем формируются ценоэлементы. Таким образом, деятельность человека может находиться в ряду причин квантованности, но не имеет облигатного характера.
Мы не ставили перед собой задачи проанализировать все механизмы формирования растительного покрова. Вместе с тем если признать перечисленные выше причины, то нетрудно прийти к выводу, что они могут приводить к формированию участков растительного покрова с разными закономерностями пространственного сложения. В существовании таких участков, т. е. в расчленении растительного покрова на отдельности, различающиеся закономерностями варьирования основных признаков, и выражается их квантованность.
Эти участки — кванты — могут быть разными: однородными, клинальными. Квант может быть гетерогенным, отличаться высоким уровнем варьирования признаков, имея внутри себя определенный набор пятен. Сами эти пятна также являются квантами, но иного масштаба. Закономерности могут носить и значительно более сложный характер. Например, под отдельно стоящим на лугу деревом изменение травяного покрова имеет радиальную клинальность. Квантованность проявляется в разном масштабе. Кванты могут представлять собой и небольшие группы растений (микрогруипировки Ярошенко или парцеллы Дылиса), выделяться в масштабе, соответствующем привычному нам масштабу фитоценоза и на макроуровне. При этом кванты, как уже указывалось, могут состоять из более мелких квантов. Таким образом, растительный покров можно уподобить мозаичному полотну, на котором более крупные детали слагаются из мелких. Те же причины служат источником квантованности и на низком, синузиальном уровне.
Реальность квантов подтверждается тем, что во всех случаях, когда исследователи ставили соответствующую задачу и использовали объективные и адекватные методы, им удавалось расчленить растительный покров на участки, различающиеся закономерностями сложения. Размер выделенных контуров зависит от размеров учетных единиц и разрешающей способности метода. Необходимо отметить, что при специальном анализе квантованности растительного покрова принимаются во внимание все виды, а не только преобладающие, поэтому кванты могут не совпадать с морфоэлементами, выделяемыми по визуальным оценкам.
Функциональная интерпретация квантов требует дополнительной информации, и может случиться, что два (или больше) морфологических кванта характеризуют одно функциональное образование. Например, трансект, пересекающий фитогенное поле одиночного дерева на лугу, может расчлениться на два клинальных кванта, граница между которыми пройдет около ствола. При детальном членении может выделиться и несколько квантов, соответствующих разным зонам кроны дерева (Прейс, Самойлов, 1977). Такие кванты сами распределены по определенному закону и образуют элементарную форму растительного покрова — коном.
231
Мелкие кванты, выделенные на основе небольших площадок, могут быть организованы по тем же законам в более крупные кванты, в своего рода "пачки" квантов. Выделение таких крупных квантов возможно с применением той же самой методики обработки материала, но с использованием более крупных учетных единиц (площадок). Квантованность растительного покрова на разных масштабных уровнях нашла свое отражение в выделении микро-, мезо- и макрокомбинаций (Исаченко, 1966). Наконец, использование разных признаков — обилие видов, суммарное проективное покрытие синузий, обилие видов с поправкой на их ценотическую значимость, с учетом распространения их корневых систем и т.д. может привести к выявлению разного рода квантов. Все это разнообразие форм квантов свидетельствует о сложной организации растительного покрова; оно же нередко, по-видимому, подталкивает исследователя к простому пути — признанию постулата о континууме растительного покрова, освобождающего его (исследователя) от порой мучительного выделения квантов в натуре.
Сопоставляя причины квантованности с причинами континуума, мы не обнаруживаем взаимозависимости между ними в том смысле, что чем сильнее выражены одни, тем слабее другие, и в крайнем своем выражении они не исключают друг друга. Континуум и квантованностъ — коренные свойства растительного покрова и выражены одновременно и всюду. Следует особо подчеркнуть, что континуум имеет место не только на границах квантов, но и внутри их. Это обстоятельство приводит к тому, что границу в виде гиперплоскости или какой-либо определенной площадки (при исследовании трансект) между квантами установить в принципе невозможно, положение ее всегда неопределенно. В соответствии с тем, что континуум и квантованность — разные свойства растительного покрова, методы их исследования должны быть разными.
ГЛАВА 7. ДИНАМИКА РАСТИТЕЛЬНЫХ СООБЩЕСТВ
Растительные сообщества находятся в постоянном изменении. На уровне особи динамика проявляется в изменении размеров растений и фенологическом развитии, на уровне популяции — в изменении возрастного спектра и обилия (численности, проективного покрытия, фитомассы). На уровне сообщества изменяется видовой состав, соотношение обилия видовых популяций, экологических групп, жизненных форм и строения.
Причины изменений разнообразны. Они заключены в природе самих растений — в генетической программе их развития, выражающейся в онтогенезе. "Растекание жизни" приводит к внедрению в сообщества новых видов. Трансформация растениями среды и, как следствие, взаимодействие растений являются сильнейшими факторами изменения сообществ. Изменение экотопа меняет условия реализации трех первых причин. Деятельность гетеротрофных организмов, воздействующих на растения как непосредственно (например, поедание растений фитофагами), так и через среду, вызывает изменения в растительных сообществах. Особую роль играет человек. Изымая фитомассу (при сенокошении, заготовке древесины), человек выступает как своеобразный фитофаг. Своей хозяйственной деятельностью он влияет и на все остальные причины динамики растительности. Производя химические вещества и сознательно используя их либо распространяя как отходы промышленности, он влияет на развитие растений. Миграция химических веществ, изменяя условия существования гетерофагов, вызывает изменение их численности и состава, а также активности. Человек— участник расселения растений. Изменяя состав и строение сообществ, человек вмешивается.в трансформацию растениями среды. Последствия выпаса домашних животных во многом аналогичны воздействию диких животных.
Все процессы, происходящие при изменениях растительности, в зависимости от причин, их порождающих, можно разделить на эндогенез (эндоэкогенез) и экзогенез (экзоэкогенез). При эндогенезе движущей силой динамики являются сами растения, а экзогенез вызывают внешние по отношению к сообществу факторы— экотоп. биоцен. человек. Динамику состава, соотношения обилия видов, строения растительности определяют одновременно и эндогенез, и экзогенез, но вклад их в зависимости от конкретных обстоятельств может быть разным— то один, то другой играет большую роль.
Изменения растительных сообществ могут иметь циклический характер. К ним относятся изменения, происходящие в течение вегетационного периода или при смене времен года в тех зонах, где вегетационный период длится круглый год. Это сезонная изменчивость. Колебания погодных условий от года к году приводят к колебанию параметров растительных сообществ около средних значений за определенный период времени. Их называют флуктуациями (флуктуационной изменчивостью), такие изменения относительно обратимы, цикличны. Направленные необратимые изменения и называются сукцессиями.
Сезонная изменчивость
Общей закономерностью сезонной изменчивости является зависимость параметров растительного сообщества от изменения экологических факторов в течение вегетационного периода. Ритм развития растений и сообщества в целом определяется в основном температурным режимом и
233
234

снабжением растений влагой. Образное описание сезонной изменчивости разнотравного пойменного ненадолго заливаемого луга около г.Вологды приводит в своем учебнике "Луговедение" А. П.Шенников (1941). "В ходе его ежегодного развития в течение периода вегетации выделяются 7 стадий. Первая стадия — предвесенняя — продолжается от времени оттаивания почвы примерно до половины мая. В это время происходит позеленение луга от появляющихся новых побегов и развертывание свежих прикорневых листьев, появляются колоски лугового хвоща и бутоны обильной здесь фиалки (Viola arenaria). У многих растений, зимовавших без зеленых листьев, появляются надземные побеги с розетками листьев. В зависимости от температуры эта стадия наступает несколько раньше или позже, в связи с временем первой весенней волны тепла, и проходит быстрее или медленнее, в связи с обычной на севере волной холода.
С новым повышением температуры луг переходит во вторую стадию — ранневесеннюю. В ранневесеннюю стадию луг имеет фиалковый аспект от массы цветущей Viola arenaria. Одновременно цветет Glechorna hederacea и пылит хвощ. Формируется подсед — кустятся злаки, начинается стебление разнотравья. Замечается прирост мха (Thuidium abietinum). Эта стадия продолжается до конца мал, затягиваемая очередной волной холода. Третья стадия — разгар (середина) фенологической весны — начинается с новым, более устойчивым повышением температуры. Аспект характеризуется обилием цветущего Trollius curopaeus; луг от него желтеет. Тогда же цветут, зацветая одновременно или почти одновременно, Geum nvale. Taraxacum vulgare, Polygala amara, Alchimilla pubescens, Carum cam.
Ranunculus acer и др.; луг делается очень цветистым. Это время усиленного роста травостоя, стебления низовых злаков, формирования 2-го яруса, постепенного затенения мха и приземистых растений. Продолжается до
10-15 июня.
Четвертая стадия — конец весны — начало лета, т.е. переход от весны к лету. Массовое цветение Polygonum bistorta, луг от него розовый. За-
цветают Trifolium pratense, Rumex haplorhizus и другие летние растения, и
цветущих растений становится больше, чем в предыдущую стадию. Ранние злаки в это время заканчивают колошение (красная овсяница, луговой
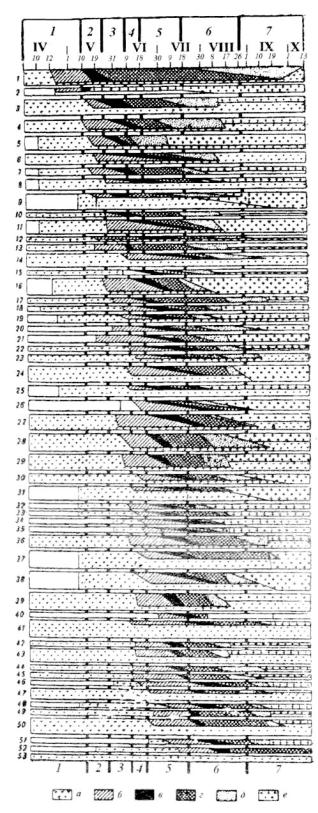
Рис. 7.1. Фенологический спектр мелкозлаково-разнотравного краткопоемного луга около Вологды (по: Шснников, 1941).
По вертикали: 1 — Viola arenaria, 2 — Equisetum pratense, 3 — Glechorna hederacea, 4— Geum rivale, 5— Taraxacum vulgare, 6— Polygala arnarella, 7- — Alchtrnilla pubescens, 8— Carum carvi, 9— Trollius europaeus, 10— Fragaria vesca, 11— Ranunculus acer, 12 — Plantago media, 13— Luzula campestris, 14— Trifolium pratense, 15 — Cerastium trivialc, 16— Polygonum bistorta, 17— Veronica chamaedrys. 18— Anthriscus silvestris, 19— Rumex haplorhizus, 20— Ranunculus polyanthemus, 21 — Polygala comosa, 22— Stellaria graminen, 23— Trifolium repens, 24— Vicia cracca, 25— Tragopogon pratensis, 26— Euphrasia hirtella, 27— Leucanthemum vulgare, 28— Poa pratensis, 29— Festuca rubra, 30— Lathyrus pratensis, 31 — Geranium pratense, 32— Silene inflata, 33— Heracleum sibiricum, 34— Alectorolophus major, 35 — Leontodon autumnalis, 36— Galium mollugo, 31— Galium boreale, 38— Campanula glomerata, 39-Deschampsia caespitosa, 40— Brunella vulgaris, 41 — Achillea millefolium, 42— Briza media, 43— Festuca pratensis, 44 — Dactylis glomerata, 4 5 — Bromus inermis, 46 — Pimpine.lla saxijraga, 47— Agrostis alba, 4$— Delphinium elatum, 49— Phleum pratense, 50 Agropyrum repens, 51 — Hieracium umbellatum, 52— Tanacetum vulgare, 53— Antennaria dioica.
Ширина полосок видовых спектров соответствует проценту встречаемости вида, ширина полосок генеративной фазы — проценту встречаемости генеративных побегов.
IV-X — апрель-октябрь. По горизонтали: 1-7— сезонные стадии луга: предвесенняя ( / ) , ранневесенняя (2), середины весны (3), конца весны-начала лета (4), разгара лета (5), конца лета (6), осенняя (7). а-е — фенологические фазы: a — вегетативная, 6— бутоны, в — цветение, г — созревание семян, д — обсеменение, е— отмирание.
235
мятлик). Формируется 1-й (верхний) полог верховых злаков и высокого разнотравья. Эта стадия обычно коротка.
Пятая стадия — разгар лета (или середина фенологического лета); начинается около 20 июня и продолжается до 15-20 июля. Начало ее зацветание низовых злаков (красной овсяницы и лугового мятлика) и вместе с ними многих других растений, в том числе массы Leucanthemum vulgare и Campanula glomerata, характеризующих пестрый аспект луга. Это время — апогей фенологического развития нашего луга. Травостой сформирован полностью. Число цветущих видов наибольшее. Зацветают и поздние злаки.
В шестой стадии — конец лета — цветистость луга уменьшается, и в аспекте его более заметны окраски обильных плодоношений. Заметны также увядание и отмирание нижних листьев и побегов. Число цветущих видов уменьшается, хотя некоторые (иозднелетние) зацветают только теперь
(Hieracium umbellatum, Agropyrum re-pens, Delphinium elatum).
Формируются почки перезимовывания. Взамен отмирающих генеративных побегов появляются новые вегетативные. Характерно, что увядание прогрессирует несмотря на то, что погода еще теплая; лишь в конце этой стадии— около половины августа — начинаются заметные похолодания. И хотя после них снова бывает еще довольно долгое время тепло, луг переходит в осеннее состояние.
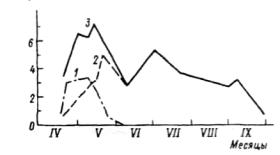
Седьмая стадия — осеннее состояние луга — время явного преобладания во внешности луга признаков увядания и отмирания травостоя. Цветущие растения единичны или отсутствуют. Продолжается новое побегообразование, особенно кущение злаков, дающих новое поколение вегетативных побегов под отмирающим весенне-летним поколением плодущих побегов. Эта стадия продолжается до морозов и снега (с.240-241). Графическое изображение прохождения фенологических фаз видами, слагающими сообщество, называется фенологическим спектром (рис.7.1). Обобщенной характеристикой фенологического состояния сообщества является число видов, находящихся в одной фазе развития, например цветения. Число цветущих видов постепенно увеличивается, достигает максимума в середине лета (с середины июня до середины июля) и постепенно уменьшается. Ритм развития видов индивидуален. Он отражает неодинаковое отношение к экологическим факторам.
и,/га
Рис. 7.2. Динамика надземной массы в травяном покрове дубравы (липо-дубняк снытевый, лесостепь) (по: Горышина, 1969).
/ — эфемероиды, 2— виды летней синузии, 3 — суммарная фитомасса.
Существенную роль в сезонной динамике играет изменение световой обстановки в толще растительного покрова. В лесостепной дубраве под
236

Рис. 7.3. Кривая цветения травянистых видов под пологом дубравы (липо-дубняк. лесостепь) (по: Горышина, 1969).
А, 1 — число цветущих видов; Б, 2— относительная освещенность (%) на уровне травяного покрова.
пологом древостоя большинство видов ускоренно накапливают фитомассу до момента полного развития листьев деревьев (рис.7.2), т.е. в условиях достаточной освещенности. Пик цветения наблюдается также до смыкания полога (рис.7.3). Значительную долю в образовании весеннераннелетнего пика составляют эфемероиды (Scilla sibinca. Corydalis halleri,
Anemone ranunculoides, Gagea lutea. G.minima. Ficana verna), требовательные к свету и способные быстро развиваться при сравнительно невысоких температурах благодаря запасенным в подземных органах питательным веществам. Пик весеннего цветения формируется и видами, цветущими несколько позднее эфемероидов, — p. Viola, Latkyrus pern us. Carex pilosa,
Asarum europaeum, Glechoma hederacea и др. (Горышина, 1969).
Рис. 7.4. Распределение листовой поверхности по высоте у лабазниковых (Filipendula camtschatica) травостоев в расчете на участок площадью
1м2, Камчатка (по: Степанова и др., 1981).
а—15 июня, 6—23 июля, в — 6 июля, г — 15 июля, д—28 июля, е —
9августа.
Втечение вегетационного периода часто изменяется вертикальное распределение листовой поверхности и фитомассы. Особенно ярко это выражено в сомкнутых крупнотравных сообществах, как, например, с господством лабазника — Ftlipendula camtschatica (рис.7.4, 7.5). Весной листо-
237

238