

221 :: 222 :: 223 :: Содержание
7.3. Факторы, определяющие чувствительность рецепторов
С какой точностью сенсорная информация передается в ЦНС? Можно ли сравнивать в этом отношении органы чувств с такими физическими приборами, как термометр, люксметр или тензометр? Из собственного опыта все мы знаем, что биологические сенсорные системы не очень надежны как индикаторы абсолютного уровня энергии. Кроме того, со временем многие ощущения изменяются. Так, например, человеку, вышедшему из темного помещения, приятный солнечный день в первые минуты может показаться болезненно ярким. По этой причине даже опытному фотографу для точного определения выдержки требуется люксметр. Человеку, входящему в необогреваемый водоем поплавать, вода сначала кажется холоднее, чем минуту-другую спустя. Такого рода феномены объединяют общим термином сенсорная адаптация. Где в организме происходит эта адаптация? Ответить на этот вопрос не просто. Одни адаптационные процессы протекают в рецепторных клетках, другие развиваются в результате изменений во вспомогательных тканях, третьи-в ЦНС.
7.3.1. Рецепторная адаптация
У рецепторов разного типа адаптация выражена в разной степени. Тонические (медленно адаптирующиеся) рецепторы при непрерывном воздействии стимула разряжаются с постоянной частотой. Так ведет себя, например, детектор смещения (рис. 7-9, А), разряжающийся с почти постоянной частотой в ответ на длительное изменение его положения. Многие рецепторы относятся к так называемому фазическому (быстро адаптирующемуся) типу-их импульсация замедляется или даже прекращается вскоре после начала воздействия постоянного стимула. Фазические рецепторы одного из подтипов разряжаются т о л ь ко во время изменения интенсивности стимула. Так, некоторые механорецепторы посылают импульсы только во время смещения, причем частота разряда зависит от скорости смещения (рис. 7-9, Б).
Адаптация рецептора может происходить на разных этапах цепи событий от воздействия стимула до генерации нервных импульсов (рис. 7-10).
1. Сенсорный фильтр может избирательно пропускать кратковременные стимулы; это характерно для многих механорецепторов.
2. При воздействии постоянного стимула может уменьшаться число молекул, преобразующих его энергию. Например, при длительном воздействии света обесцвечивается значительная часть молекул зрительного пигмента, и для того, чтобы он
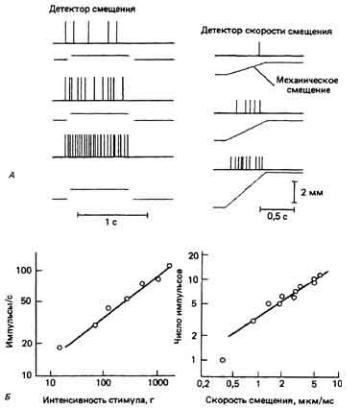
Рис. 7.9. Сравнение детекторов смещения и
скорости смещения. А. Поведение детектора смещения. Данный тонический (т.е. медленно адаптирующийся) механорецептор реагирует на длительное смещение (показанное цветными линиями) импулъсацией сравнительно постоянной частоты. Вверху: ответ детектора на три стимула разной интенсивности. Внизу: зависимость частоты разряда в фазе стабильной импульсации от растягивающей силы. Б. Поведение детектора скорости смещения. Этот быстро адаптирующийся механорецептор реагирует на скорость изменения положения тканей. Вверху: чем выше скорость смещения (цветные линии), тем больше возникает импульсов. Внизу: число импульсов в ответе на 0,5-секундное растяжение пропорционально логарифму скорости смещения. (Schmidt, 1971.)
221

Рис. 7.70. Различные уровни, на которых возможна сенсорная адаптация. Пунктирные линии означают, что в одних случаях процессы сенсорной фильтрации и (или) модуляции импульсного разряда происходят в самой рецепторной клетке, а в других-за ее пределами.
вновь приобрел способность реагировать на свет, необходима его метаболическая регенерация.
3. В некоторых рецепторах в результате накопления регуляторного вещества может ингибироваться каскад ферментативных реакций, активируемый рецепторной молекулой. Полученные недавно данные указывают на то, что при адаптации к свету зрительных рецепторов позвоночных такую функцию регулятора, возможно, рыполняет Са2+.
4. Во время длительной стимуляции
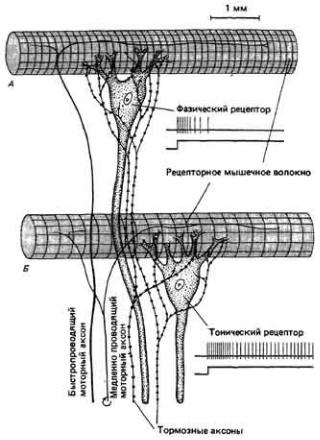
Рис. 7.11. Фазический (А) и тонический (Б) рецепторы растяжения речного рака. Фазический рецептор при длительном растяжении быстро адаптируется и генерирует лишь короткую серию импульсов.
Тонический же рецептор в этом случае дает продолжительный разряд.
рецепторной клетки могут изменяться ее электрические свойства. В некоторых рецепторах длительная стимуляция ведет к накоплению Са2+ внутри клетки, что снижает активность рецепторных каналов. Накопление Са2+ может также приводить к активации Са2+-зависимых калиевых каналов и в результате-к сдвигу мембранного потенциала в сторону потенциала покоя.
5. Возможна адаптация мембраны в зоне инициации импульсов (см. разд. 5.6.1 и рис. 7-4).
6. Сенсорная адаптация может происходить и в центральной нервной системе (к которой относится и сетчатка позвоночных животных).
Первый и пятый из перечисленных механизмов адаптации свойственны, в частности, мышечным рецепторам растяжения речного рака и омара. Эти рецепторы расположены в мускулатуре брюшка попарно, причем каждая пара состоит из фазического и тонического рецепторов. Скачкообразное увеличение длины рецепторного мышечного волокна вызывает кратковременную реакцию первого из них и
222
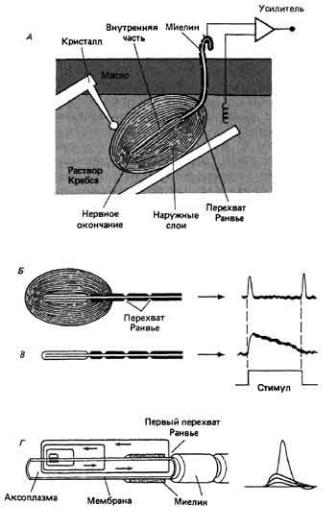
Рис. 7.12. Адаптация тельца Пачини, A. Экспериментальная установка для раздражения рецептора иглой с пъезокристаллическим приводом. Электрическая активность рецептора регистрируется между крючковидным электродом и поверхностью раздела масло/вода. Б, Электрический ответ интактного тельца Пачини. Как при включении, так и при выключении стимула возникают кратковременные деполяризационные сдвиги. B. Если освободить рецептор от наружного пластинчатого слоя, его ответ на длительное смещение становится длительным. Г. Рецепторный ток, вызываемый деформацией чувствительной зоны аксона. Импульс возникает из рецепторного потенциала (справа) в первом перехвате Ранвье. (Loewenstein, 1960.)
длительный разряд второго (рис, 7-11, Б). Если рецепторный ток шунтировать прямым пропусканием через микроэлектрод деполяризующего тока, обе рецепторные клетки сохраняют некоторые из своих электрических свойств. Например, фазический рецептор будет реагировать на длительное пропускание тока более быстрым снижением частоты импульсации, чем тонический рецептор при тех же условиях стимуляции.
"Фильтрующее" действие вспомогательных структур (первый из перечисленных выше механизмов адаптации) играет важную роль в быстрой адаптации телец Пачини-рецепторов давления и вибрации, имеющихся в коже, мышцах, брыжейке, сухожилиях и суставах млекопитающих (рис. 7-12). Они состоят из нервного окончания" чувствительного к механическому раздражению, и окружающей его капсулы-концентрических слоев соединительной ткани, подобных слоям чешуек в луковице. Когда тельце при давлении на него деформируется, эта деформация через капсулу механически передается чувствительной мембране нервного окончания. Последнее обычно реагирует кратковременной деполяризацией как в начале деформации, так и при

ее снятии (рис. 7-12, Б). Если нервное окончание тельца Пачини освободить от капсулы, позволив тем самым механическому стимулу воздействовать прямо на голый аксон, то возникнет рецепторный потенциал, который будет гораздо более точным аналогом внешнего стимула, чем рецепторный потенциал в интактном тельце Пачини. Хотя рецепторный потенциал и в этом случае обнаруживает некоторую адаптацию (спад на рис. 7-12, В), он появляется только в начале деформации. Быстрая адаптация интактного тельца Пачини к длительно поддерживаемой деформации отчасти, вероятно, обусловлена фильтрующими свойствами капсулы, избирательно передающей быстрые изменения давления. Этим можно частично объяснить тот факт, что мы быстро перестаем ощущать постоянное давление умеренной силы.
Адаптация независимо от места и механизмов ее развития играет важную роль в расширении динамического диапазона сенсорной рецепции. Наряду с логарифмическим характером первичного преобразования стимула сенсорная адаптация позволяет животному замечать малые изменения в силе стимула при самых различных уровнях его абсолютной интенсивности (которая может различаться на много порядков величины).
223
221 :: 222 :: 223 :: Содержание

Физиология животных: механизмы и адаптации. Учебник в двух томах. Том 1. Эккерт Р., Рэндалл Д.,
Огастин Д. М.: Мир, 1991. 424c. ISBN 5-03-001457-8
Содержание
От редактора перевода |
5 |
Предисловие |
6 |
Благодарности |
7 |
Глава 1. Предмет физиологии |
|
1.1. Для чего нужна физиология животных |
9 |
1.2. Физиология и медицина |
10 |
1.3. Физиология и познание |
10 |
1.4. Основные концепции физиологии |
|
1.4.1. В основе любой функции лежит структура |
11 |
1.4.2. Генетика и физиология |
11 |
1.4.3. Принцип гомеостаза |
12 |
1.5. Физиологическая литература |
14 |
1.6. Резюме |
16 |
1.7. Вопросы для повторения |
16 |
Литература |
16 |
Глава 2. Физические и химические концепции |
17 |
2.1. Атомы, связи и молекулы |
19 |
2.2. Свойства Н, О, N и C как основа для возникновения жизни |
21 |
2.3. Вода |
|
2.3.1. Молекула воды |
22 |
2.3.2. Свойства воды |
24 |
2.3.3. Вода как растворитель |
24 |
2.4. Растворы и их коллигативные свойства |
26 |
2.5. Растворы электролитов |
|
2.5.1. Ионизация воды |
28 |
2.5.2. Кислоты и основания |
29 |
2.5.3. Биологическая роль рН |
30 |
2.5.4. Уравнение Гендерсона-Хассельбалха |
30 |
2.5.5. Буферные системы |
31 |
2.6. Электрический ток в водных растворах |
32 |
2.7. Ионная избирательность |
34 |
2.8. Биологические молекулы |
|
2.8.1. Липиды |
36 |
2.8.2. Углеводы |
37 |
2.8.3. Белки |
38 |
2.8.4. Нуклеиновые кислоты |
44 |
2.9. Резюме |
46 |
2.10. Вопросы для повторения |
48 |
Литература |
48 |
Глава 3. Ферменты и энергетика |
49 |
3.1. Энергия: понятия и определения |
50 |
3.2. Перенос химической энергии в системе сопряженных реакций |
53 |
3.3. АТР и высокоэнергетическая фосфатная группа |
55 |
3.4. Температура и скорость реакции |
57 |
3.5. Ферменты |
58 |
3.5.1. Специфичность фермента |
59 |
3.5.2. Каталитическая активность |
60 |
3.5.3. Температура и скорость реакции |
61 |
3.5.4. Чувствительность к рН |
61 |
3.5.5. Регуляция ферментативной активности |
61 |
3.5.6. Кофакторы |
62 |
3.5.7. Кинетика ферментативных реакций |
63 |
3.5.8. Сродство между ферментом и субстратом |
64 |
3.5.9. Подавление активности ферментов |
65 |
3.6. Механизмы регуляции метаболизма |
|
3.6.1. Генетическая регуляция синтеза ферментов |
67 |
3.6.2. Метаболическое ингибирование по типу обратной связи |
68 |
3.6.3. Активация ферментов |
69 |
3.7. Образование АТР в процессе метаболизма |
69 |
3.8. Окисление, фосфорилирование и перенос энергии |
72 |
3.8.1. Электронпереносящие коферменты |
73 |
3.9. Цепь переноса электронов |
75 |
3.10. Гликолиз |
78 |
3.11. Цикл трикарбоновых кислот (цикл Кребса) |
80 |
3.12. Эффективность энергетического метаболизма |
82 |
3.13. Кислородная задолженность |
83 |
3.14. Резюме |
83 |
3.15. Вопросы для повторения |
84 |
Литература |
85 |
Глава 4. Проницаемость и транспорт |
86 |
4.1. Состав мембран |
87 |
4.2. Организация мембран |
|
4.2.1. Простые модели бислоев |
89 |
4.2.2. Жидкостно-мозаичная модель |
90 |
4.2.3. Субъединичная модель |
92 |
4.3. Физические основы проницаемости мембран |
|
4.3.1. Диффузия |
94 |
4.3.2. Трансмембранный поток |
94 |
4.3.3. Осмос |
95 |
4.3.4. Осмоляркость и тоничность |
96 |
4.3.5. Влияние электрических сил на распределение ионов |
97 |
4.3.6. Доннановское равновесие |
97 |
4.4. Осмотические свойства клеток |
|
4.4.1. Стационарное состояние |
99 |
4.4.2. Объем клеток |
100 |
4.5. Механизмы пассивного транспорта |
101 |
4.5.1. Простая диффузия через липидный бислой |
102 |
4.5.2. Диффузия через мембранные каналы |
103 |
4.5.3. Облегченная диффузия |
104 |
4.6. Активный транспорт |
105 |
4.7. Ионные градиенты как источники энергии в клетке |
107 |
4.7.1. Симпорт |
109 |
4.7.2. Антипорт |
110 |
4.8. Селективность мембран |
110 |
4.8.1. Селективность к электролитам |
111 |
4.8.2. Селективность к неэлектролитам |
111 |
4.9. Эндоцитоз и экзоцитоз |
112 |
4.10. Межклеточные контакты |
|
4.10.1. Щелевые контакты |
114 |
4.10.2. Плотные контакты |
116 |
4.11. Эпителиальный транспорт |
116 |
4.11.1. Активный транспорт солей через эпителий |
117 |
4.11.2. Транспорт воды |
120 |
4.12. Резюме |
123 |
4.13. Вопросы для повторения |
124 |
Литература |
125 |
Глава 5. Ионы и возбуждение |
126 |
5.1. Мембранная теория возбуждения |
127 |
5.2. Пассивные электрические свойства клеточных мембран |
|
5.2.1. Проводимость мембраны |
132 |
5.2.2. Емкость мембраны |
133 |
5.2.3. Электротонический потенциал |
134 |
|
5.3. Электрохимический потенциал |
135 |
|
5.3.1. Уравнение Нернста |
137 |
|
5.4. Потенциал покоя |
|
|
5.4.1. Роль ионных градиентов и ионных каналов |
138 |
|
5.4.2. Роль активного транспорта |
139 |
|
5.5. Активные электрические процессы |
141 |
|
5.6. Ионные основы потенциала действия |
|
|
5.6.1. Общие свойства потенциала действия |
142 |
|
5.6.2. Натриевая гипотеза |
144 |
|
5.6.3. Натриевые каналы |
148 |
|
5.6.4. Цикл Ходжкина |
150 |
|
5.6.5. Калиевый ток |
152 |
|
5.6.6. Ионные механизмы потенциала действия: краткая сводка |
154 |
|
5.6.7. Изменение концентрации ионов во время возбуждения |
155 |
|
5.7. Другие электровозбудимые каналы |
155 |
|
5.8. Пейсмекерные потенциалы |
158 |
|
5.9. Резюме |
162 |
|
5.10. Вопросы для повторения |
162 |
|
Литература |
163 |
|
Глава 6. Распространение и передача нервных импульсов |
|
|
6.1. Нервные клетки |
164 |
|
6.1.1. Два основных типа электрических сигналов в нервных |
167 |
|
клетках |
||
|
||
6.2. Пассивное распространение электрических сигналов |
168 |
|
6.3. Распространение нервных импульсов |
169 |
|
6.3.1. Скорость распространения нервных импульсов |
171 |
|
6.3.2. Сальтаторное проведение |
173 |
|
6.4. Представление о синапсах |
173 |
|
6.5. Передача возбуждения в электрических синапсах |
174 |
|
6.6. Передача сигналов в химических синапсах |
175 |
|
6.6.1. Строение химических синапсов |
177 |
|
6.6.2. Синаптические потенциалы |
179 |
|
6.6.3. Синаптические токи |
180 |
|
6.6.4. Потенциал реверсии |
181 |
|
6.6.5. Постсинаптическое торможение |
184 |
|
6.6.6. Пресинаптическое торможение |
186 |
|
6.7. Постсинаптические рецепторы и каналы |
186 |
|
6.7.1. АцХ-активируемый канал |
187 |
|
6.8. Выделение медиаторов пресинаптическими окончаниями |
189 |
6.8.1. Квантовое выделение медиаторов |
190 |
|
6.8.2. Электросекреторное сопряжение |
192 |
|
6.9. Синаптическая интеграция |
193 |
|
6.9.1. Суммация |
195 |
|
6.10. Функциональная пластичность синапсов |
199 |
|
6.10.1. Гомосинаптическая модуляция |
200 |
|
6.10.2. Гетеросинаптическая модуляция |
202 |
|
6.11. Медиаторы |
204 |
|
6.11.1. Биогенные амины |
205 |
|
6.11.2. Аминокислоты |
205 |
|
6.11.3. Нейропептиды |
206 |
|
6.11.4. Эндогенные опиоиды |
207 |
|
6.12. Резюме |
211 |
|
6.13. Вопросы для повторения |
212 |
|
Литература |
213 |
|
Глава 7. Сенсорные механизмы |
214 |
|
7.1. Рецепторные клетки как сенсорные преобразователи |
215 |
|
7.1.1. События, происходящие между сенсорным |
216 |
|
преобразованием и сенсорным выходом |
||
|
||
7.2. Кодирование интенсивности стимула |
|
|
7.2.1. Зависимость между входом и выходом |
219 |
|
7.2.2. Дробление динамического диапазона |
220 |
|
7.3. Факторы, определяющие чувствительность рецепторов |
|
|
7.3.1. Рецепторная адаптация |
221 |
|
7.3.2. Механизмы повышения чувствительности |
223 |
|
7.3.3. Эфферентный контроль чувствительности рецепторов |
224 |
|
7.3.4. Торможение рецепторов по принципу обратной связи |
225 |
|
7.4. Хеморецепция |
225 |
|
7.5. Механорецепция |
228 |
|
7.5.1. Волосковые клетки |
229 |
|
7.5.2. Органы равновесия |
231 |
|
7.6. Ухо млекопитающих |
|
|
7.6.1. Структура и функции улитки |
233 |
|
7.6.2. Возбуждение волосковых клеток улитки |
235 |
|
7.6.3. Частотный анализ звуков в улитке |
236 |
|
7.7. Электрорецепторы |
236 |
|
7.8. Терморецепторы |
238 |
|
7.9. Фоторецепторы |
|
|
7.9.1. Фоторецепторы беспозвоночных |
239 |
|
7.9.2. Зрительные рецепторные клетки позвоночных |
241 |
|
7.9.3. Зрительные пигменты |
245 |
7.9.4. Фотохимия зрительных пигментов |
246 |
|
7.9.5. Цветовое зрение |
248 |
|
7.10. Оптические приспособления |
|
|
7.10.1. Сложные глаза членистоногих |
250 |
|
7.10.2. Глаз позвоночных |
253 |
|
7.11. Резюме |
256 |
|
7.12. Вопросы для повторения |
257 |
|
Литература |
258 |
|
Глава 8. Переработка информации в нервной системе и поведение |
259 |
|
8.1. Эволюция нервной системы |
260 |
|
8.2. Нервная система позвоночных |
263 |
|
8.2.1. Главные отделы центральной нервной системы |
265 |
|
8.2.2. Вегетативная нервная система |
267 |
|
8.3. Нейронные цепи |
270 |
|
8.4. Сети сенсорной фильтрации |
|
|
8.4.1. Латеральное торможение |
274 |
|
8.4.2. Переработка зрительной информации в сетчатке |
277 |
|
позвоночных |
||
|
||
8.4.3. Переработка информации в зрительной коре |
281 |
|
8.5. Двигательные нейронные сети |
284 |
|
8.5.1. Миотатический рефлекс (рефлекс на растяжение) |
285 |
|
8.5.2. Эфферентный контроль через гамма-петлю |
287 |
|
8.5.3. Сухожильный рефлекс Гольджи |
287 |
|
8.5.4. Сгибательный рефлекс и реципрокная иннервация |
290 |
|
8.6. Запрограммированное поведение |
|
|
8.6.1. Центрально генерируемые двигательные ритмы |
290 |
|
8.6.2. Комплексы фиксированных действий |
293 |
|
8.7. Поведение животных, не имеющих нервной системы |
294 |
|
8.8. Инстинктивное поведение |
297 |
|
8.9. Модификация поведения |
299 |
|
8.10. Ориентация животных |
|
|
8.10.1. Таксисы и корректирующие реакции |
301 |
|
8.10.2. Вибрационная ориентация |
303 |
|
8.10.3. Эхолокация |
304 |
|
8.11. Навигация животных |
|
|
8.11.1. Использование биологических часов |
306 |
|
8.11.2. Геомагнитные ориентиры |
306 |
|
8.12. Резюме |
308 |
|
8.13. Вопросы для повторения |
309 |
|
Литература |
310 |
Глава 9. Химические посредники и регуляторы |
311 |
|
9.1. Концепция первого и второго посредников |
314 |
|
9.2. Циклические нуклеотиды как вторые посредники |
|
|
9.2.1. Циклический аденозинмонофосфат |
315 |
|
9.2.2. Множественные эффекты сАМР |
320 |
|
9.2.3. Мобилизация глюкозы: биохимическая модель действия |
321 |
|
сАМР как второго посредника |
||
|
||
9.2.4. Циклический гуанозинмонофосфат |
321 |
|
9.3. Мембранные липиды в роли посредника |
322 |
|
9.4. Кальций как внутриклеточный посредник |
323 |
|
9.4.1. Кальмодулин и другие белки, связывающие кальций |
328 |
|
9.5. Внутриклеточное действие стероидных гормонов |
328 |
|
9.6. Классификация гормонов |
|
|
9.6.1. Идентификация эндокринных желез и гормонов |
332 |
|
9.7. Регуляция секреции гормонов |
|
|
9.7.1. Обратная связь |
333 |
|
9.7.2. "Упаковка" и хранение гормонов |
334 |
|
9.7.3. Механизм секреции |
335 |
|
9.8. Нервно-эндокринные связи |
337 |
|
9.9. Нейросекреция |
338 |
|
9.10. Эндокринные системы позвоночных |
339 |
|
9.11. Гипоталамус и гипофиз |
|
|
9.11.1. Гормоны нейрогипофиза |
339 |
|
9.11.2. Гормоны аденогипофиза |
340 |
|
9.11.3. Гипоталамический контроль функций аденогипофиза |
342 |
|
9.12. Гормоны, регулирующие метаболизм и развитие |
|
|
9.12.1. Глюкокортикоиды |
344 |
|
9.12.2. Гормоны щитовидной железы (тиреоидные гормоны) |
345 |
|
9.12.3. Катехоламины |
348 |
|
9.12.4. Инсулин и глюкагон |
351 |
|
9.12.5. Гормон роста (соматотропный гормон, СТГ) |
353 |
|
9.13. Гормональная регуляция водно-солевого баланса |
354 |
|
9.14. Простагландины |
356 |
|
9.15. Половые гормоны |
357 |
|
9.15.1. Репродуктивные циклы |
360 |
|
9.16. Эндокринные системы насекомых |
363 |
|
9.16.1. Регуляция развития насекомых |
365 |
|
9.17. Резюме |
373 |
|
9.18. Вопросы для повторения |
374 |
|
Литература |
375 |
|
Глава 10. Мышцы и движение |
|
10.1. Структурные основы сокращения |
376 |
|
10.1.1. Тонкая структура миофиламентов |
379 |
|
10.2. Теория скольжения нитей |
381 |
|
10.2.1. Кривая зависимости "длина -сила" |
383 |
|
10.3. Функция поперечных мостиков и развитие силы |
|
|
10.3.1. Химия активности поперечных мостиков |
383 |
|
10.3.2. Активность поперечных мостиков и мышечное |
384 |
|
сокращение |
||
|
||
10.4. Роль кальция в процессе сокращения |
|
|
10.4.1. Активация поперечных мостиков |
387 |
|
10.4.2. Инактивация поперечных мостиков и расслабление |
388 |
|
мышцы |
||
|
||
10.5. Электромеханическое сопряжение |
389 |
|
10.5.1. Мембранный потенциал и сокращение |
390 |
|
10.5.2. Саркотубулярная система |
391 |
|
10.5.3. Саркоплазматический ретикулум |
393 |
|
10.5.4. Высвобождение кальция саркоплазматическим |
393 |
|
ретикулумом |
||
|
||
10.5.5. Краткое описание процессов сокращения и расслабления |
396 |
|
10.6. Механические свойства сокращающейся мышцы |
397 |
|
10.6.1. Длина саркомера и сократительные свойства |
398 |
|
10.6.2. Латентный период |
398 |
|
10.6.3. Зависимость "сила - скорость" |
398 |
|
10.6.4. Последовательные эластические компоненты |
399 |
|
10.6.5. Активное состояние |
400 |
|
10.6.6. Одиночное и тетаиическос сокращение |
401 |
|
10.6.7. Энергетика сокращения |
402 |
|
10.7. Метаболические подтипы поперечнополосатых мышц |
403 |
|
10.8. Нервная регуляция мышечною сокращения |
|
|
10.8.1. Нейромоторная организация позвоночных |
405 |
|
10.8.2. Нервно-мышечная организация членистоногих |
407 |
|
10.8.3. Асинхронные летательные мышцы насекомых |
408 |
|
10.9. Сердечная мышца |
411 |
|
10.10. Гладкая мышца |
412 |
|
10.11. Скелетно-мышечная механика |
414 |
|
10.12. Резюме |
419 |
|
10.13. Вопросы для повторения |
420 |
|
Литература |
421 |