

6 курс / Эндокринология / Механизмы_нейроэндокринной_регуляции_Угрюмов_М_В_1999
.pdf


ВВЕДЕНИЕ
НЕЙРОЭНДОКРИННАЯ РЕГУЛЯЦИЯ
УВЫСШИХ ПОЗВОНОЧНЫХ
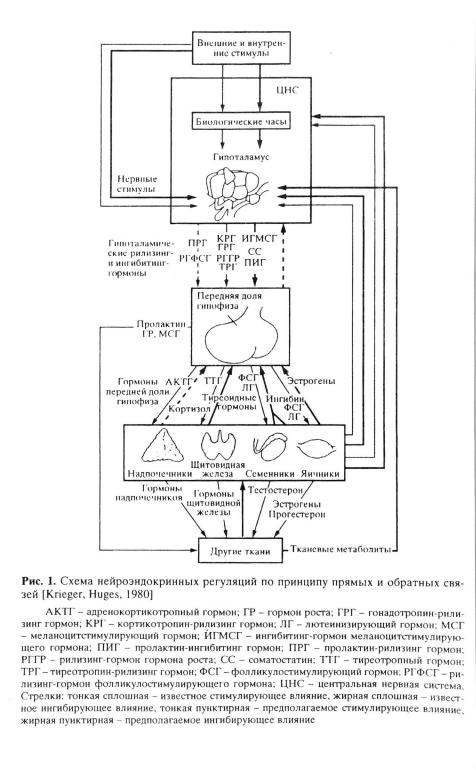
впоследние три десятилетия получены убедительные доказательства того, что нейроэндокринная регуляция играет ключевую роль в интеграции организма и в поддержании гомеостаза, обеспечивая при этом контроль важнейших функций, таких как репродукция, адаптация и др. [Jackson, Mueller, 1982; Vincent, Simonnet, 1986; Baulieu, Kelly, 1990]. Эти регуляторные влияния, проявляющиеся на всех уровнях - от генома до поведения, обеспечиваются нейроэндокринной системой, которая, в свою очередь, состоит из нескольких взаимосвязанных и взаиморегулируемых иерархических уровней; 1) ткани и ор- ганы-мише™; 2) периферические эндокринные железы; 3) гипофиз; 4) гипоталамус; 5) регуляторные системы гшюталамических нейроэндокринных центров (рис. 1) [Сентаготаи и др., 1965; Jackson, Mueller, 1982; Поленов, 1993].
Любая регуляторная реакция начинается с того, что информация от интеро- и экстерорецепторов, закодированная в виде нервных импульсов, достигает гипоталамуса, где она трансформируется нейросекреторными нейронами в сигнальные молекулы пептидных и моноаминовых нейрогормонов. Нейрогормоны выделяются из терминалей аксонов нейросекреторных нейронов в гуморальные среды - кровь или спинномозговую жидкость (ликвор) - в области так называемых циркумвентрикулярных органов (срединное возвышение или сосудистый орган терминальной пластины) или нейрогемальных органов (задняя доля гипофиза), где отсутствует гематоэнцефалический барьер (рис. 2). В срединном возвышении нейрогормоны поступают в гипоталамо-гипофизарную портальную систему циркуляции и достигают передней доли гипофиза, обеспечивая контроль аденогипофизарных функций, а опосредованно и функций висцеральных орга- нов-мишеней, в первую очередь периферических эндокринных желез. Часть нейрогормонов поступает в общую систему циркуляции в задней доле гипофиза и в сосудистом органе терминальной пластины, реализуя парааденогипофизарный путь регуляции висцеральных органов-мишеней [Jackson, Mueller, 1982; Поленов, 1993]. Нейрогормоны выделяются также
вжелудочки мозга, по которым они переносятся ликвором и влияют дистантно на нейроны мозга - мишени [Wood, 1982]. В свою очередь функциональная активность нейроэндокринных центров гипоталамуса регулируется гормонами периферических эндокринных желез и аденогипофиза, а также гипоталамическими нейрогормонами по принципу обратных связей.
Нейрон
> 3-й желудочек
Эпендима
. Срединное Г возвышение
Первичное
портальное
сплетение
капилляров
Аденогипофиз
Рис. 2. Схематическое изображение строения циркумвентрикулярного органа - срединного возвышения и его роли в нейроэндокринной регуляции по принципу прямых и обратных связей
Стрелками обозначен транспорт нейрогормонов, обеспечивающих прямую регуляцию железистых клеток аденогипофиза (/) и регуляцию перивентрикулярно расположенных нейронов по принципу ультракороткой обратной связи (2), а также транспорт гормонов аденогипофиза, обеспечивающих регуляцию нейроэндокринных центров мозга по принципу короткой обратной связи(3)
соответственно длинных, коротких и ультракоротких (см. рис. 1, 2). Гормоны периферических эндокринных желез также обеспечивают регуляцию аденогипофиза и ряда негипоталамических отделов мозга-мишеней по принципу обратной связи [Jackson, Mueller, 1982; Baulieu, Kelly, 1990].
Наряду с централизованной и высокоспециализированной нейроэндокринной системой существует региональная нейроэндокринная система,

роэндокринное дистантное действие в результате переноса регуляторных молекул жидкими средами организма (ликвор, кровь и др.): г) нейротрансмиттерное действие в области синаптических контактов; д) нейромодуляторное действие на рецепторы, распределенные по всей поверхности клетки, вне зон специализированных межклеточных контактов [Vincent, Simonnet, 1986]. Одной из наиболее важных особенностей клеток, синтезирующих нейрогормоны, является их высокая пластичность, что проявляется в переключении специфических синтезов при изменении функционального состояния организма. Оказалось, что экспрессия специфических генов и, следовательно, синтезов модулируется сигнальными молекулами межклеточной среды [Galas, 1994].
Существует ряд гипотез, пытающихся объяснить наличие идентичных специфических синтезов в различных по природе клетках, например в нейронах, железистых эндокринных клетках и иммунокомпетентных клетках. Согласно первоначальной APUD-концепции, нейроэндокринные клетки желудочно-кишечного тракта и головного мозга должны были иметь одних и тех же предшественников в фило- и онтогенезе [Pearse, 1969]. Сходная концепция Kobayashi [Fujita, Kobayashi, 1979] предполагает существование особой популяции так называемых паранейронов, обладающих свойствами как железистых клеток, так и нейронов.
Несмотря на определенный вклад существующих гипотез в понимание механизмов секреторного процесса, ни одна из них не учитывала того, что большинство биологически активных веществ (нейропептидов, нейротрансмиттеров, гормонов и др.) синтезируется не только у высших позвоночных, но и у беспозвоночных, включая простейших и бактерии. Другими словами, синтез биологически активных веществ, характерных для высших позвоночных, в фило- и онтогенезе появляется задолго до формирования высокоспециализированных эндокринной, нервной и иммунной систем [LeRoith, Roth, 1984]. Действрггельно, у одноклеточных эукариотов были обнаружены такие типы сигнальных молекул, как пептиды (инсулин, АКТГ, (3-эндорфин, соматостатин (СС) и др.), биогенные амины (катехоламины, серотонин), стероиды и т.д. Более того, у одноклеточных организмов регуляторные эффекты сигнальных молекул опосредованы через такие же, как и у высших позвоночных, специфические рецепторы [LeRoith, Roth, 1984]. На основании вышеизложенных фактов была сформулирована "уницеллюлярная" теория, которая позволила объяснить практически все противоречия предыдущих концепций о природе клеток, секретирующих биологически активные вещества. Согласно этой теории, все клетки являются первично секреторными и синтезирующими все типы сигнальных молекул, которые обеспечивают регуляцию как собственных функций, так и функций окружающих клеток-мишеней.
Возникновение многоклеточных организмов в филогенезе характеризуется появлением специализированных секреторных клеток, что создает новые возможности для межклеточных взаимоотношений и их регуляции. В нервной системе гипотетические секреторные клетки-предшественники превращаются в нейроны, синтезируемые ими сигнальные молекулы начинают играть роль нейротрансмиттеров, а межклеточная среда представлена синаптической щелью. В эндокринной системе секреторные клетки-
