

6 курс / Эндокринология / Механизмы_нейроэндокринной_регуляции_Угрюмов_М_В_1999
.pdfТирозингидроксилаза-иммунореактивные нейроны аркуатного ядра представляют собой наиболее позднюю популяцию. Действительно, максимальный прирост нейронов этого ядра наблюдается позднее, чем в зоне инсерта и в перивентрикулярном ядре. При этом у самок пик образования нейронов приходится на 14-й день, а у самцов - на 15-й день. Половые различия этим не Офаничиваются, так как у самцов, в отличие от самок, отмечены транзиторное снижение прироста нейронов на 13-й день и появление второго пика образования нейронов на 12-й день (см. рис. 36, в). Наличие двух пиков рассматривается как показатель существования двух популяций нейронов, экспрессирующих тирозингидроксилазу. В пользу этого предположения свидетельствует то, что, во-первых, образование однородных популяций нейронов характеризуется единственным пиком [Hoffman et al., 1980; Wray, 1989; Rodier et al., 1990], a, во-вторых, в аркуатном ядре взрослых животных тирозингидроксилаза экспрессируется не только в дофаминергических, но и в пептидергических нейронах [Everitl et al., 1992].
Приведенные выше данные позволили прийти к выводу о том, что образование нейронов, экспрессирующих тирозингидроксилазу, происходит в соответствии с дорсовентральным градиентом и что существует половой диморфизм в динамике образования этих нейронов. Половой диморфизм, по всей вероятности, запрограммирован генетически и не является результатом действия эстрогенов, образующихся в мозге из циркулирующих в крови андрогенов. Действительно, половые различия в уровне циркулирующего тестостерона у крыс проявляются не ранее, чем на 17-18-й день внутриутробного развития [Weisz, Ward, 1980; Baum et al., 1991], т.е. уже после образования этих нейронов.
О времени образования дофаминергических нейронов у зрелорождающихся животных, в частности у человека, судили по первому выявлению в них катехоламинов или ферментов их синтеза. В большинстве работ появление дофаминергических нейронов в гипоталамусе человека относят к 10-15-й неделе внутриутробного развития [Nobin, Bjorklund, 1973; Olson et al., 1973; Pearson et al., 1980; Picker et al., 1980], и лишь в единственном исследовании Vemey с соавт. [ 1991] тирозингидроксилаза-иммунореактивные нейроны обнаружены уже на 5-й неделе.
Динамика выселения дофаминергических нейронов в места их окончательной закладки в гипоталамусе была изучена с помо1Цью мономечения - гистофлуоресцентного метода, а также иммуногистохимии тирозингидроксилазы или декарбоксилазы ароматических L-аминокислот. Важно подчеркнуть, однако, что первоначальное выявление дофаминергических нейронов по специфическим маркерам в той или иной области развивающегося мозга может быть результатом не только их миграции в эту область, но и результатом экспрессии дофаминергического фенотипа нейронами, уже выселившимися в эту область. У незрелорождающихся животных (крысы) одиночные дофаминергические нейроны обнаружены в гипоталамусе сразу же после их образования - на 12-13-й день [Schlumpf et al., 1980; Specht et al., 1981a; Ugrumov et al., 1989d]. В течение последующих двух-четырех дней эти нейроны выселяются в дорсомедиальную область гипоталамуса, а затем и в преоптическую область, образуя там массивные скопления (рис. 37, А) fSchlumpf et al., 1980; Jaeger, 1986; Ugrumov et al., 1989d].
90
Рекомендовано к изучению сайтом МедУнивер - https://meduniver.com/
Основной процесс выселения нейронов в места их окончательной закладки происходит у незрелорождающихся животных в перинатальном периоде - у крыс начиная с 18-го дня внутриутробного развития (см. рис. 37, Б) [Ugrumov, 1994]. Первые нейроны появляются в сосудистом органе терминальной пластины, в области диагонального пучка Брока, в медиальной и латеральной преоптической областях, вдоль вентральной поверхности гипоталамуса, в оптических нервах и тракте, в паравентрикулярном ядре, в перивентрикулярном ядре, в супрахиазматическом ядре, в зоне инсерта, в дорсомедиальном ядре и в медиобазальном гипоталамусе (см. рис. 37, Б). В течение перинатального периода количество нейронов и область их распространения постепенно увеличиваются. Уже к концу пренатального периода образуются крупные скопления нейронов в перивентрикулярной области, в ретрохиазматической области, в зоне инсерта и в медиобазальном гипоталамусе [Jaeger, 1986; Ugrumov et al., 1989d; Borisova et al., 1991].
У зрелорождающихся млекопитающих выселение дофаминергических нейронов, по всей вероятности, осуществляется в основном в пренатальном периоде [Nobin, Bjorklund, 1973; Olson et al., 1973]. Так, у человека первое небольшое скопление дофаминергических нейронов появляется в инфундибулярном (аркуатном) ядре в течение третьего-четвертого месяца внутриутробного развития. К концу пренатального развития (7-9-й месяц) эти нейроны уже распределены таким же образом, как и у взрослых особей [Su et al., 1987], образуя четыре основных скопления. Первое - каудальное скопление нейронов (АН) распространяется от мезэнцефалона до мамиллярной области и занимает заднее гипоталамическое ядро. Второе - дорсальное скопление (А13) расположено в перивентрикулярной области гипоталамуса на всем ее протяжении в рострокаудальном направлении от переднего гипоталамуса до мамиллярных тел. Третье - наиболее многочисленное латеральное скопление занимает туберомамиллярное ядро и латеральное гипоталамическое ядро. Эта группа не имеет аналога у крыс, однако соответствует скоплению дофаминергических нейронов в латеральном гипоталамусе у кошек и кроликов [Parent et al., 1984]. Вентромедиальное скопление (А12) нейронов занимает область аркуатного и вентромедиального ядер. И, наконец, передняя группа (А 14) сосредоточена в переднем гипоталамусе [Su et al., 1987]. Особого внимания заслуживают данные Panayotacopulou и Swaab [1993] о том, что количество нейронов, экспрессирующих тирозингидроксилазу, в супраоптическом и паравентрикулярном ядрах достигает максимума у новорожденных в течение первых трех месяцев жизни, а затем в течение последующего года снижается. Вполне возможно, что в данном случае выявляются не дофаминергические, а вазопрессинергические нейроны, экспрессирующие тирозингидроксилазу в состоянии повышенной функциональной активности [Calas, 1994].
Дифференцировка дофаминергических нейронов
Морфология. Дофаминергические нейроны претерпевают существенные изменения в процессе дифференцировки [Ugrumov et al., 1989d]. Вновь образованные нейроны представляют собой мелкие овальные клетки с одним или двумя короткими неразветвленными отростками, заканчивающи-
91


мпо
лпо
Рис. 37 (продолжение)
в) довольно крупные биполярные нейроны, софедоточенные в перивентрикулярном ядре. Нейроны этих трех популяций, хотя и являются дофаминергическими, по-видимому, имеют разные клеточные предшественники, а следовательно, и генетические программы, о чем свидетельствуют различия в динамике их образования [Balan et al., 1996]. Региональные особенности морфогенеза дофаминергических нейронов могут быть также связаны с различиями в их микроокружении. Однако это предположение маловероятно, так как в диссоциированной культуре эмбрионального гипоталамуса выявлены такие же типы нейронов, как и in situ [Bennett, Morris, 1989; Murray, Gillies, 1993; Kapatos, Hirayama, 1994].
Эфферентные аксональные проекции. Использованные до настоящего времени методические подходы: иммуногистохимия одного из фермен-
94
Рекомендовано к изучению сайтом МедУнивер - https://meduniver.com/
/\^ж |
|
|
1/"• |
с о |
МП |
•• |
с > \ |
|
•-пев
Рис. 37 (окончание)
тов синтеза катехоламинов и гистофлуоресценция, в большинстве случаев не позволяют идентифицировать эфферентные проекции дофаминергических нейронов гипоталамуса на отделы-мишени мозга, а дают лишь общую картину иннервации гипоталамуса катехоламинергическими волокнами как гипоталамического, так и экстрагипоталамического происхождения.
У незрелорождающихся животных первые катехоламинергические волокна прорастают в гипоталамус из ствола мозга и мезэнцефалона [Spechl et al., 1981а] в составе медиального пучка переднего мозга в начале второй половины внутриутробного развития - у крыс начиная с 13-го дня [Jaeger, 1986; Ugramov el al., 1989e; Borisova et al., 1991]. Вскоре катехоламинергические волокна покидают медиальный пучок переднего мозга, прорастая в такие отделы-мишени мозга, как медиальная ретрохиазматическая об-
95
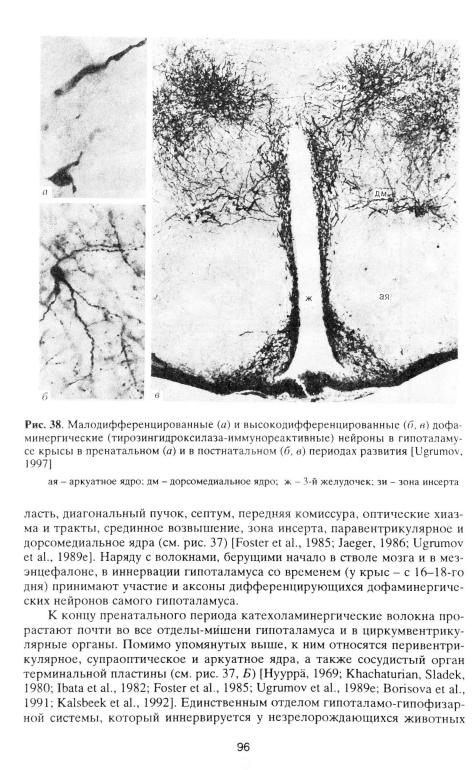

Рис. 39. Схематическое изображение синаптогенеза в супрахиазматическом ядре у крысы в пренатальном {а, ж) и постнатальном {б-е, з, и) периодах развития [Ugrumov et al., 1994а]
Пресинапсы и синапсы образованы тирозингидроксилаза-иммунореактивными телами нейронов и волокнами (точки) с иммунонегативными аксонами (а-е), а также тирозингидрок- силаза-иммунореактивными аксонами (точки) с иммунонегативными дендритами (ж-и). П - постнатальный день; Э - эмбриональный день. / - пресинапсы; 2 - симметричные синапсы; 3 - асимметричные синапсы
Рекомендовано к изучению сайтом МедУнивер - https://meduniver.com/