

2 курс / Нормальная физиология / Физиология.-Шукуров-Ф.А
.pdfПри проведении возбуждения по ЦНС отмечаются следующие особенности:
1)Одностороннее проведение возбуждения, так как в синапсах возбуждение передается только от пресинаптической мембраны (здесь выделяется медиатор при помощи которого происходит передача возбуждения в синапсах) к постсинаптической.
2)Суммация возбуждения (рис.34).
Рис.34. Последовательная и пространственная суммация. На данном рисунке отражается один из особенностей проведения возбуждения в центральной нервной системе (ЦНС) – суммация возбуждения. Различают два вида суммации: 1) последовательная, или временная суммация (А), которая возникает при действии ритмического раздражителя определенной частоты (F1). В данном случае в ЦНС происходит суммация выделяющегося медиатора в пресинаптической мембране. При очень маленькой частоте (1-5 Гц) раздражителя суммация не возникает, так как при этом до поступления очередного импульса, медиатор от предыдущего импульса диффундирует в синаптическую щель и разрушается; 2) пространственная, или одновременная суммация (Б), которая возникает при одновременном действии одиночного или ритмического раздражителя (F2) на два и более рецептора одного и того же рецептивного поля (совокупность рецепторов при раздражении которых возникает одна и та же реакция). При этом происходит суммация медиатора в синаптической щели.
81
Различают два вида суммаций: а) пространственная, или одновременная (рис.34 В). Этот вид суммации возникает благодаря тому, что рецепторов значительно больше, чем афферентных нейронов, поэтому из 2-х и более рецепторов сигналы поступают в один и тот же нейрон. Для того, чтобы убедиться в наличии данной суммации, необходимо подействовать на рецептор подпороговой силой – никакого ответа не будет. Затем подействовать одновременно двумя подпороговыми силами на два рецептора одного и того же рецептивного поля (совокупность рецепторов при раздражении которых происходит одна и та же реакция) – будет ответная реакция. При действии одной подпороговой силой выделяется мало медиатора и ВПСП на постсинаптической мембране не доходит до критического уровня, поэтому нет ответной реакции. При одновременном действии двух подпороговых сил медиатор выделяется одновременно в двух нервных окончаниях, которые заканчиваются на одном и том же нейроне. Таким образом, количество медиатора становится достаточным для того, чтобы ВПСП дошел до критического уровня;
б)последовательная суммация, или временная (рис.34 А).
Этот вид суммации происходит при действии ритмического раздражителя. При этом частота ритмического раздражителя должна быть достаточно высокой, чтобы медиатор, выделившийся на первый стимул не успел разрушиться и выделяется новая порция медиатора от последующих раздражений. Из выше изложенного следует, что при обоих видах суммаций происходит суммация медиатора. При пространственной (одновременной) суммац ии медиатор суммируется в синоптической щели. При последовательной (временной) суммации медиатор суммируется в пресинаптической мембране.
82
3)Замедление проведения возбуждения по ЦНС. Эта особенность обусловлена тем, что в ЦНС имеются много синапсов, а в каждом синапсе передача возбуждения осуществляется за счет медиатора – необходимо время для выделения медиатора, его диффузии через пресинаптическую мембрану и взаимодействие с реактивной субстанцией постсинаптической мембраны с возникновением ВПСП.
4)Конвергенция, то есть схождение импульсов к одному нейрону – можно сказать это следствие пространственной суммации (рис.34 В). Эта особенность обусловлена тем, что рецепторов значительно больше, чем нейронов, поэтому от нескольких рецепторов (одного и того же рецептивного поля) импульсы поступают к одному нейрону – это и есть конвергенция.
5)Окклюзия, закупорка – уменьшение величины ответной реакции при одновременном раздражении двух и более рецепторов по сравнению с суммарной реакцией, полученной при раздражении каждого рецептора в отдельности (рис.35).
Из рисунка видно, что при раздражении каждого рецептора (А, Б) в отдельности в ЦНС происходит возбуждение по 4 нейрона: при раздражении рецептора А возбуждаются нейроны 1,2,3,4, а при раздражении рецептора
Бвозбуждаются нейроны 3,4,5,6. Из данной схемы видно, что 3 и 4 нейроны в ЦНС являются общими для рецепторов А и Б. При одновременном раздражении в ЦНС возбуждаются не 8 нейронов, а 6. Нейроны 3 и 4 возбуждаются по конкурентному механизму либо за счет раздражения рецептора А, либо за счет раздражения рецептора Б. Если нейроны 3,4 возбуждаются за счет импульсов, возникающих при раздражении рецептора А, то определенное время эти
нейроны будут находиться в рефрактерной фазе
83

возбудимости и не реагируют на импульсы, возникающие при раздражении рецептора Б и наоборот. Таким образом, в результате окклюзии (закупорки импульсов, возникающих при раздражении рецептора А или Б) при одновременном раздражении рецепторов А и Б реакция эффекторов М1 и М2 будет меньше суммы реакции каждого эффектора при раздражении каждого рецептора в отдельности. Из выше изложенного видно, что окклюзия является следствием конвергенции.
Рис. 35. Схема возникновения окклюзии.
6)Дивергенция - иррадиация – расхождение импульсов
вЦНС от одного нейрона к другому (рис.36)
Из рисунка видно, что расхождение импульсов в ЦНС обусловлено наличием большого количества вставочных нейронов и многочисленных отростков. При раздражении рецептора 2 происходит распространение (иррадиация) возбуждения в ЦНС от одного нейрона (А) к другим (В,С,D,Е) и даже к тем нейронам, которые не относятся к данному рецептивному полю (2). При пороговом и субмаксимальном раздражении рецепторов 1, 2, 3 соответственно возбуждаются нейроны С, А, Е и реагируют эффекторы 4, 6 и 8. При действии максимального и сверхмаксимального раздражителя на рецептор 3 происходи возбуждение не только нейрона А, но В, С, D и Е через отростки а, в, с, d, е,
84

то есть происходит иррадиация возбуждения в ЦНС по многочисленным вставочным нейронам.
В результате иррадиации процессом возбуждения охватываются большое количество эффекторов (рабочих органов), и даже тех, которые не относятся к данному рецептивному полю (4-5; 7-8). Из выше изложенного следует, что иррадиация является следствием дивергенции.
Рис. 36. Схема возникновения дивергенции (иррадиации).
7) Последействие – сохранение возбуждения в ЦНС после прекращения раздражения (рис.37).
Различают два вида последействия: а) кратковременное последействие – его причиной является высокий уровень следовой деполяризации в нейронах ЦНС (рис.37 Б); б) длительное последействие – его причиной является циркуляция импульсов по коллатералям через вставочные нейроны (рис.37 А). Длительное последействие (А) возникает на действие максимального или сверхмаксимального раздражителя (F). При этом происходит длительная циркуляция импульса в ЦНС, через вставочный возбуждающий нейрон, поэтому при одиночном раздражении рецептора в ЦНС длительно сохраняется возбуждение и
85
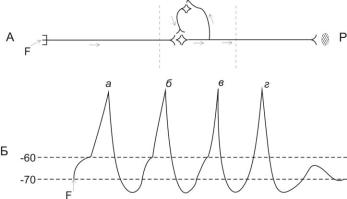
отмечается длительная ответная реакция эффектора (Р) после прекращения раздражения рецептора. Кратковременное последействие (Б) может возникнуть при действии порогового раздражителя (F). Это связано с особенностью возникновения МПД в нейронах и с низким порогом раздражения: разница между МПП (-70 мв) и критическим уровнем деполяризации (-60 мв). При возникновении МПД в нейронах возникает высокий уровень следовой деполяризации (d, d1, d2), поэтому одиночное раздражение вызывает 4 реакции: реакция а в ответ на действие раздражителя, ответы б, в, г – это последействие – реакция на высокий уровень следовой деполяризации.
Рис. 32. Схема возникновения кратковременного и длительного последействия.
8) Трансформация ритма – изменение количества импульсов возбуждения, выходящих из нервного центра, по сравнению с числом импульсов, приходящих к нему. Различают два вида трансформации: а) понижающая трансформация при этом на несколько возбуждений, пришедших к нейрону возникает только одно возбуждение – в основе этой трансформации лежит суммация возбуждения
86
в ЦНС; б) повышающая трансформация – при этом увеличивается количество импульсов возбуждения – в основе этой трансформации лежит последействие.
Нейроны как и синапсы обладают быстрой утомляемостью, а нервные волокна практически не утомляемые.
Нейроны как и синапсы обладают низкой лабильностью и в них легко возникает процесс торможения.
Нервные центры обладают тонусом, поэтому даже при отсутствии специальных раздражений они посылают импульсы к рабочим органам.
ПРИНЦИПЫ КООРДИНАЦИОННОЙ ДЕЯТЕЛЬНОСТИ В ЦНС
Координация – это объединение действия в единое целое, объединение различныхнейронов в единый функциональный ансамбль, решающий конкретную задачу. Благодаря координации осуществляется согласованная деятельность всех органов и систем организма. Выделяют следующие принципы координационной деятельности ЦНС:
Принцип доминанты - был открыт А.А. Ухтомским. Изучая ответы скелетной мышцы кошки на электрические раздражения коры больших полушарий, он обнаружил, что при акте дефикации ответы мышцы прекращаются. Проанализировав этот факт А.А. Ухтомский пришел к мнению о наличии в ЦНС явления доминанты – господствующего очага возбуждения. Дело в том, что среди многочисленных рефлекторных актов, которые могут быть выполнены в данный момент, осуществляются те реакции, которые наиболее значимы для организма в данный момент времени. Центры, участвующие в реализации более значимых для
87

организма реакций, А.А. Ухтомский назвал «доминантным очагом возбуждения». Этот «очаг» обладает рядом важных свойств: а) повышенной возбудимостью; б) стойкостью возбуждения (инертностью), поэтому этот очаг возбуждения трудно подавить другим очагом возбуждения; в) способностью к суммации субдоминантных возбуждений, то есть доминантный очаг возбуждения притягивает к себе другие очаги возбуждения, менее значимые для организма в данный момент времени; г) способностью тормозит ь субдоминантные очаги возбуждения.
Принцип обратной связи – обеспечивает связь выхода системы с ее входом (рис.38).
1
Рис.38 Схема обратной связи. Импульсы от рецептора (Р) по афферентному пути (2) поступают в ЦНС (3), затем по эфферентным путям (4) к эффектору (5 – Э), возникает результат (Р). Результат работы эффектора по обратной связи (6) поступает в ЦНС либо от рецепторов эффектора (а), либо от рецептора, воспринимающего результат (б).
ЦНС посылает к рабочим органам определенные команды (каким должен быть результат). Фактический результат работы органа несколько отличается от должного. Разница фактического и должного результата называется рассогласованием: чем больше рассо гласование, тем менее оптимальными являются процессы управления. Обратная связь несет информацию в ЦНС о параметрах фактического результата и в ЦНС осуществляется сравнение фактических параметров с теми,
88
которые должны быть в данных условиях. Таким образом, обратная связь несет информацию в ЦНС о фактическом результате. Здесь происходит сравнение фактического результата с заданным (должным) результатом. Если фактический результат больше заданного уменьшается функция эффектора (отрицательная обратная связь), если фактический результат меньше заданного – усиливается функция эффектора (положительная обратная связь). Из вышеизложенного следует, что благодаря сигналам по обратной связи происходит коррекция (исправление) командных сигналов от ЦНС, чтобы приблизить параметры фактического результата к должным параметрам – в результате уменьшается рассогласование и происходит оптимизация процессов управления. При положительной обратной происходит усиление параметров результата, а при отрицательной - уменьшение. В биологических системах положительная обратная связь реализуется в основном в патологических ситуациях. Отрицательная обратная связь улучшает устойчивость системы, то есть способность возвращаться к первоначальному состоянию после прекращения влияния возмущающих факторов (воздействий на систему, при которых происходит увеличение рассогласования).
Принцип реципрокности (сочетанности,
сопряженности, взаимоисключения). Этот принцип будет рассмотрен ниже в разделе реципрокного торможения.
Принцип реципрокности играет важную роль в автоматической координации двигательных актов.
Принцип общего конечного пути. Эффекторные нейроны ЦНС (прежде всего мотонейроны спинного мозга) являются конечными в цепочке состоящей из афферентных, промежуточных и эффекторных нейронов. Эти мотонейроны могут вовлекаться в осуществление различных реакций организма возбуждениями, приходящими к ним от большого
89

числа афферентных и промежуточных нейронов, для которых они являются конечным путем (от ЦНС к эффектору). Например, на мотонейронах спинного мозга, иннервирующих мышцы конечностей, оканчиваются волокна афферентных нейронов, нейронов пирамидного тракта, ядер мозжечка, ретикулярной формации мозга и др. (рис. 39).
Рис. 39. Схема принципа конечного пути.
Таким образом, эти нейроны, обеспечивающие рефлекторную деятельность конечности, рассматриваются как конечный путь для общей реализации на конечность многих нервных влияний. Из рисунка видно, что альфа мотонейрон (1) одновременно поступают импулься от мозжечка (М), коры больших полушарий (К) и рецептора (Р). При этом выбор конечного пути может осуществляться двумя способами: 1) по конкурентному принципу (А), то есть происходит одна ответная реакция, наиболее важная путем торможения других; 2) по союзному механизму (Б), то есть отмечаются все три реакции взаимно усиливая друг друга.
Формирование конечного пути осуществляется двумя механизмами: а) конкурентный механизм – при этом реализутся какое-то одно влияние (более значимое для организма в данный момент) на мотонейроны за счет торможения других воздействий; б) союзный механизм – при
90