

2 курс / Нормальная физиология / Физиология.-Шукуров-Ф.А
.pdf…-… – выход … … из … – изменение … … – уход … в … … нитей – соединение головки … … с … – образование … комплекса.
5.При сокращении мышц отмечается … разрыв … …, что … к … … вдоль
…Состояние …, или … окоченение мышц возникает при … … головки
…… к …
Задачи
1.При раздражении нерва мышца доведена до утомления. Что произойдет, если в это время наносить раздражение непосредственно на мышцу. Показать на схеме и объяснить результат
2.Изобразите графическое изменение ответной реакции мышцы, если на нее действовать пороговой, пороговой и максимальной силой при следующих состояниях мембраны, взяв исходную ответную реакцию в состоянии поляризации: в середине порога деполяризации, периода деполяризации и реполяризации пика МПД, следовой деполяризации и на вершине следовой гиперполяризации.
3.В медицинской практике иногда пропускают через мышечную ткань токи высокой частоты (100000Гц) и мощностью около 200Вт. Хронаксия
мышцы, которая подвергается действию этого тока равна 0,1 мс. Будет ли в этих условиях сокращаться мышца. Объясните результат.
4. При миастении, которая хактеризуется патологической слабостью и утомляемостью скелетных мышц, уменьшается вероятность взаимодействия медиатора ацетилхолина с рецепторами постсинаптической мембраны нервно-мышечных синапсов. Причина заключается в том, что у больных появляются антитела к этим рецепторам, которые блокируют их и способствуют их разрушению. Основываясь на ваших знаниях механизмов передачи в нервномышечном синапсе, предложите виды лекарств (по характеру действия), которые в этих условиях могли бы улучшить синаптическую передачу.
ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА НЕРВОВ. МИОНЕВРАЛЬНЫЙ СИНАПС
Нерв и нервные стволы, которые можно видеть невооруженным глазом – есть совокупность аксонов, окруженные общей соединительно-тканной оболочкой.
51

В центральной нервной системе (ЦНС) аксон окружают клетки, в совокупности называемые нейроглией; за пределами ЦНС аксон заключен в оболочку из швановских клеток – неврилемма. Аксон оканчивается группой концевых разветвлений, которые служат для передачи сигналов другим нейронам или исполнительным органам. Вокруг некоторых аксонов швановские клетки откладывают спиральные слои изолирующего материала – миелин. Миелин имеет липидную природу и выполняет следующие основные функции: 1) является изолятором, в результате чего препятствует прохождению ионов и возникновению местных токов на участках нерва, покрытых миелином; 2) выполняет трофическую функцию: регулирует обмен веществ, регулирует рост осевого цилиндра, принимает участие в синтезе ацетилхолина. Благодаря наличию или отсутствию миелина все нервы по своей структуре делятся на: миелиновые и безмиелиновые (рис.23)
Рис.23 Классификация нервов по структуре. Все нервы
(совокупность отростков нейрона) по своей структуре делятся на две группы: 1) безмиелиновые нервы, или безмякотные (А); 2) миелиновые нервы (Б), или мякотные. Безмиелиновые нервы состоят и осевого цилиндра (1 – совокупность отростков) и Швановской оболочки (2). Миелиновые нервы состоят из: 1) осевого цилиндра (1); 2) миелина (2) – липид, выполняющий функцию изолятора, а также участвующего в синтезе медиатора; 3) перехваты Ранвье (3) – участок нерва не покрытого миелином; 4) Швановская оболочка (4).
Вокруг аксонов, находящихя в ЦНС, миелин образуется не швановскими клетками, а особыми клетками сателлитами
– олигодендроцитами. Нервы, не имеющие миелин,
52

называются безмиелиновыми. Таким образом, по своей структуре нервы бывают миелиновые и безмиелиновые.
Каждый нерв обладает двумя физиологическими свойствами:
возбудимостью и проводимостью
Механизм распространения возбуждения по нерву. Распространение возбуждения по нерву объясняется теорией местных и круговых токов (рис.24).
Рис.24. Механизм передачи возбуждения по нервам. На схеме отражается механизм передачи возбуждения в миелиновых (Б) и безмиелиновых (А) нервах. При действии раздражителя на безмиелиновый нерв (А2) происходит деполяризация мембраны в результате чего возникают местные токи (а,б) и происходит деполяризация мембраны соседних участков слева и справа, возбуждение распространяется в обе стороны (двухстороннее распространение возбуждения). В данном случае возбуждение распространяется по всей поверхности мембраны, поэтому скорость передачи возбуждения в безмиелиновых нервах очень низкая (0,5 – 10 м/с). При действии раздражителя на миелиновый нерв (Б2) происходит деполяризация мембраны в результате чего возникают круговые токи (а,б) между перехватами Ранвье слева и справа и происходит деполяризация мембраны этих перехватов, возбуждение распространяется в обе стороны (двухстороннее распространение возбуждения). В данном случае возбуждение распространяется от одного перехвата Ранвье к другому или сразу к третьему, поэтому скорость передачи возбуждения в миелиновых нервах очень высокая (до 70 – 120 м/с).
При любом раздражении нерва (электрическим током, прикосновением, каким-либо повреждающим фактором и т.д.) происходит изменение МПП. До раздражения нерв поляризован, а при стимуляции нерва он деполяризуется. Если при этом приложить к поверхности нерва два электрода, из которых один будет находиться в точке
53
раздражения, а другой на участке, не подвергнутом раздражению, то прибор покажет, что первый электрод заряжен отрицательно по отношению ко второму. Такая местная деполяризация является начальным этапом возникновения нервного импульса. Этот местный процесс развивается медленно и, если он не достигнет критической величины, то , происходящие в нерве восстановительные процессы приводят к реполяризации мембраны нерва и изменения останутся чисто локальными (местными, не распространяющимися). Если деполяризация достаточно быстро достигает критического уровня, то она сама вызовет раздражение соседних участков нерва по обе стороны от себя. Таким образом, в результате местной стимуляции, возникает электрический ток, направленный от двух неактивных пока участков к деполяризованному участку. В результате этого местного тока происходит деполяризация двух новых участков нерва, находящихся по обе стороны от места раздражения. Эти два новых деполяризованных участка начнут стимулировать примыкающие к ним неактивные области и т.д., то есть возникает цепная реакция. Таким образом, это «самопроизводящееся» состояние деполяризации будет распространяться по нервному волокну. Такую распространяющуюся деполяризацию называют МПД нерва, который возникает быстро (его длительность около 2 мс). После каждого МПД должен пройти определенное время, прежде, чем сможет возникнуть второй МПД. Этот промежуток называется абсолютным рефрактерным периодом (от 0,2 до 2 мс). Механизм передачи возбуждения по нерву будет зависеть от его структуры. В безмиелиновом нервном волокне происходит
непрерывное распространение возбуждения, поэтому местные токи проходят через большую поверхность мембраны. В миелиновых нервах участки, покрытые миелином обладают очень высоким сопротивлением (100000
54
– 160000 ом/см2). Таким образом, строение миелинового нерва создает предпосылки для качественно нового типа проведения возбуждения – сальтаторного (от латинского persaltum – прыжки). Местные токи, в одном из перехватов не проходят через всю мембрану (как это отмечалось в безмиелиновых нервах), а идут через соседние перехваты, то есть, возникший МПД в участке раздражения, как бы перескакивают («прыгают») от одного перехвата Ранвье к другому. При этом МПД может перескакивать через 2-3 и даже 5 перехватов Ранвье. Нетрудно убедиться в преимуществе сальтаторного типа проведения возбуждения по нерву: 1) при этом увеличивается скорость распространения возбуждения (до 120 м/с); 2) распространение возбуждения происходит с меньшими затратами энергии, то есть более экономно.
Скорость проведения импульсов возрастает с увеличением диаметра аксона, так как при этом снижается внутреннее сопротивление. Таким образом, толстые нервы проводят возбуждение быстрее, чем тонкие. Однако у позвоночных высокие скорости достигаются иным путем. Важную роль при этом играет миелиновая оболочка, которая служит хорошим изолятором, так что прохождение тока между жидкостью, окружающей эту оболочку, и внутренней средой аксона, невозможно. Местные токи могут проходить только в перехватах Ранвье. Миелинизированное нервное волокно толщиной всего лишь в несколько микрон способно проводить импульсы со скоростью до 100 м/с, тогда как у самых толстых волокон (1 мм в диаметре), не покрытых миелином, скорость проведения составляет лишь 20 – 50 м/с. Если же брать только миелиновые нервные волокна, то здесь отмечается четкая зависимость скорости проведения возбуждения от диаметра нервных волокон.
Законы проведения возбуждения по нерву (рис.25, 26, 27). Различают три закона проведения возбуждения по
55

нерву. Первый закон свидетельствует о том, что возбуждение распространяется по нерву только в случае его
анатомической и физиологической целостности (рис.25).
Рис.25 Закон анатомической и физиологической целостности нерва. При раздражении нерва (1) с анатомической и физиологической целостностью возникает ответная реакция (4). При нарушении анатомической (3) или физиологической (2) целостности раздражение нерва (1б, 1в) не вызывает ответную реакцию (5, 6).
Так, если перерезать нерв (3б - нарушить анатомическую целостность), то возбуждение по нерву распространяться не будет. Если каким-либо повреждающим агентом (хлороформ, эфир, раствор KCl) подействовать на нерв, мы нарушаем временно физиологическую целостность нерва (2в). В этом случае возбуждение также не будет распространяться по нерву.
Второй закон свидетельствует о том, что возбуждение распространяется по нерву в обе стороны от места раздражения (рис.26), то есть в нервах отмечается двухстороннее проведение возбуждения.
Третий закон свидетельствует об изолированном проведении возбуждения по нерву, то есть возбуждение с одного нерва не передается на другой нерв как бы близко не находились нервные волокна (рис.27).
56

Рис. 26. Закон двухстороннего проведения возбуждения по нерву.
При раздражении нерва (2) импульсы идут в двух направлениях, о чем свидетельствует регистрация мембранного потенциала действия (3) слева и справа от места раздражения
Рис. 27. Закон изолированного проведения возбуждения по нерву.
Эфферентные нервы 1 и 2 соответственно заканчиваются на эффекторах 4 и 5. На протяжении А-Б эфферентные нервы 1 и 2 идут рядом. При раздражении рецептора 3 импульсы по эфферентному нерву идут до эффектора 5, мышца сокращается (6). Импульсы от эфферентного нерва 2 на участке А-Б не переходят на эфферентный нерв 1, поэтому эффектор 4 не реагирует (7) на раздражение рецептора 3.
Следует отметить, что закон физиологической целостности нерва в какой-то мере относителен для миелиновых волокон, в которых возбуждение распространяется скачкообразно, поэтому при небольшом участке альтерации нерва возбуждение может перейти этот участок. Закон изолированного проведения возбуждения относителен для безмиелиновых нервов, так как при сильном
57
возбуждении оно может перейти с одного безмиелинового нерва на другой безмиелиновый нерв
Механизм передачи возбуждения через синапс.
Нервная система состоит из отдельных нейронов, разделенных межклеточными промежутками, поэтому должен существовать механизм, который позволяет нервному импульсу «перескакивать» с аксона одного нейрона на дендриты или тела другого или, в случае нервномышечного соединения - на мышцу.
Вистории изучения передачи возбуждения с нерва на мышцу почетное место принадлежит физиологу Александру Григорьевичу Геницинскому. Он отдал изучению этой проблемы 15 лет своей исключительно плодотворной жизни.
Впериод 1935 – 1950 гг. был большой научный спор между сторонниками двух теорий синаптического проведения возбуждения в нервно-мышечном соединении – электрической и химической. И только в 1949 г на Пражском симпозиуме по свидетельству Экклса «почти всеми было признано, что передача импульсов в нервно-мышечном соединении и в ганглиях осуществляется посредством ацетилхолина».
Вклассическом эксперименте Отто Леви в 1921 г. с двумя сердцами, было наглядно продемонстрированно, что нервы при раздражении могут выделять какое-то, способное к диффузии, вещество. Когда два изолированных сердца соединяли таким образом, что кровь, выходившая из одного сердца, поступало в другое . при раздражении блуждающего нерва одного сердца останавливались оба сердца. Позже было установлено, что диффундирующее вещество представляет собой ацетилхолин.
Существующие межнейронные соединения Шеррингтон назвал синапсами. Синапс – это структурное образование, где происходит переход одного нервного волокна на другой, или переход нерва на нейрон и мышцу (рис. 28).
58
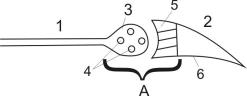
Рис. 28. Мионевральный синапс. На данной схеме отражается структура мионеврального синапса (А), через который осуществляяется передача возбуждения от нерва (1) на мышцу (2). Мионевральный синапс состоит из: 1) пресинаптической мембраны (3), в которой находятся пузырьки (4) заполненные физиологически активным веществом (медиатор); 2) постсинаптической мембраны (5), которая отличается от внесинаптической мембраны (6) наличием реактивной субстанции, обладающей высокой чувствительностью к медиатору; 3) синаптической щели между пре- и постсинаптической мембраной.
Для синаптического участка аксона (3) характерно скопление мелких округлых телец – синаптических пузырьков (везикул) диаметром от 10 до 20 нм. Эти пузырьки содержат специфическое вещество, которое освобождается при возбуждении аксона и называется медиатором. Окончание аксона с пузырьками называется пресинаптической мембраной. Участок нерва, нейрона или мышцы, куда непосредственно передается возбуждение называется
постсинаптической мембраной (5). Между этими двумя структурами имеется небольшой промежуток (не более 50 нм), который называется синаптической щелью. Таким образом, любой синапс состоит из трех частей:
пресинаптической мембраны, синаптической щели и постсинаптической мембраны. Из вышеизложенного следует, что в синапсах передача возбуждения осуществляется химическим способом и происходит это за счет трех процессов: 1) освобождения медиатора из пузырьков; 2) диффузии медиатора в синаптическую щель; 3) соединением этого медиатора со специфическими
59

реактивными структурами постсинаптической мембраны, что приводит к образованию нового импульса.
При каждом нервном импульсе происходит освобождение медиатора из пузырьков, который диффундирует в синаптическую щель и соединяется с реактивными структурами (субстанциями, рецепторами) постсинаптической мембраны и при этом возникает усиление проницаемости для ионов натрия или ионов калия (рис.29)
Рис. 29. Возникновение возбуждающего (ВПСП) и тормозного (ТПСП) постсенаптического потенциала. Разница между МПП (-80) и
КУД (-60) определяет возбудимость постсенаптической мембраны: чем больше эта разница (порог деполяризации), тем меньше возбудимость. Порог деполяризации в состоянии покоя 9А) равен 20 mV.
В первом случае за счёт возбуждающего медиатора увеличивается проницаемость для ионов натрия и происходит деполяризация постсинаптической мембраны с образованием возбуждающего постсинаптического потенциала (ВПСП). Если ВПСП не доходит до критического уровня (Б), возникает локальный ответ. Если ВПСП доходит до критического уровня (это зависит от количества медиатора), тогда во внесинаптической мембране возникает МПД. Во втором случае за счет тормозоного медиатора увеличивается проницаемость для ионов калия и происходит гиперполяризация (Г) мембраны с образованием
60