

35269707
.pdfРАЗДЕЛ 2. Вопросы источниковедения и историографии истории Улуса Джучи |
231 |
|
|
материалы султанского архива, среди которых находятся послания Ахмед-хана от 1477 г. к султану Мухаммеду (2, б. 108–109)», – дип уза.
С.Е. Малов кыска, лəкин югары бəя биреп, «А. Курат использовал в своей книге литературу на всех языках: русском, восточных и западноевропейских», дигəн (3, б. 188). Күрсəтелгəн бəялəмə Ə.Н. Куратның 1940 елда чыккан ярлыклар вə битеклəр китабына карата гомум бəялəмə булып, Алтын Урда ханнарының битеклəренə дə карый (12, б. 6–50).
М.Г. Сəфəргалиев тə «недавно были обнаружены в султанском архиве в Константинополе ярлыки последних ханов – Улук-Мухаммеда (1427 г.), Махмуда (1466 г.), Ахмета (1477 г.), опубликованные в 1940 г. Акдас Нигмет Куратом». (4, б. 287) дигəн югары бəясен бирə.
Ə.Н. Курат бастырып чыгарган битеклəрне тикшереп өйрəнгəн Т.Н. Солтанов «В архиве музея Топкапы в Стамбуле под № 10202 хранится письмо золотоордынского хана УлугМухаммада турецкому султану Мураду II, написанное в марте 1428 г. Этот важный исторический документ, имеющий большое значение для понимания событий, связанных с борьбой за золотоордынский престол в 20-х гг. XV в., был опубликован … Акдес Нимет Куратом (в 1937 г. и вторично в 1940 г.)» (5, б. 53) дип бəяли.
Икенче бер язмасында Мəхмүт хан битеген тикшергəндə Т.И. Солтанов «письмо Махмутхана впервые было введено в научный оборот ... А.Н. Куратом, опубликовавшим в 1940 г. Фотокопию, транскрипцию, текст и перевод письма ... с комментариями. В русском переводе письмо еще не публиковалось. Хотя оно знакомо советским востоковедам» (5а, б. 240) дип Ə.Н. Курат хезмəтлəренə югары бəя бирə.
М.Г. Госманов тарафыннан Җүчи Олысы тарихына караган Ə.Н. Курат хезмəтлəре чит иллəрдə басылып чыккан хезмəтлəр арасында «фундаменталь əһəмияткə ия булган тикшеренүлəр» сыйфатында югары бəялəнə (6, б. 23).
В.Х. Хаков Олуг Мөхəммəт ханның эпистоляр мирасы – төрек солтаны Морат IIгə язган рəсми-дипломатик хатының «беренче мəртəбə... Курат тарафыннан төзелгəн тарихи язмалар җыентыгында басылып» соңыннан башка хезмəтлəрдə дə дөнья күрүен яза (7, б. 139).
И.Л. Измайлов татар тарих белеме үсешен өйрəнү өлкəсендə совет хакимияте чорында өзелгəн Ш. Мəрҗани традициялəрен Һиҗрəттə дəвам иткəн Ə.Н. Куратның Җүчи Олысы тарихын «гомүмтөрки тарихы кысаларында» өйрəнүен югары бəяли (8, б. 40), «чыганаклар белəн эш итəргə менə дигəн əзерлекле» һəм «тарих өлкəсендə зур гыйлем иясе» булуына күрсəтə. (8а).
И.М. Миргалиев тарафыннан Ə. Н. Курат хезмəтлəре төрек тарихчыларының Җүчи Олысы тарихына караган хезмəтлəре арасында иң «көчле һəм ышанычлы» хезмəт сыйфатында бəялəнə
(10, б. 21).
Р.Ю. Почекаев чыганакларны тикшереп өйрəнүче «белгечлəрнең хезмəтлəрен аерып күрсəтеп», хан ярлыкларын тикшереп өйрəнүче хезмəтлəрдəн Ə.Н. Куратның китабын 1940 атый (11,
б. 11).
Г.В. Вернадский (1953), Г.А. Федоров-Давыдов (1973) шулай ук Ə.Н. Куратның Алтын Урда тарихына караган хезмəтлəренə югары бəя бирəлəр.
1.Греков, Б.Д., Якубовский А.Ю. Золотая Орда и ее падение. – М.: Богородский Печатник, 1998; беренче басмасы 1950 ел.
2.Базилевич К.В. Внешняя политика русского централизованного государства, вторая половина ХV в.
–М.: Изд-во Московского университета, 1952. – 544 с.
3.Малов С.Е. Изучение ярлыков и восточных грамот // Академику Владимиру Александровичу Гордлевскому к его семидесятипятилетию Сборник статей. – М.: Изд-во Академии наук СССР, 1953. – С. 187– 196 с.
4.Сафаргалиев М.Г. Распад Золотой Орды // На стыке континентов и цивилизаций… – М.: «Инсан», 1996. – С. 277–526. (Сарфаргалиев М.Г. Распад Золотой Орды. – Саранск: Мордовское книжное издательст-
во, 1960).
5.Султанов Т.И. Письмо золотоордынского хана Улуг-Мухаммада турецкому султану Мураду II // Тюркологический сборник, 1973. – М.: Изд-во «Наука», 1975. – С. 53–61.
5а. Султанов Т.И. Письма золотоордынских ханов // Тюркологический сборник, 1975. – М.: Изд-во
«Наука», 1978. – С. 235–251.
6.Госманов М. Мирас һəм бурычлар // Миркасыйм Госманов. Гасырдан – гасырга. – Казан: Татарстан китап нəшрияты, 2004. – Б. 20–28; беренче басмасы: Татарстан. 1993. № 6.
7.Хаков В. Казан ханлыгы чорында əдəби тел // Мирас. 1993. № 8. – Б. 132–140.
232 |
ЗОЛОТООРДЫНСКОЕ НАСЛЕДИЕ. Выпуск 2. 2011 |
|
|
8.Измайлов И.Л. Улус Джучи: взгляд на историю средневековой империи // Татарстан. 1993. №7. –
Б. 39–48.
8а. Измайлов И.Л. История изучения Улуса Джучи в отечественной и зарубежной науке // История татар. – Т. III. Улус Джучи (Золотая Орда). XIII – середина XV в. – Казань, 2009.
9.Асланапа Октай. Ататөрекнең университет реформасында казанлы остазларыбыз // Мəгариф. 2001.
№12 – Б. 75–77.
10.Миргалеев И.М. Политическая история Золотой Орды периода правления Токтамыш-хана. – Ка-
зань: Алма–Лит, 2003. – 164с.
11.Почекаев Р.Ю. Право Золотой Орды. – Казань: Изд-во «Фэн» АН РТ, 2009. – 260 с.
12.Kurat Akdes Nimet. Topkapı Sarayı Müzesi Arşivindeki Altın Ordu, Kırım ve Türkistan Hanlarına ait yarlık ve bitikler. – Istanbul: Burhaneddın Matbaası, 1940. – 212 s.
13.Kurat Akdes Nimet. Rusya Tarihi Başlangıςtan 1917’ye кadar. – Ankara: Türk Tarıh Kurumu basımevi, 1999. – 537 s.; I басмасы 1948 ел.
14.Kurat Akdes Nimet. IV–XVIII yüzyıllarda Karadeniz Kuzeindeki Türk Kavimleri ve Devletleri. – Ankara: Türk Tarih Kurumu Basımevi, Sayı: 182. Universıtesi Dil ve Tarih – Coğrafya Fakültesi yayınları, 1972. – 511 s.
15.Kurat A.N. Uluğ – Muhammed Han (?–1445) // Хамидуллин Б.Л. Народы Казанского ханства. – Казань: Татарское книжное издательство, 2002. – С. 286–298; I басмасы 1940 ел.
16.Kurat Akdes Nimet. Altinordu Devleti. – Ankara: Ayylldlz Matbaasi, 1976. – S. 926–932. (электронную версию см.: http://www.vatankirim.net/yazi.asp?YasiNo=43) (дата обращения: 08.04.2011).
17.Tahiroglu A.N. Orta Zaman Tarihi icin kısa bir Bibliyografya. – Istanbul: Burhaneddın Matbaası, 1934. – 69 s.
РЕЗЮМЕ
Ф.Х. Шарафутдинов
История Золотой Орды в трудах А.Н. Курата
А.Н. Курат специализировался в области истории народов Восточной Европы. Труды основаны на архивных, археологических и эпистолярных источниках. Выпустил книгу – сборник писем правителей Золотой Орды, Крыма, Туркестана, которыми тюркологи широко пользуются в своих трудах. В книгу включены письма Улуг Мухаммада (1428), Махмуд-хана (1466), Ахмад-хана (1477), которые А.Н. Курат извлек из архивов и ввел впервые в научный оборот. Его труд «IV–XVIII yuzyιllarda Karadeniz Kuzeyindeki Turk Kavimleri ve Devletleri» состоит из описания истории тюрков, начиная с искет-сарматов по XVIII в. Oчерк «Altιnordu Devleti», изданный в 1976 г. является кратким содержанием главы из этого его труда. «Rusya Tarihi» включает в себя Золотоордынский период с подробным описанием эпохи Джучиева улуса. Oчерк «Ulug Muhammed Han (?–1445)» посвящен описанию правления этого хана, параллельно отображая борьбу за престол в поздний период существования Джучиева Улуса, также подробному описанию родословной Улуг Мухаммада.
Труды А.Н. Курата в области изучения истории Джучиева Улуса и ярлыки и письма ханов Улуса Джучи обнаруженные в архивах и опубликованные им высоко оценены отечественными исследователями: А.Ю. Якубовским (1), К.В. Базилевичем (2), С.Е. Маловым (3), М.Г. Сафаргалиевым (4), Т.И. Султановым (5), (5а); М.А. Усмановым (6), В.Х. Хаковым (7); И.Л. Измайловым (8), (8а); И.М. Миргалеевым (10), Р.Ю. Почекаевым (11) и др.
ABSTRACT
Fazil Sharafutdinov
The Golden Horde History in A.N. Kurat works
In the article post-graduate student of the History Institute of the Academy of sciences of the Republic of Tatarstan F.Kh.Sarafutdinov gives briefly description A.N. Kurat works dedicate of the Golden Horde history and criticis comparative analysis of condition research in district of the Golden Horde history.
РАЗДЕЛ 2. Вопросы источниковедения и историографии истории Улуса Джучи |
233 |
|
|
О.А. Кравцова, И.Р. Газимзянов, И.М. Миргалеев
Генетический облик поволжских татар
Среднее Поволжье занимает территорию, которая по своим природно-климатическим особенностям является пограничной между лесом и степью, а в историческом плане делит Восток и Запад. Выгодное географическое положение и богатые запасы биоресурсов делали Среднее Поволжье контактной зоной активного этногенетического взаимодействия между народами, различающимися по происхождению, языку и культуре. Справедливо считается, что Среднее Поволжье
– это колыбель, где проходила история формирования современных этносов, проживающих здесь: татары, чуваши, мордва, марийцы и т.д.
Формирование тюркских племен, к которым относятся татары – второй по численности народ РФ, по археологическим данным, проходило в эпоху неолита и бронзы в результате сложного смешения представителей европеоидной и монголоидной рас. Вопросы происхождения татарского народа, несмотря на более, чем столетнее изучение, усложнены тем, что его этноним – татары – нередко служит определяющим при выяснении проблемы.
По традиции, роль важнейшей дисциплины, призванной выявлять и анализировать источники по ранним периодам истории этносов, играет археология. В процессе реконструкции исторических событий немаловажную роль играют и данные о биологическом разнообразии популяций человека, которое является конечным результатом формирования определенной этнической группы. К числу таких подходов относится и анализ структуры генофонда популяции на основе вариабельности ДНК – носителя наследственной информации.
Вданной работе на основании данных о генетическом разнообразии (полиморфизме) маркеров ДНК изучена структура генофонда современных популяций поволжских татар, представленных двумя этнографическими группами: казанскими татарами и татарами-мишарями из Азнакаевского и Буинского районов Республики Татарстан соответственно [1].
Все маркеры ДНК с позиций популяционных исследований можно разделить на три группы: маркеры митохондриальной ДНК, аутосомные полиморфные локусы (мини- и микросателлиты) и маркеры Y-хромосомы. Популяционный полиморфизм каждой группы определяется факторами микроэволюции: миграцией, генетическим дрейфом, мутациями, однако характер их вариабельности по-разному отражает действие и результат этих процессов. Основной особенностью полиморфизма митохондриальной ДНК является отсутствие рекомбинации и материнский тип наследования. Y-хромосомный полиморфизм является комплементарным митохондриальному, тоже обнаруживает отсутствие рекомбинации, но имеет отцовское наследование. Оба типа полиморфизма дополняют друг друга, давая раздельную информацию об отцовском и материнском вкладе
вэволюцию популяций. Ядерные аутосомные локусы характеризуют общество в целом, не акцентируя внимание на особенностях генетического вклада различных полов [2].
Всвязи с этим, для выявления особенностей структуры генофонда двух этнографических групп татар и проведения филогенетического анализа с мировыми популяциям нам изучен полиморфизм 12 аутосомных STR-маркеров, 11 STR-локусов Y-хромосомы (7 локусов т.н. «минимального гаплотипа» и дополнительно маркеры DYS388, DYS426, DYS439 и DYS464a/b/c/d). Митотипическое разнообразие современного генофонда оценивали с помощью ПДРФ-анализа D-петли и высокоразрешающего рестрикционного анализа кодирующей части митохондриальной ДНК.
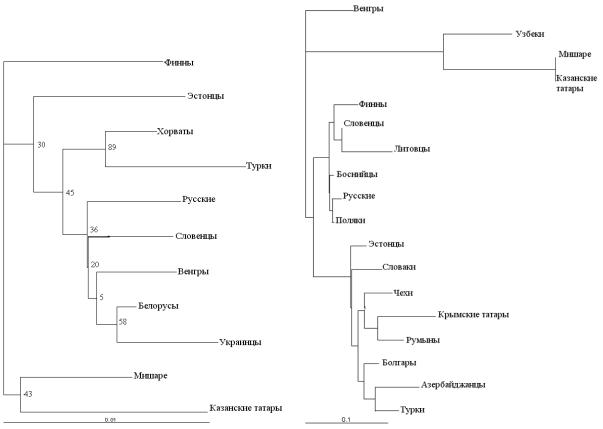
Необходимо отметить, что данные по генетическому полиморфизму аутосомных STR-локусов являются наиболее разрозненными. Так, полиморфизм STR маркеров широко изучен для популяций Европы, однако для населения РФ и стран СНГ данные носят фрагментарный характер. Поэтому, при сравнении этнографических группа поволжских татар с мировыми популяциями, в анализ вошли, преимущественно, представители славяноязычных и финно-угорских групп (рис. 1).
Топология полученной дендрограммы, с точки зрения лингвистики, представляется не совсем корректной, о чем, в принципе, свидетельствуют и довольно низкие значения достоверности для большинства из полученных кластеров (например, кластеризация популяций венгров с общим кластером белорусов и украинцев, или тюркоязычных турок со славяноязычными хорватами). Полученные результаты также трудно объяснить и с точки зрения географической близости взятых в анализ популяций. Однако, на полученной дендрограмме обе популяции поволжских татар образуют единый кластер, который занимает обособленное местоположение, что может указы-
234 |
ЗОЛОТООРДЫНСКОЕ НАСЛЕДИЕ. Выпуск 2. 2011 |
|
|
вать на их отличие от чисто европеоидных групп наличием в своем составе небольшой, но всетаки монголоидной примеси.
Рис. 1. Филогенетическое древо |
Рис. 2. Филогенетическое древо на основе |
на основе данных микросателлитных локусов |
данных полиморфизма мтДНК в европеоидных |
в европеоидных популяциях. Цифрами внутри |
популяциях. Для всех кластеров значения |
кластера обозначены значения бутстрепа, %. |
бутстрепа равны 100%. |
Также крайнее местоположение обе популяции поволжских татар занимают и при кластеризации с представителями монголоидов центрально-азиатского типа (монголы, буряты) и монголоидов с небольшой европеоидной примесью (алтайцы, хакасы), которые образуют один общий кластер, что лишний раз указывает на их европеоидную основу (здесь и далее дендрограммы не представлены).
Исходя из проведенного филогенетического анализа при сравнении поволжских татар с представителями народов Волго-Уральского региона, наблюдается объединение в один кластер двух групп мордвы: мокши и эрзи, и казанских татар и татар-мишар (здесь и далее графические результаты сравнительного анализа не представлены). Эти группы сближает общая европеоидная подоснова (в их составе по данным антропологии выделяют европеоидный компонент, как светлый европеоидный, так и темный европеоидный) и, вероятно этногенетические связи с эпохи Волжской Булгарии, Золотой Орды и Казанского ханства.
В отличие от аутосомных маркеров, полиморфизм митохондриальной ДНК является наиболее изученным. Существующая общепринятая классификация гаплогрупп мтДНК и определенный набор однонуклеотидных полиморфизмов, необходимый для отнесения того или иного типа мтДНК к определенной гаплогруппе, делает этот тип полиморфизма особенно удобным для проведения этногенетических исследований. Кроме того, благодаря материнскому типу наследования, можно довольно четко позволяет проследить основные этапы формирования того или иного этноса.
Исходя из данных, полученных в результате филогенетического анализа взаимоотношений изученных популяций татар с европеоидными представителями, можно отметить объединение этнографических групп татар с узбеками (рис. 2). Возможно, их объединяет не столько общая языковая принадлежность (тюркоязычные), сколько наличие в антропологическом составе общих
РАЗДЕЛ 2. Вопросы источниковедения и историографии истории Улуса Джучи |
235 |
|
|
компонентов: европеоидного темного и монголоидного южно-сибирского типа. По последнему, данные тюркоязычные группы и противопоставляются всем остальным чисто европеоидным группам.
При рассмотрении результатов филогенетического анализа популяций татар с представителями монголоидного типа, необходимо отметить наличие общего кластера между этими двумя этнографическими группами татар, который входит в одни общий кластер, образованный казахами и каракалпаками, с одной стороны, и ногайцами, с другой, что может говорить не только об их общей языковой принадлежности (тюркоязычной), но и общими этногенетическими связями, особенно, в составе Золотой Орды.
При сравнительном анализе полиморфизма митохондриальной ДНК популяций татар с народами Среднего Поволжья выявляется четкая кластеризация представителей тюркоязычной группы народов (татары и чуваши) и западные (в географическом плане) финноязычые группы, которые противопоставляются восточным, северо-восточным соседям: башкиры, удмурты и коми.
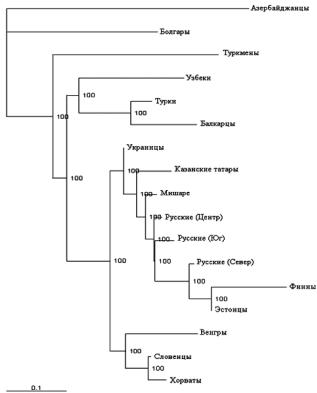
Филогения мужских линий показывает генетическое сходство поволжских групп татар с представителями западно-финской группы – финнов и эстонцев, и восточно-славянских популяций – русских и украинцев (рис. 3). Вероятно, их объединение можно объяснить наличием в их составе общего антропологического компонента и общими моментами этногенеза. Если включение древнефинского населения в состав русских и волжских булгар, а затем и татар хорошо согласуется с данными археологи, этнографии и антропологии, то включение в этот кластер украинских популяций может говорить об участии каких-то древнетюркских групп в формировании генофонда украинского народа в эпоху раннего средневековья и, особенно в эпоху Золотой Орды, когда территория современного проживания украинцев входила в состав Хазарского каганата и Улуса Джучи.
Рис. 3. Филогенетическое древо, основанное на распределении гаплогрупп Y-хромосомы, представителей европеоидных популяций. Цифрами обозначены значения бутстрепа, %.
При сравнительном анализе кластеризации монголоидных представителей с популяциями татар стоит отметить наличие общего кластера, образованного северными алтайцами, хантами и якутами и группами поволжских татар. Вероятно, данная ситуация отражает наиболее древние этногенетические связи. Попадание хантов (угроязычных) в общий кластер тюркоязычных представителей, на наш взгляд, объясняется участием угорских племен в формировании тюрков, при продвижении последних с востока на запад.
При сравнительном анализе этнографических группа татар с народами Среднего Поволжья по системе маркеров Y-хромосомы, выявляется некоторая обособленность татар в данном регио-
236 |
ЗОЛОТООРДЫНСКОЕ НАСЛЕДИЕ. Выпуск 2. 2011 |
|
|
не. Анализируя топологию полученного филогенетического древа, можно выявить наличие общего кластера, объединяющего татар Среднего Поволжья и чувашей. Вероятно, их объединяет не только общая языковая принадлежность (тюркоязычная), но и общие моменты этногенеза, связанные с эпохой великого переселения народов и Волжской Булгарии. Не исключено и наличие в этих группах общего сублапоноидного компонента, на что может указывать присоединение к данному кластеру кластер, объединивший наиболее ярких представителей сублапоноидного типа: марийцы и удмурты.
Таким образом, сравнительный анализ средневолжских татар с представителями монголоидного антропологического типа (монголы, якуты, буряты и т.д.) выявляет их относительную генетическую отдаленность, которую можно объяснить как территориальной удаленностью, так и разными моментами их этногенеза. В тоже время, при сравнении с европеоидными популяциями, выявляется небольшой, но ощутимый, монголоидный компонент (по данным Т.А. Трофимовой, выделяется два монголоидных типа: южно-сибирский и сублапаноидный). Его наличие в антропологическом составе поволжских татар можно объяснить несколькими причинами.
Во-первых: как проявление древнетюркских истоков (Центральная Азия, Алтай). Во-вторых: включением в свой состав местного финно-угорского населения с момента переселения раннеболгарских племен на Среднюю Волгу и ставшего одним из главных компонентов в формировании нового этноса – волжских булгар, а затем и татар. В-третьих: участием в формировании татар кыпчакских групп (казахи, киргизы) в период существования Золотой Орды и Казанского ханства.
По данным разнообразия основных филогенетических линий (женских и мужских, митохондриальной ДНК и Y-хромосомы), доля европеоидного компонента в генофонде современных татар составляет около 80%. Чем это можно объяснить? На всем протяжении формирования поволжских татар в их состав входили группы населения европеоидного облика, как на ранних этапах, так и по настоящее время. По данным археологии и антропологии известно, что при продвижении древнетюркских племен, в том числе и раннеболгарских, с Востока на Запад, они включали в свой состав позднесарматские (аланские) группы, которые по антропологическому типу, несомненно, являлись европеоидами. При переселении ранних болгар на Среднию Волгу были ассимилированы местные финно-угорские племена (о чем свидетельствуют данные, полученные по полиморфизму мтДНК некоторых захоронений древнего народонаселения Среднего Поволжья), которые также являются, в основном, носителями европеоидного антропологического типа.
Однако, как было отмечено нами ранее, антропологический облик татар не являлся постоянным, тоже самое касается и генофонда популяций, который в значительной степени подвержен дрейфу генов, но также не последнюю роль играют процессы миграции и метисации. На это указывают данные, полученные в ходе анализа полиморфизма митохондриальной ДНК: если для древних популяций Среднего Поволжья характерно преобладание прародительских финноугорских черт, то современные популяции татар выявлют сходство с тюркоязычными. Однако, генофонд популяции является более-менее стабильной организацией, на что указывает наличие филогенетических связей по мужской линии современных популяций татар с типичными представителями финно-угорской группы.
Еще одним важным выводом проведенного исследования является определение генетической близости двух этнографических групп поволжских татар: казанских татар и татар-мишар. Несмотря на некоторые различия в языке и культурных традиций, формирование их этногенетического облика шло в общем русле. Детализация этого процесса задача будущих исследований.
1.Кравцова О.А. Молекулярно-генетический анализ древних и современных образцов ДНК. Дис. ...
канд. биол. наук. – Казань, 2006. – 160 с.
2.Лимборская С.А., Хуснутдинова Э.К., Балановская Е.В. Этногеномика и геногеография народов Восточной Европы. – М.: Наука, 2002. – 261 с.
ABSTRACT
Olga Kravtsova, Ilgizar Gazimzyanov, Ilnur Mirgaleev
Genetic portrait of Middle-Volga River region Tatars
The aim of this research is an evaluation of genetic originality of two ethnographic groups of Volga river Tatars: Kazan Tatars and Tatar-Mishares resident on territory of the Republic of Tatarstan. On the base of data on 12
РАЗДЕЛ 2. Вопросы источниковедения и историографии истории Улуса Джучи |
237 |
|
|
autosomal microsarellite STR loci, 11 STR markers of Y-chromosome and RFLP-analysis of mitochondrial DNA comparative analysis with the European and Mongolian populations, and also with population of Middle-Volga river region was conducted.
Our study showed the prevalence of European component in the genofond of modern Tatars (about 80%) both in paternal and maternal lineages (Y-chromosome haplotypes and mitochondrial halpogroups gene pool). Our data are also in accordance with anthropological data. Besides, the results of phylogenetic analysis specify on close genetic relationships of two groups of Volga Tatars. Probably, their formation, in spite of some language and cultural differences, flowed under the same conditions.
Ж.М. Сабитов, О.А. Балаганская, М.А. Кузнецова, И.М. Тажигулова, О.П. Балановский, Д.М. Дубербаев
Возможности популяционной генетики
вреконструкции этногенеза тюркских народов
В60-е годы 20 века лауреат двух Нобелевских премий Лайнус Полинг высказал идею, что скорость накопления мутаций в ДНК является постоянной, и ее можно использовать как своего рода молекулярные часы эволюционной истории, в том числе и для изучения событий эволюционной истории человека. Таким образом, сравнив два ДНК можно определить генетическое время расхождения этих двух ДНК. Данный метод стал использоваться в биологии для нахождения времени расхождения различных близких биологических видов. Тогда ученные начали изучать с помощью этого метода человеческие популяции.
Хромосомный набор человека состоит из 23 пар хромосом, из которых 22 пары – «обычные» хромосомы, или аутосомы, и одна пара – половые, определяющие принадлежность к женскому или мужскому полу. У женщин они представлены парой одинаковых Х-хромосом, а у мужчин – одной Х- и одной Y-хромосомой, от наличия или отсутствия которой зависит, будет ребенок мальчиком или девочкой. При зачатии будущий ребенок наследует половину своих хромосом от отца, а половину – от матери; что касается половых хромосом, то от матери могут наследоваться только Х, а от отца – либо Х, либо Y. Таким образом и происходит определение пола ребенка [9, с. 49].
В1987 году вышла одна из первых статей по применению в популяционных исследованиях сиквенсов митохондриальной ДНК. Для анализа Аллан Уилсон, профессор Калифорнийского университета в Беркли, выбрал не ядерную ДНК, а ДНК митохондрий – одного из органоидов клетки, так как митохондриальная ДНК (мтДНК) – небольшая кольцевая молекула в женской Х- хромосоме размером 16 600 пар нуклеотидов, содержащая всего 37 генов.
Аллан К. Уилсон, на основе анализа митохондриальной ДНК утверждал, что все человечество произошло от одной женщины, когда-то жившей в Африке, потомки которой заселили остальные континенты, породив все расовое разнообразие человечества. Подробные результаты этих исследований были опубликованы в 1992 году в авторитетном журнале «Science». Позже различные группы популяционных генетиков начали изучать Y-хромосомы современных людей, собрав материал для анализа почти во всех регионах мира. Результат изучения нескольких тысяч проб, взятых от представителей разных народностей, показал, что все мужчины на Земле восходят к единому предку, названному «генетическим Адамом». По оценке исследователей, «генетический Адам» жил 150– 160 тысяч лет в Африке и потом от него выделились различные генеалогические линии.
Наиболее интересными для историков представляются исследования по Y-хромосоме. Здесь можно процитировать известного популяционного генетика с мировым именем Льва
Анатольевича Животовского:
«Y-хромосома – самая маленькая и самая неинформативная в человеческом геноме, она содержит всего около 80 генов. Другие хромосомы намного больше – в среднем каждая из них несет около 1.5 тыс. генов. Y-хромосома не только мала, но еще и узко специализирована – почти все ее гены ответственны за чисто мужские признаки. Долгое время из-за скромных размеров и небольшого числа генов Y-хромосома считалась «генетической пустышкой», малоинтересной для ученых, разве что только в связи с проблемами мужского бесплодия. Однако к концу прошлого века исследователи заинтересовались этой хромосомой с неожиданной стороны. Y-хромосома передается по отцовской линии, т.е. исключительно от отца к сыну. Поэтому, маркируя эту хромосому, т.е. пометив ее каким-либо образом, теоретически можно отследить ее происхождение по муж-
238 |
ЗОЛОТООРДЫНСКОЕ НАСЛЕДИЕ. Выпуск 2. 2011 |
|
|
ской линии до далеких предков. Маркерами Y-хромосомы служат ее мутации, возникающие в разное время и в разных географических регионах. Другими словами, каждый мужчина несет в своей Y-хромосоме генетическую память в виде всех тех мутаций, которые возникали у всех его прямых предков по мужской линии: его отца, деда, прадеда. Вторая важная особенность Y- хромосомы – непарность: она единственная хромосома человеческого генома, не имеющая пары. В обычных, парных хромосомах, в каждом поколении гены перемешиваются благодаря тому, что хромосомы обмениваются своими частями (рекомбинируют). Если в такой хромосоме возникает мутация, то в результате рекомбинации она может перейти в пару этой хромосомы, т.е. потеряться. Но бедной одинокой Y-хромосоме не с кем обмениваться генами, поэтому в ней любые изменения сохраняются в течение неограниченного времени» [10, с. 49].
В2002 году консорциум по Y-хромосоме предложил номенклатуру линий Y-хромосомы, основанную на последовательности происхождения маркеров. Выделено 18 основных гаплогрупп, обозначаемых буквами латинского алфавита от А до R. Позже зачастую вносились незначительные поправки в эту классификацию. Современные методы генетического анализа позволяют установить порядок возникновения мутаций и нарисовать филогенетическое дерево, описывающее родственные связи всех людей по мужской линии. Чем раньше в эволюции возникла мутация, тем ближе к "корню дерева" она располагается, и тем больше ветвей будут ее содержать. При известной скорости накопления мутаций в расчете на поколение, их число можно перевести в абсолютное время, прошедшее с момента появления генетических линий, т.е. установить время, когда жил общий предок этих линий [7, c. 7].
Мутации в Y-хромосоме бывают двух видов: SNP-маркеры (Single-nucleotide polymorphism – однонуклеотидные полиморфизмы) и STR-маркеры (short tandem repeats – короткие тандемные повторы).
Мутации в SNP-маркерах относятся к мутациям, которые позже присутствуют у всех мужских потомков данного субъекта. Их используют, чтобы показать по какой генетической линии человек восходит к генетическому Адаму. С помощью SNP-маркеров можно определить гаплогруппу, которую, проводя аналогии, можно назвать «генетической фамилией». Количество таких «генетических фамилий» довольно мало, всего около 20 общих гаплогрупп (обозначены буквами латинского алфавита) и около 400 разных вариантов, входящих в эти 20 основных гаплогрупп.
Вто время как STR-маркеры можно использовать для поиска близкого родства. STR-мар- керы представляют собой совокупность определенных частей ДНК (DYS – DNA Y Segment), в которых были замерены тандемные повторы определенных нуклеотидов.
Например, можно привести примеры различных STR-маркеров:
Квадруплетный маркер ТАGA (тимин-аденин-гуанин-аденин) носит название DYS#19. Триплетный маркер ATT (аденин-тимин-тимин) носит название DYS#388 Квадруплетный маркер TCTA (тимин-цитозин-тимин-аденин) носит название DYS#391 Триплетный маркер ТAT (тимин-аденин- тимин) носит название DYS#392. Квадруплетный маркер AGAT (аденин-гуанин-аденин-тимин) носит название DYS#393.
Если в DYS 19 всего 16 повторов ТАGA, то, следовательно, значение маркера будет 16. Последовательность таких цифр (значений повторов) называется гаплотип. Например, у одного из авторов статьи следующий гаплотип (значения повторов):
13, 31, 25, 15, 17, 11, 14, 15, 14, 11, 20, 13, 10, 11, 13, 10, 23, 12, 15.
Формат записи (номера DYS):
389-I, 389-II, 390, 456, 19, 385а, 385b, 458, 437, 438, 448, GATA H4, 391, 392, 393, 439, 635, 388, 426.
Вобщем, говоря простым языком, у ближайших родственников по мужской линии гаплотип должен совпадать. Обычно гаплотип передается неизменным от отца к сыну, но иногда раз в n-ое количество поколений случаются мутации и, например, в DYS 19 вместо 16 повторов, как у отца, появляются либо 15, либо 17 повторов (мутация произошла).
Тут стоит отметить, что мутация носит случайный характер, хоть и подчиняется определенным статистическим законам. Например, у отца и сына различие один шаг мутации в одном маркере, мы на основе только ДНК можем предположить, что они родственники, но скорее всего не близкие. Хотя знание документальной генеалогии поможет нам заметить, что они являются близкими родственниками, просто случайная мутация «отдалила» их родство. Также, например, у двух потомков одного человека могут быть разные гаплотипы, у одного в отличие от предка слу-
РАЗДЕЛ 2. Вопросы источниковедения и историографии истории Улуса Джучи |
239 |
|
|
чилось две мутации, в то время как у другого ни одной. Поэтому чтобы избежать таких казусов, всегда следует исследовать не малое количество участников, а увеличивать свою выборку количественно. В этом случае статистические погрешности уже не будут играть такую большую роль.
Также стоит отметить такое явление как Гомоплазия. Бывают редкие случаи, когда гаплотипы разных гаплогрупп имеют одинаковые значения повторов. Но, в то же время, они принадлежат разным гаплогруппам. Тут стоит отметить, что мутации в этих гаплогруппах привели конвергентным образом к тому, что значения в STR-маркерах стали похожи, хотя генетическое родство между ними может измеряться десятками тысяч лет (например, некоторые гаплотипы гаплогрупп R1a и С3, которые генетически разошлись более 80 тысяч лет назад). Идентичные значения в гаплотипе должны быть обязательно проверены первым видом тестирования: SNP-маркерами. Вероятность гомоплазии изначально статистически мала, но пренебрегать ею не стоит. Поэтому тестируя и изучая Y-хромосому, стоит проводить как тестирование по SNP-маркерам, так и тестирование по STR-маркерам. Причем стоит отметить, что чем больше будет количество тестируемых STR-маркеров (в вышеуказанном гаплотипе их было 19), тем точнее будут результаты сравнения с другими гаплотипами из базы данных.
Нелишним будет также отметить, что во многом успешность сравнения зависит от базы данных, с которой можно сравнивать полученные гаплотипы. Например, в Европейских странах базы данных большие и сравнивать всегда есть с кем, в то же время данных по тюркским народам бывшего улуса Джучи данных очень мало и большинство из них имеют малое количество тестированных STR-маркеров.
Здесь упомянуть диссертации различных популяционных генетиков, которые изучали ДНКполиморфизм у разных тюркских народов Евразии: Кравцова О.А. [12], Боготова З.И. [9], Юнусбаев Б.Б. [19], Лобов А.С. [14], Харьков В.Н. [18]. Кутуев И.А. [13].
Существует большое количество публикаций о гаплотипах и гаплогруппах народов-наслед- ников Золотой Орды. Здесь стоит перечислить наиболее значимые работы:
Встатье Яницы Дж. и соавторов 2005 г. были исследованы гаплотипы 125 польско-литов-
ских татар по 9 маркерам (DYS19, DYS385a, DYS385b, DYS389-1, DYS389-2, DYS390, DYS391, DYS392, DYS393) [3, c. 91].
Вдиссертации Кравцовой О.А. были исследованы гаплотипы 36 казанских татар Азнакаевского района и 64 гаплотипа татар-мишар Буинского района по 12 маркерам (DYS19, DYS385a, DYS385b, DYS389-1, DYS391, DYS392, DYS393, DYS448, DYS464a, DYS464b, DYS464c, DYS464d) [11, c. 156–160]. Также интересным является выделение ДНК из различных захоронений на территории Татарстана (всего 49 образцов), но, к сожалению, данные образцы тестировались по аутосомным маркерам [12, c. 50–53], что в плане истории менее информативно, чем тестирование по Y-хромосоме. К этому стоит добавить, что тестирование древних образцов по Y- хромосоме технологически сложнее, чем анализ древней митохондриальной ДНК, поскольку затруднено тем, что ядерная ДНК разрушается быстрее, чем другие части ДНК.
В1999 году в рамках исследования четырех популяций: уйгуров, высокогорных киргизов, киргизов из низкогорий и казахов были исследованы гаплотипы 172 человек (49 казахов, 39 уйгур, 41 киргизов из равнин, 43 высокогорных киргизов) по семи маркерам (DYS19, DYS388, DYS389-1, DYS390, DYS391, DYS392, DYS393) [4, c. 208].
Довольно интересными оказались опубликованные на специализированном сайте http://www.yhrd.org данные Томаса Ротамеля по казахам из города Тараз, где было получено 181
гаплотипов по 12 маркерам (DYS19, DYS385a, DYS385b, DYS388, DYS389-1, DYS389-2, DYS390, DYS391, DYS392, DYS393, DYS426, DYS439).
Также очень интересными представляются данные собранные венгерскими генетиками о гаплотипах 45 казахских маджар, проживающих в Тургае [1, c. 305]. Гаплотипы исследовались по
12 маркерам (DYS19, DYS385a, DYS385b, DYS389-1, DYS389-2, DYS390, DYS391, DYS392, DYS393, DYS437, DYS438, DYS439). Данное исследование примечательно тем, что в его основу была положена историческая гипотеза о родстве венгров-мадьяр и казахских маджар («являются ли мадьяры и тургайские маджары родственниками»), до этого в основу всех статей лежали биологические вопросы («какие гаплотипы у определенного народа»). В итоге гипотеза не подтвердилась, так как гаплотипы венгров и гаплотипы маджар не совпали. При этом маджары по шежире (генеалогические записи, передаваемые от отца к сыну, содержащие как данные по происхождению рода так и некоторые родовые предания) относятся к племени Аргын, и их гаплотипы ока-
240 |
ЗОЛОТООРДЫНСКОЕ НАСЛЕДИЕ. Выпуск 2. 2011 |
|
|
зались близки представителям этого племени, что подтвердило версию о том, что тургайские маджары часть Аргынов.
Среди исследований также есть исследования о гаплогруппах татар, проживающих в Китае и переселившихся туда в 19 веке. Всего были исследованы 33 образца китайских татар [6, c. 314]. Недостатком этой работы было то, что тестирование проводилось всего по 8 STR-маркерам
(DYS19, DYS388, DYS389-1, DYS389-2, DYS390, DYS391, DYS392, DYS393).
Также весьма интересным является исследование группы ученных по каракалпакам. Каракалпаки состоят из 2 больших племенных групп:
1.Конгират (в состав конгиратов кроме самих конгиратских родов входят кияты, уйгуры, муйтены и др.).
2.Он торт уру (14 «родов») (в эту группу входят кыпчак, китай, кенегес, мангыт).
В2007 году были опубликованы первые 6-маркерные гаплотипы каракалпаков (исследованы следующие маркеры: DYS388, DYS389-1, DYS390, DYS391, DYS392, DYS393) [2, c. 43–48]. Ос-
новными недостатками того исследования были короткие 6-маркерные гаплотипы, которые обладают малой информативностью для человека, изучающего родоплеменную структуру каракалпаков. Также в статье была озвучена субэтническая принадлежность каракалпакских образцов: 54 каракалпаков-конгиратов и 54 каракалпаков-он торт уру. Но, к сожалению, не были озвучены данные по более низким родовым делениям. По косвенным данным из таблиц, мы можем узнать, что среди племенной группы Он торт уру большинство тестированных каракалпаков относились
кследующим родам: Клан Омир и клан Нукис племени Кенегес, Клан Карасирак племени Мангыт. К сожалению, из малого количества маркеров довольно трудно будет получить какую-то новую информацию по гаплотипам, ведь на 6 маркерах эффект гомоплазии имеет большую вероятность, а расчет времени жизни предка популяции по 6 маркерам может иметь очень большую погрешность.
Также интересными являются исследования по хазарейцам:
Хазарейцы как народ согласно Рашид ад-Дину возникают из джучидских отрядов Бату и Орды, находившихся в армии Хулагу. В 1262 г. из-за конфликта между Берке-ханом и Хулагу Джучиды в хулагидской армии оказались под угрозой уничтожения. Часть из них прорвалась на север
кБерке, часть ушла к мамлюкам на запад, а часть, спасаясь от преследований, ушла на восток – в Хорасан. Здесь они заняли области от Газны до Мултана и Лахора. Имя старшего амира джучидов было Никудер [17, c. 81–82]. Основываясь на том, что именно потомки Орда-эджена именовали себя царями Газны и Бамиана (хотя фактически не контролировали данные территории), можно предположить, что хазарейцы и их предки некудерийцы были потомками тумена Кули, сына Ор- да-эджена. Как мы знаем в походе Хулагу в Иран участвовали три тумена от джучидов: Тутар, сын Мингкадара, сын Бувала, сына Джучи во главе тумена от Западного крыла улуса Джучи (остатки Тумена во главе с Ногаем ушли на север в Золотую Орду), Балакан, сын Шибана, сына Джучи, во главе тумена от центра улуса Джучи и Кули, сын Орда-эджена, от восточного крыла улуса Джучи. В тумен Кули входили помимо войск улуса Орда-эджена, войска улусов ТукаТимура, Шингкума, Сингкума и Удура, которые входили в Восточное крыло улуса Джучи. Говоря простым языком, хазарейцы являются мини-копией населения восточного крыла улуса Джучи 13 в. Судя по публикациям популяционных генетиков: половина хазарейцев имеет «монгольское происхождение» (гаплогруппы С3, О3), а другая половина «дештикыпчакское происхождение» (R1b, Q) [5, c. 202]. Как известно гаплогруппа R1b в огромном количестве встречается среди башкир [14, c. 76], а также по предварительным данным среди казахстанских кыпчаков. Таким образом, анализируя данную популяцию, мы вполне можем ответить на поставленный Костюковым В.П. вопрос «каково было соотношение кыпчаков и монгол в улусе Джучи» [11, c. 197], хотя естественно это будет частью ответа и будет касаться в основном восточной части улуса Джучи. При этом не следует забывать, что дрейф генов (включая эффект основателя и бутылочного горлышка) мог значительно изменить это соотношение.
Вданном обзоре мы слабо затронули исследования по гаплогруппам, так как без гаплотипов не обойтись в довольно узкой области: реконструкции сравнительно молодых (в пределах 1000 лет) родовых групп.
К сожалению почти не проводились исследования популяционных генетиков по STRмаркерам среди таких народов как крымские, астраханские, сибирские татары, башкиры, ногайцы, кумыки.