Білковий обмін
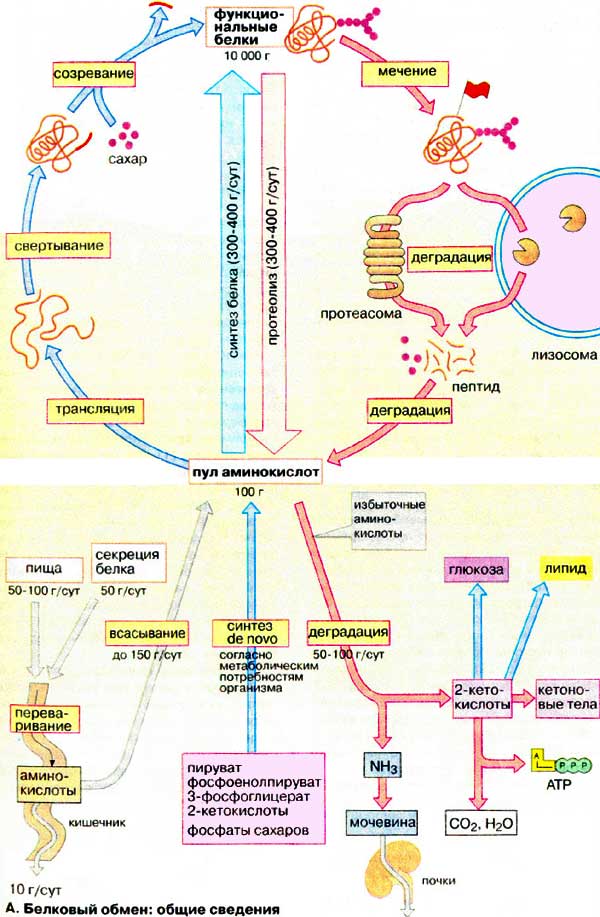
Білковий обмін: загальні відомості
Одержані з їжею білки підлягають повному гідролізу в шлунково-кишковому тракті до амінокислот, які всмоктуються та з кровотоком розподіляються в організмі. 8 з 20 білкових амінокислот не здатні синтезуватися в організмі людини – це незамінні амінокислоти, що повинні надходити з їжею.
Через кишечник і також через нирки в невеликих кількостях організм постійно втрачає білок, тому щодобово необхідно отримувати з їжею не менше 30 г білка.
Фізіологічний мінімум ‑ кількість білка в раціоні, необхідна для підтримування балансу азоту в організмі. В індустріальних країнах вміст білка в їжі найчастіше значно перевищує норму. Амінокислоти не запасаються в організмі. При надлишковому надходженні амінокислот в печінці окислюється або використовується до 100г амінокислот за добу. Азот, що міститься в них, перетворюється на сечовину і в такій формі виділяється з сечею, а вуглецевий скелет використовується в синтезі вуглеводів, ліпідів або окислюється з утворенням АТФ.
Вважають, що в організмі дорослої людини щодобово розщеплюється до амінокислот 300-400г білка (протеоліз). Одночасно приблизно така ж сама кількість амінокислот включається до утворених молекул білків (білковий біосинтез).. Високий обіг білка в організмі необхідний тому, що багато білків відносно недовговічні: вони починають оновлюватися через декілька годин після синтезу, а біохімічний напівперіод оновлення складає 2-8 днів. Навпаки довговічними є структурні білки, гістони, гемоглобін або компоненти цитоскелету.
Ключеві ферменти проміжного обміну оновлюються через декілька годин після синтезу. Поєднання процесів розщеплення та ресинтезу дозволяє клітинам швидко регулювати рівень і активність найбільш важливих ферментів відповідно з метаболічними потребами. Майже всі клітини здатні здійснювати біосинтез білків (на схемі наверху зліва). Побудова пептидного ланцюга шляхом трансляції проходить у рибосомах. Однак, активні форми більшості білків виникають тільки після ряду подальших кроків. По-перше за допомогою допоміжних білків шаперонів повинна утворитися біологічна активна конформація пептидного ланцюга (згортання). При посттрансляційному дозріванні у багатьох білках видаляються частини пептидного ланцюга або приєднуються додаткові групи, наприклад олігосахариди або ліпіди. Такі процеси відбуваються в ендоплазматичному ретикулумі та в апараті Гольджи. Потім, білки повинні транспортуватися у відповідну тканину або орган (сортирування).
Внутришньоклітинне розщеплення білків (протеоліз) відбувається частково в ліпосомах. Крім того, в цитоплазмі наявні органели ‑ протеасоми, в яких підлягають руйнуванню неправильно згорнуті або денатуровані білки. Такі молекули ідентифікують за допомогою спеціальних маркерів.
Етапи розщеплення білків:
І. Шлунок (початок розщеплення білків): утворюється суміш пептидів і молекул білків, що не розщепилися.
Ферменти:
пепсин (протеолітичний фермент шлункового соку), утворюється в головних клітинах залоз слизової оболонки шлунка в неактивній формі у вигляді проферменту – пепсиногену, який перетворюється на пепсин під дією шлункового соку, а також під впливом пепсину (аутокаталітичний процес активації).Розщеплює внутрішні пептидні зв’язки білка, продукти гідролізу – пептони, або пептиди;
хімозин (у немовлят і молодих тварин, що харчуються тільки материнським молоком), специфічно розщеплює основний білок молока – казеїноген, перетворюючи його на казеїн.
Роль НСІ:
забезпечує необхідну кислотність для каталітичної дії ферментів (рН = 1,5 – 2,5);
каталізує процеси перетворення пепсиногену на пепсин;
каталізує процеси набухання і розпушування білків (полегшує каталітичну дію ферментів;
запобігає розвитку в шлунку процесів гниття та бродіння (шлунковий сік містить 0,4-0,5% вільної соляної кислоти).
ІІ. Тонка кишка (розщеплення білків і пептидів до амінокислот)
Перетравлювання білків і пептидів відбувається:
у порожнині кишок (піддаються гідролітичному розщепленню в основному молекули білка);
на поверхні слизової оболонки кишок (пристінне перетравлювання) – гідролітичне розщеплення молекул пептидів з різною кількістю залишків амінокислот.
Ферменти:
трипсин (фермент соку підшлункової залози), виділяється в неактивній формі у вигляді трипсиногену. Фермент кишкового соку ентерокіназа перетворює трипсиноген у трипсин, іони кальцію (активація процесу). Оптимальна активність при рН = 7 – 8, діє на білки, що не розщепилися, і відносно високомолекулярні пептиди.
хімотрипсин виділяється соком підшлункової залози в неактивні йформі у вигляді хімотрипсиногену (циклічний полі пептид), гідролізує пептидні зв’язки, утворені карбоксильнми групами амінокислот фенілаланіну, тирозину, триптофану, метіоніну та лейцину. Оптимальна активність при рН = 7 – 8.
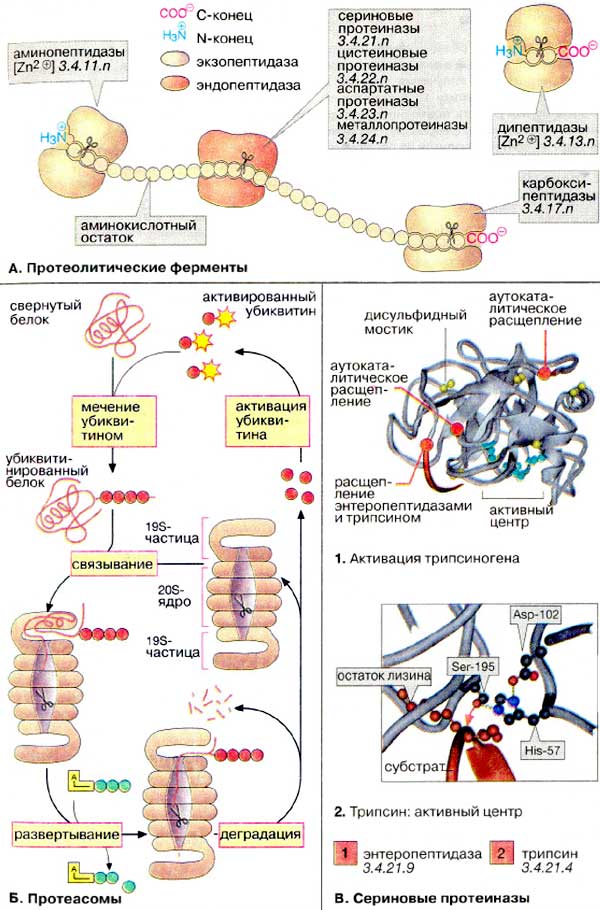
екзопептидази (амінопептидази, карбоксипептидази, дипептидази), належать до металоферментів, тобто активуються іонами Мg2+, Mn2+, Co2+, Zn2+ (забезпечують утворення фермент-субстратного комплексу).
|
Карбоксипептидази каталізують процес відщеплення амінокислотних залишків з С-кінця пептичного ланцюга, активуються трипсином. Амінопептидази каталізують відщеплення N-кінцевих залишків амінокислот за умови вільної NH2-групи. Дипептидази розщеплюють дипептиди до амінокислот. |
|
|
|
||
Карбоксипептидаза |
Активація трипсиногену |
Активний центр трипсину |
||
|
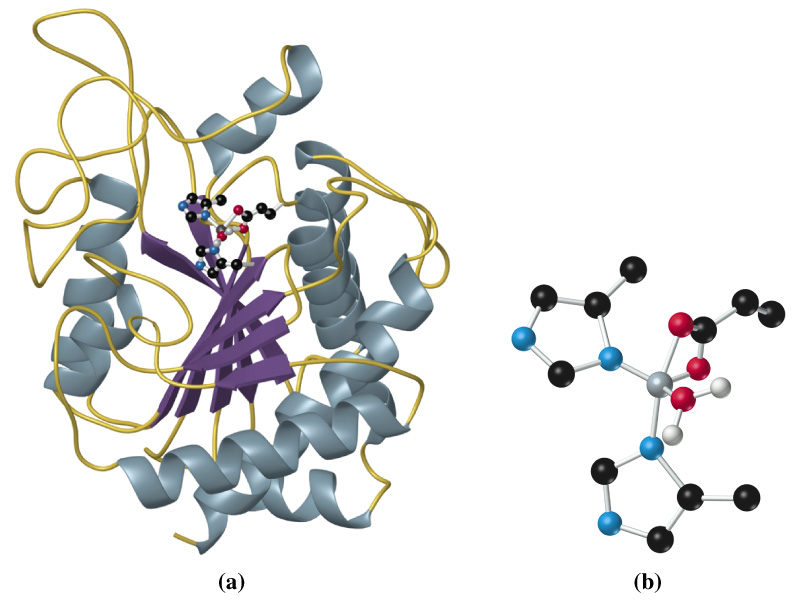
Велика група протеіназ містить в активному центрі серин: ферменти травлення трипсин, хімотрипсин і еластаза, фактори згортання крові, фермент плазмін і його активатори. Панкреатичні протеінази виділяються у вигляді проферментів (зимогенів). Активація таких ферментів основана на протеолітичному розщепленні. Процес активації показаний на прикладі трипсиногену, попередника трипсину. Процес активації починається з відщеплення N-кінцевого гексапептиду ентеропептидазою («ентерокіназою»), специфічною сериновою протеіназою, яка локалізована на мембранах кишкового епітелію. Продукт розщеплення (β-трипсин) ферментативно активний і розщеплює наступну молекулу трипсиногену в місцях, позначеним червоним кольором (аутокаталітична активація). Залишок серину за участі залишків гістидину та аспартату атакує зв’язок (червона стрілка). Частина пептидного субстрату розміщена в С-кінцевій стороні від залишку лізину, боковий ланцюг якого під час каталізу фіксується в спеціальному «кармані» ферменту. Протеасоми Оскільки функціональні білки клітини повинні бути захищені від передчасного протеолізу, частина протеолітичних ферментів клітини вміщена в ліпосоми. Інша система деградації білків локалізована в цитоплазмі. Вона складається з великих білкових комплексів, протеасом. Протеасоми містять бочковидне ядро з 28 субодиниць. Протеолітична активність (на схемі показана у вигляді ножиць) локалізована у внутрішньому 20S-ядре. С торців бочки запираються складно побудованими 19S-часточками, як контролюють доступ в ядро. Білки, які будуть розщеплюватися в протеасомі (наприклад, ті, що містять помилки транскрипції або старі молекули), маркуються шляхом ковалентного зв’язування білком убіквітіном. Убіквітин активований завдяки наявності тіолскладного зв’язку. Помічені убіквітином молекули розпізнаються 19S-часточками з використанням АТФ і потрапляють в ядро, де відбувається їх деградація. Убіквітин не руйнується та після активації використовується знову. |
|
||