

2 курс / Нормальная физиология / Общий_курс_физиологии_человека_и_животных_Том_2_Ноздрачев_А_Д_,
.pdf8
Функции кровообращения
Непрерывное движение крови в организме обеспечивается системой органов кровообращения — сердцем и сосудами. Посредством кровообращения достигается интеграция различных функций организма и его участие в реакциях на изменение окружающей среды.
8.1 Элементы эволюции
Совершенствование организации сердечно-сосудистой системы представляет собой результат длительного филогенетического развития. Система транспорта у простейших — диффузия — дополняется интенсивным движением протоплазмы. Это свойство сохранилось почти у всех клеток многоклеточных. Губки и кишечнополостные используют для транспорта веществ среду своего обитания. Жгутиками или сокращением мышц они проталкивают воду по межклеточным каналам. У нематод, иглокожих, некоторых полихет и голотурий движение жидкости поддерживается соматическими мышцами.
У членистоногих, моллюсков, низших хордовых появляется незамкнутая ииркуляторная система, в которой сердце перекачивает гемолимфу. По системе трубок она достигает различных частей тела, где сосуды прерываются специальными лакунами, у которых отсутствуют собственные стенки. Гемолимфа смешивается с тканевой жидкостью, проникает в межклеточное пространство и затем вновь возвращается в сердце. Это пока еще незамкнутая система.
Характерной чертой эволюции является постепенное обособление аппарата кровообращения и превращение его в замкнутую систему (рис. 8.1). У олигохет, многих полихет, головоногих моллюсков появляется замкнутая система кровообращения с одним или несколькими нагнетающими устройствами. На этом этапе эволюции кровь соприкасается с тканями при помощи капилляров. У некоторых животных образуются специальные синусы.
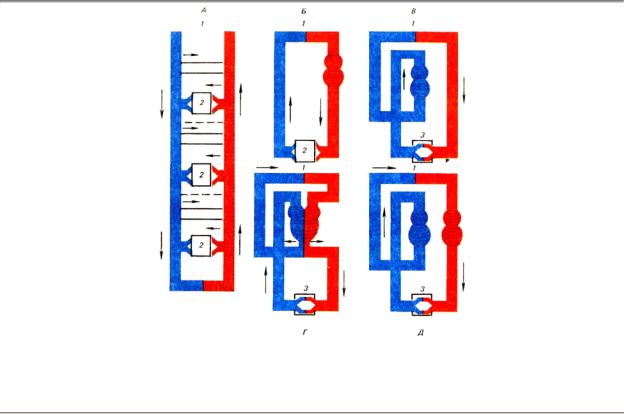
Рис. 8.1. Эволюция системы кровообращения. А — черви; Б — моллюски; В — рыбы; Г — амфибии; Д
— высшие позвоночные: Сократительное образование, сообщающее движение крови, заштриховано; артериальная система обозначена красным цветом, венозная — синим; 1 — местоположение органа дыхания (на рисунке не обозначен), 2 — лакунарная часть системы, 3 — капилляры
Таким образом, возникает два вида внутренней среды: кровь, циркулирующая по сосудам, и тканевая жидкость, или лимфа, находящаяся в межклеточных пространствах.
Одновременно с усложнением периферической части кровеносной системы происходит постепенное структурное и функциональное обособление специализированного центрального органа — сердца. У некоторых низших хордовых животных, таких, как ланцетник, роль насоса выполняет пульсирующий брюшной сосуд, у червей — спинной.
Упиявок существует два боковых пульсирующих сосуда, сокращающихся поочередно.
Учленистоногих, моллюсков произошло обособление сердца, что значительно увеличило эффективность системы кровообращения. Сердце большинства членистоногих представляет собой сокращающуюся трубку, у некоторых видов оно свободно лежит в перикардиальной полости. У многих насекомых сердце «подвешено» при помощи мышц. Сокращаясь, мышцы растягивают сердце, создавая в нем отрицательное давление.
Следующим этапом эволюции явилось возникновение камерных сердец, а также выделение большого и малого (легочного) кругов кровообращения. У рыб существует только один круг, сердце разделено на две камеры. Из желудочков вся кровь направляется в жабры; обогащенная кислородом, она разносится, затем по всему телу и после этого возвращается в предсердия. Сердца моллюсков уже состоят из одного-двух предсердий и желудочка. У головоногих моллюсков вблизи, основания жабр находятся еще дополнительные, проталкивающие венозную кровь сердца. Такая система оказывается эффективнее системы кровообращения рыб, в которой сердцу приходится преодолевать двойное сопротивление: жаберных сосудов и сосудов остальных частей тела. У рыб происходит выделение лимфатической системы в самостоятельную. Она представлена у них лимфатическими мешками и сложноразветвленными трубками. Периферические концы трубок замкнуты, центральные — открываются в вены.
Выход из водной среды на сушу, повышенная гравитация предъявили новые требования к системе кровообращения: возросла масса сердца, изменились сосудистые стенки. Низкое давление в малом (легочном) круге кровообращения и соответственно более высокое давление в большом круге позволяют рептилиям и гомойотермным животным выдерживать дополнительную нагрузку, связанную с переходом к наземному

образу жизни. У амфибий разделение двух кругов кровообрашениаяедшнаое. У них нет межжелудочковой перегородки, хотя имеются межжелудочковые складки. Венозная кровь из органов поступает по венам в правое предсердие, а оттуда в желудочек. Желудочек нагнетает кровь в малый круг кровообращения. Прошедшая через легкие и обогащенная кислородом аэрированная кровь возвращается в левое предсердие и затем поступает в тот же общий желудочек. Считают, что из-за наличия межжелудочковых складок в сердце не происходит полного смешивания крови. Аэрированная кровь направляется сердцем в аорту и далее в сосуды головы. В остальные органы поступает смешанная венозная и аэрированная кровь.
Сердце рептилий характеризуется наличием двух желудочков, разделенных перегородкой. У ящериц и змей она неполная, но потоки аэрированной и неаэрированной крови в какой-то степени разделены. У высших рептилий (крокодил), птиц и млекопитающих межжелудочковая перегородка сформирована полностью. В связи с разделением кровообращения на большой и малый круги в правой половине сердца циркулирует только венозная кровь, в левой — артериальная (рис. 8.2). В сердце плода млекопитающих между предсердиями имеется овальное отверстие, которое зарастает после рождения.
Рис. 8.2. Кровообращение млекопитающих (А) и распределение крови в сосудах различного типа (Б): 1
— капилляры головы, 2 — легочная вена, 3 — дуга аорты, 4 — левое предсердие, 5 — левый желудочек, 6 — брюшная аорта, 7 — капилляры большою круга, 8 — кишечная артерия, 9 — воротная вена, 10 — печеночная вена, 11 — правый желудочек, 12 — правое предсердие, 13 — легочная артерия, 14 — капилляры, 15 — русло емкостных сосудов, 16 — русло резистивных сосудов, 17 — ткани, 18 — русло обменных сосудов
Совершенствование функций сердца и сосудов в процессе филогенеза происходило одновременно с совершенствованием нервных связей системы кровообращения. Сердце низших позвоночных не имеет нервных связей с другими системами. Иннервация сердца появляется только у миног, миксин. Она осуществляется блуждающим нервом, симпатической иннервации еще нет. Эти особенности определяются низкими адаптационными возможностями системы кровообращения животных, обитающих в относительно более постоянных условиях среды, чем наземные.
8.2. Функции сердца
Источником энергии, необходимой для продвижения крови по сосудам, является работа сердца. Его строение изменялось и совершенствовалось в процессе филогенеза. Различают следующие типы сердец: пульсирующие сосуды, трубкообразные сердца, камерные сердца, ампулярные добавочные сердца.
Пульсирующие сосуды наиболее распространены на низших стадиях эволюции и особенно отчетливо выражены у кольчатых червей. Они могут сокращаться перистальтически или локально. Жидкость циркулирует в них медленно. Пульсирующие сосуды у насекомых трудно отличить от трубкообразных сердец.
Типичные трубкообразные сердца имеются у большинства членистоногих. В этих сердцах для некоторых видов характерно наличие приемной камеры, представляющей прообраз предсердия, для других — появление клапанов. Сокращение может охватывать все трубкообразное сердце целиком или распространяться волнообразно. Кровь в такое сердце обычно поступает через парные отверстия, оттекает по одной или нескольким артериям.
Следующим этапом филогенеза являются камерные сердца моллюсков и позвоночных. Это уже более совершенный тип насоса, состоящий из предсердий, желудочков и клапанного аппарата. У моллюсков и рыб сердце двухкамерное, состоящее из предсердия с предшествующим ему венозным синусом и желудочка, от которого отходит артериальный сосуд — брюшная аорта. Сердце рыб перекачивает только венозную кровь, поступающую от органов. От сердца кровь гонится к жабрам, где она обогащается кислородом.
Эволюция двухкамерного сердца начинается с двоякодышащих рыб. Появление легких у земноводных сопровождается образованием двух предсердий, и сердце становится трехкамерным. У этих животных возникают передние и задние полые вены, отводящие кровь от тела в правое предсердие. У высших позвоночных и человека появились четырехкамерные сердца с полным разделением потоков артериальной и венозной крови (рис. 8.2). Они будут рассмотрены ниже.
Ампулярные сердца существуют как добавочные к основным. Они располагаются преимущественно в местах повышенного сопротивления сосудов, где для прокачивания крови требуется создание повышенного давления. У моллюсков, например, они находятся
вжабрах, у насекомых — в местах прикрепления крыльев и конечностей.
8.2.1.Общие принципы строения
Увысших позвоночных (рис. 8.3) сердце состоит из двух половин: левой (системной) и правой (легочной). Их функциональное разделение происходит только после рождения. В каждой половине находятся предсердие и желудочек. Предсердие и желудочек соответствующей половины соединены между собой атриовентрикулярным отверстием, снабженным в левой половине двустворчатым, в правой — трехстворчатым клапанами
(рис. 8.4).

Рис. 8.3. Сердце млекопитающих (поперечный разрез): 1 — левое предсердие, 2 — ветви левой легочной вены, 3 — париетальный листок перикарда, 4 — полость перикарда, 5 — митральный клапан, 6 — висцеральный листок перикарда (эпикард), 7 — миокард, 8 — эндокард, 9 — левый желудочек, 10 — верхушка, 11 — межжелудочковая перегородка, 12 — правый желудочек, 13 — трехстворчатый клапан, 14
— правое предсердие
Рис. 8.4. Сердце млекопитающих (внутреннее строение): 1 — плечеголовной ствол, 2 — левая общая сонная артерия, 3 — левая подключичная артерия, 4 — аорта, 5 — ветви левой легочной артерии, 6 — левые легочные вены, 7 — левое предсердие, 8 — двустворчатый (левый атриовентрикулярный) клапан, 9 — аортальный полулунный клапан, 10 — левый желудочек,11 — межжелудочковая перегородка, 12 — верхушка сердца, 13 — нисходящая аорта, 14 — правый желудочек, 15 — сосочковая мышца, 16 — нижняя полая вена, 17 — сухожильные струны, 18 — трехстворчатый (правый атриовентрикулярный) клапан, 19
— правое предсердие, 20 — легочный полулунный клапан, 21 — правые легочные вены, 22 — ветви правой легочной артерии, 23 — верхняя полая вена; стрелками на красном фоне обозначено артериальное кровообращение, на синем — венозное
Со стороны желудочков к клапанам прикрепляются сухожильные нити, что позволяет клапанам открываться только в сторону желудочков. Помимо клапанов отверстия имеют кольцевые мышцы, участвующие в замыкании отверстий. От левого желудочка отходит аорта, которой начинается большой круг кровообращения, а от правого желудочка — легочная артерия, являющаяся началом малого, или легочного, круга кровообращения. Отверстия, которыми начинаются эти сосуды, закрыты полулунными клапанами, открывающимися только во время сокращения желудочков.
Стенка сердца состоит из трех слоев: эндокарда, миокарда (см. рис. 8.3) и эпикарда. Основную массу составляет миокард, имеющий наиболее сложное строение. Его

образуют отдельные мышечные волокна, каждое из которых является функциональной единицей. Миокард представляет собой цепочку соединенных последовательно (конец в конец) клеток, имеющих общую мембрану. Ткань миокарда, сохраняя сходство с поперечно-полосатой скелетной мышечной тканью, существенно отличается от нее рядом признаков, в том числе особой насыщенностью кардиомиоцитов митохондриями, что отражает высокий уровень метаболизма ткани, обладающей непрерывной активностью.
Поперечный срез, сделанный через середину обоих желудочков, указывает на значительно большую толщину левого. Различия касаются также и внутреннего строения. Стенки левого желудочка представляют собой мощный цилиндр из циркулярной мускулатуры, покрытый снаружи и изнутри спиральными волокнами. В правом желудочке циркулярный слой развит относительно слабо, а основную массу составляют спиральные волокна. Такие различия в строении отражают функциональные особенности, т. е. те усилия, которые развиваются каждым из желудочков.
В миокарде кроме сократительных, или рабочих, волокон имеется особая система мышечных единиц, обладающих способностью к генерации спонтанной ритмической активности, распространению возбуждения ко всем мышечным слоям и координации последовательности сокращения камер сердца. Эти специализированные мышечные волокна составляют проводящую систему сердца (рис. 8.5).
Рис. 8.5. Проводящая система сердца (А) и потенциалы действия различных отделов проводящей системы (Б): 1 — синусный узел, 2 — ножки пучка Гиса, 1 — атриовентрикулярный узел, 4 — пучок Гиса; стрелками показано направление распространения возбуждения; I — ПД синусного узла, II — ПД миокарда желудочков, МДД — медленная диастолическая деполяризация

Волокна рабочего миокарда соединяются друг с другом с помощью вставочных дисков
— нексусов, обладающих незначительным электрическим сопротивлением. Они служат местом перехода возбуждения между клетками, обеспечивая функциональное единство миокарда.
Таким образом, сердечная мышечная ткань ведет себя как функциональный синцитий. Эта особенность организации является основой для проявления закона «все или ничего». Суть этого закона состоит в том, что при действии раздражителя сердце либо отвечает сокращением всех рабочих мышечных волокон, либо (если раздражитель подпороговый) не отвечает совсем (рис. 8.6).
Рис. 8.6. Сократительные свойства сердечной мышцы (закон «все или ничего») (А) и скелетной мышцы
(градуалность) (Б): I – амплитуда сокращений, II — сила раздражения
Этим свойством сердечная мышца отличается от скелетных мышц и нервов, где каждая клетка возбуждается изолированно.
8.2.2.Свойства сердечной мышцы
Косновным свойствам сердечной мышцы относятся автоматия, возбудимость, проводимость, сократимость.
Автоматия сердца. Способность к ритмическому сокращению без всяких видимых раздражений под влиянием импульсов, возникающих в самом органе, является характерной особенностью сердца. Это свойство называется автоматизмом. В том случае, когда импульсы появляются в мышечных волокнах или их эквивалентах, сердечная деятельность считается миогенной, если же импульсы возникают в клетках нервных ганглиев сердца —
нейрогенной.
В миогенном сердце перерезка всех идущих к нему, внешних нервов не прекращает автоматизма, а извлеченное из тела сердце теплокровных и особенно холоднокровных животных при создании необходимых условий способно сокращаться в течение нескольких часов и даже суток. Ритмическое сокращение сердца проявляется уже на ранних стадиях эмбрионального развития (у человеческого эмбриона — на 18—20-й день). Так же ритмичейки сокращаются сердечные клетки эмбриона в культуре ткани.
Рассматривая вопрос о координации сердечного цикла с эволюционной точки зрения, В. Гаcкелл установил, что для червей, имеющих сердце трубчатого типа, характерно кольцеобразное расположение мышечных волокон со способностью их к ритмическим сокращениям. Дальнейшее филогенетическое развитие сердца шло за счет образования мощной мышечной массы, при этом утрачивалась способность всей массы к автоматической ритмике. И только в определенных участках сердца оставались, еще более совершенствуясь, группы мышечных волокон, сохранившие первичное свойство автоматизма. Они имеют особое гистологическое строение, отличное от волокон рабочей мускулатуры. За этими волокнами установилось название aтипическая мышечная ткань.
У холоднокровных эти волокна идут правильными рядами и кольцеобразно охватывают венозный синус. Они сконцентрированы также в атриовентрикулярном мостике и бульбусе аорты. На границе предсердий и желудочка волокна образуют спираль и далее спускаются по желудочку к верхушке сердца. Способностью к спонтанной генерации ритма обладают более примитивные клетки, а не высокоспециализированные рабочие кардиомиоциты. На ранних стадиях онтогенеза ритмической активностью обладают все клетки закладки сердца.
Природа автоматии до сих пор до конца не выяснена. У высших позвоночных и птиц возникновение импульсов связано с функцией атипических мышечных клеток — пейсмекеров, заложенных в узлах сердца. Нервные структуры способны оказывать влияние на силу и частоту их разрядов, однако сам процесс генерации импульсов является специфической особенностью этих клеток.
Атипическая ткань в сердце птиц и млекопитающих животных локализуется в областях, гомологичных венозному синусу и атриовентрикулярной области холоднокровных. Первый узел проводящей системы расположен в месте впадения полых вен в правое предсердие — синусно-предсердчый (синусный, синоатриальный, синусноаурикулярный, Кейт-Флака) узел (см. рис. 8.5). Он является главным центром автоматики сердца — пейсмекером первого порядка.
От узла возбуждение распространяется к рабочим клеткам миокарда предсердий и диффузно, и по специальным внутрисердечным проводящим пучкам. Оба потока достигают второго узла — не менее нажного места, обладающего способностью генерировать импульсы, — атриовентрикулярного (предсердно-жедудочкового, Ашоффа
— Товара) узла. Он расположен в толще сердечной перегородки на границе предсердий и желудочков. Узел состоит из трех обладающих собственной частотой возбуждения частей: верхней предсердий, средней и нижней желудочковых. Этот узел является пейсмекером второго порядка. Возбуждение через предсердно-желудочковый узел в нормальных условиях может проходить только в одном направлении. Ретроградное проведение импульсов невозможно.
Так достигается направленность движения возбудительного процесса и, как следствие, координированность работы предсердий и желудочков.
При прохождении возбуждения через предсердно-желудочковый узел импульсы задерживаются на 0,02—0,04 с. Это явление получило название итриовентрикулярной задержки. Ее функциональное значение состоит в том, что за время задержки успевает завершиться систола желудочков и их волокна будут находиться в фазе рефрактерности.
Третий уровень, обеспечивающий ритмическую активность сердца, расположен в пучке Гиса и волокнах Пуркинье. Пучок Гиса берет начало от предсердно-желудочкояого узла и образует две ножки, одна из которых идет к левому, другая — к правому) желудочку. Эти ножки ветвятся на более тонкие проводящие пути, заканчивающиеся волокнами Пуркинье, которые непосредственно контактируют с рабочими клетками миокарда.
Центры автоматии, распложенные в проводящей системе желудочков, носят название пейсмекеров третьего порядка. Таким образом, возбуждение по ножкам пучка Гиса направляется к верхушке сердца и оттуда по разветвлениям ножек и волокнам Пуркинье возвращается к основанию сердца. В результате этого сокращение сердца в целом осуществляется в определенной последовательности: сначала сокращаются предсердия, затем верхушки желудочков и, наконец, их основания. Пучок Гиса и волокна Пуркинье отсутствуют у пойкилотермных животных. Возбуждение у них распространяется по волокнам миокарда.
Перечисленные водители ритма сердца находятся в соподчиненном положении. В сердце существует так называемый градиент автоматии. Он выражается в убывающей способности к автоматии различных структур проводящей системы по мере их удаления от синусно-предсердного узла. В синусно-предсердном узле число разрядов составляет в среднем 60-80 имп/мин, в предсердно-желудочковом — 40—50, в клетках пучка Гиса —
30—40, в волокна Пуркинье — около 20 имп/мин.
Таким образом, в сердце существует определеннаяиерархия центров автоматии, что позволило В. Гаскеллу сформулировать правило, согласно которому степень автоматии отдела тем выше, чем ближе он расположен к синусно-предсердному узлу. Это правило получило название закона градиента сердца.
Вобычных условиях частоту активности миокарда всего сердца в целом определяет синусно-предсердньш узел. Он подчиняет все нижележащие образования проводящей системы, навязывая им свой ритм. Поэтому все отдельные части проводящей системы, хотя и имеют собственный ритм, начинают работать в едином ритме. Явление, при котором структуры с замедленным ритмом генераций потенциалов усваивают более частый ритм других участков проводящей системы, называют усвоением ритма.
Следует заметить, что в областях узлов — синусно-предсержном, предсердножелудочковом — находятся нервные клетки. Их скопления и многочисленные волокна образуют густую нервную сеть, пронизывающую структуру узла. Эти нервные клетки относятся к кардиальной части метасимпатической нервной системы (см. разд. 5.4.3).
Вмышечных волокнах предсердий и желудочков скорость проведения возбуждения колеблется в узких пределах, составляя 0,9-1,0 м/с, в волокнах предсердно-желудочкого узла — 0,05, в пучке Гиса — 1,0—1,5, в волокнах Пуркинье — 3,0 м/с. Быстрое проведение в волокнах Пуркинье определяет почти одновременное возбуждение всех участков желудочков. Время полного охвата, составляет около 10-15 мс. В связи с этим возрастают мощность сокращения и эффективность работы, связанная с проталкиванием желудочком крови.
Для обеспечения работы сердца необходимым условием является анатомическая целостность его проводящей системы. В том случае, если в пейсмекере первого порядка
всилу каких-то причин не возникает возбуждение или блокируется его передача, роль водителя берет на себя пейсмекер второго порядка. В случае же невозможности передачи возбуждения к желудочкам они начинают сокращаться в ритме пейсмекеров третьего порядка. При поперечной блокаде предсердия и желудочки сокращаются каждый в своем ритме. Повреждение водителей ритма ведет к полной остановке сердца.
На начальных стадиях эволюции встречается сугубо нейрогенный механизм регуляции сердечного ритма, так называемое нейрогенное сердце. Например, у большинства ракообразных на дорсальной поверхности сердца расположено несколько ганглиозных нервных клеток. Это генераторы ритма, т. е. пейсмекерные клетки сердца. Возникающее в них возбуждение широкой волной разливается на мышечные клетки, вызывая их сокращение. Частота генерации находится в прямой зависимости от наполнения сердца, т. е. состояния растянутости его стенок. Подобные ганглионарные нервные клеткипейсмекеры есть и в сердце некоторых насекомых: у взрослых пчел, тараканов. Обнаружены они у ряда кольчатых червей. Удаление ганглиев приводит к немедленной остановке сердца. Нейрогенные сердца чрезвычайно чувствительны к эфиру. Даже малые его концентрации вызывают остановку сокращения сердца.
Клетки атипической мышечной ткани, составляющие проводящую систему сердца, функционально неоднородны. Из всей массы синусно-предсердного узла только несколько клеток, называемых истинными пейсмекерами, обладают способностью к спонтанной генерации потенциала действия. Остальные клетки относятся к потенциальным (латентным) водителям ритма. Они, как и рабочие кардиомиоциты, разряжаются в результате пришедшего к ним возбуждения. Потенциальные пейсмекеры отличаются от истинных особенностями медленной диастолической деполяризации и более низкой частотой разрядов. В свою очередь, от рабочих миокардиоцитов они отличаются тем, что последние вообще не способны к спонтанной деполяризации. В случае прекращения функции истинного пейсмекера потенциальный пейсмекер может взять на себя его ведущую роль, в то время как рабочие кардиомиоциты такой способностью не обладают.
Особенностью механизма возникновения ритмов истинных и потенциальных пейсмекеров является то, что в отличие от волкоон сократительного миокарда мембрана этих клеток во время диастолы приобретает большую ионную проницаемость, что ведет к развитию медленной диастолитической деполяризации — пейсмекерного потенциала. В
этот момент возникает местное нераспространяющееся возбуждение. У потенциальных пейсмекеров эта фаза достигает порогового уровня позже, чем у истинных. С достижением диастолитического порогового уровня происходит возникновение распространяющегося ПД.
Ионный механизм пейсмекерного потенциала состоит в том, что во время фазы реполяризации клеточная мембрана становится более проницаемой для внутриклеточного К+ и мембранный потенциал приюлижается к равновесному калиевому потенциалу Ек. В результате проникновения внутрь клетки Na+ и Са2+ и уменьшения скорости выхода из клетки К+ возникает меделенная диастолическая деполяризация. Когда уровень потенциала уменьшится посравнению с исходным приблизительно на 2 мВ, наступает резкое увеличение проницаемости мембраны вначале для Na+ а позже для Са2+. Этот ионный ток приводит к возникновению пика ПД. Общая амплитуда ПД составляет около 100 мВ. С закрытием ионных каналов положительный заряд наружной поверхности мембраны восстанавливается. Появление ПД у пейсмекерной клетки сопровождается возникновением деполяризации у прилежащих к ней ведомых рабочих кардиомиоцитов, не обладающих автоматизмом, и распространением возбуждения. Рабочие клетки миокарда в отличие от водителей ритма в состоянии покоя характеризуются чрезвычайно низкой ионной проницаемостью для Na+ и Са2+, так что движение через их каналы К+ не вызывает сдвигов мембранного потенциала.
Ритм работы сердца зависит от массы животного и уровня метаболизма. Как правило, частота сердечных сокращений у животных с низкой общей подвижностью меньше, чем у подвижных. У улиток она колеблется от 0,2 до 20 в 1 мин, у кальмара, и осьминога — от 40 до 80. У мелких животных частота сердечных сокращений, как правило, выше, чем у крупных. Например, частота сокращений у мелких птиц составляет несколько сотен в 1 мин, у домашних — 150—300, у мыши — от 300 до 500, кролика — 200, кошки — 125, собаки — 80, лошади, слона — 25-40
Зависимость между величиной животного и частотой сердечных сокращений объясняется более высоким уровнем метаболизма мелких животных. Высокий уровень может обеспечиваться только интенсивным кровоснабжением органов и тканей, а, следовательно, более быстрым режимом работы сердца. Частота сердечных сокращений резко усиливается при мышечной работе. У ныряющих млекопитающих, птиц и черепах во время ныряния наблюдается урежение сердечной деятельности.
Возбудимость сердечной мышцы. Под действием электрических, химических, термических и других раздражителей сердце способно приходить в состояние возбуждения. В основе процесса возбуждения лежит появление отрицательного электрического тенциала в первоначально возбужденном участке.
Как и у любой возбудимой ткани, мембрана рабочих клеток сердца поляризована. Снаружи она заряжена положительно, изнутри отрицательно. Это состояние возникает в результате разной концентрации Na+ и К+ по обе стороны мембраны, а также разной проницаемости мембраны для этих ионов. В покое мембрана кардиомиоцитов почти непроницаема для Na+ и частично для К+. В результате процесса диффузии ионы К+, выходя из клетки, увеличивают положительный заряд на ее поверхности. Внутренняя сторона мембраны при этом становится отрицательной. В миокарде теплокровных разность потенциалов в покое (потенциал покоя) составляет 60—80 мВ. При действии раздражителя любой природы, прихода возбуждения от соседней клетки или пейесмера происходит поступление Na+ в клетку. В этот момент на поверхности мембраны возникает отрицательный электрический заряд и развивается реверсия потенциала. Амплитуда ПД составляет 100 мВ и более. Возникший потенциал деполяризует мембраны соседних,