

Книги по МРТ КТ на английском языке / The Embryonic Human Brain An Atlas of Developmental Stages. Third Edition. 2006. By Ronan O'Rahilly
.pdf
C H A P T E R 20
STAGE 20: THE CHOROID
PLEXUS OF THE LATERAL
VENTRICLES, THE OPTIC
AND HABENULAR
COMMISSURES, AND THE
INTERPEDUNCULAR AND
SEPTAL NUCLEI
Approximately 18–22 mm in Greatest Length;
Approximately 47 Postfertilizational Days
The globus pallidus externus is distinct in the subthalamic area. The optic commissure is present, and crossing fibers begin to form in the future habenu-
lar commissure. The medial septal nucleus and the nucleus of the diagonal band are differentiating, and fiber connections with the hippocampus and the medial prosencephalic fasciculus are present. Important fiber connections of the olfactory system are now identifiable: from the olfactory bulb to the olfactory tubercle, to the septal nuclei, to the amygdaloid nuclei and via the stria medullaris thalami to the habenular nuclei; then from there via the habenulo-interpeduncular tract to the interpeduncular nucleus. Connections exist also from the mesencephalic tegmentum via the medial prosencephalic fasciculus to the amygdaloid and septal nuclei and olfactory tubercle. The
choroid plexuses of the lateral ventricles at stage 20 are at the “club-shaped” phase. Cuneate and gracile decussating fibers are present. Histological differences are now apparent between perimural arteries and veins (although not in the intramural vessels until trimester 2).
Catecholaminergic cell groups have been detected at as early as 5 weeks in the rhombencephalon and mesencephalon, and at 6 weeks in the hypothalamus (Verney et al., 1991). See also Figure 17–13 and Table 17–3. A band of densely packed cells that corresponds to the primordium of the dopaminergic substantia nigra and ventral tegmental region has been recorded at 7 weeks (Verney et al., 1991). Nigrostriatal fibers develop at this time and extend initially to the subventricular zone of the lateral ventricular eminence (Freeman et al., 1995).
The Embryonic Human Brain: An Atlas of Developmental Stages, Third Edition. By O’Rahilly and Muller¨ Copyright C 2006 John Wiley & Sons, Inc.
169

170 C h a p t e r 2 0 : THE CHOROID PLEXUS THE LATERAL VENTRICLES, THE OPTIC AND HABENULAR COMMISSURES
M
Di.
T
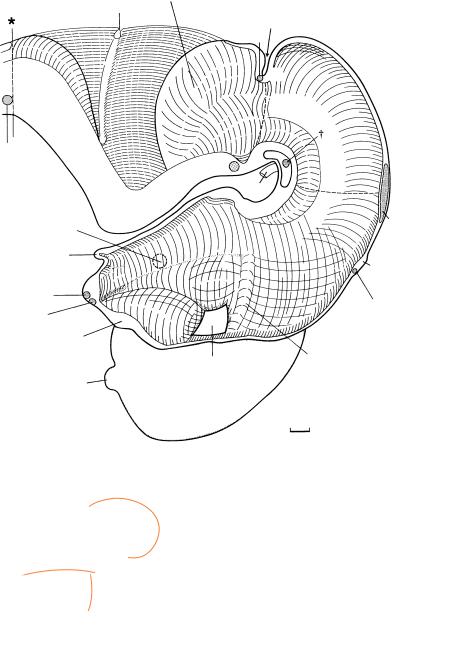
Figure 20–1. Right lateral view of the brain. The frontal, occipital, and temporal poles are indicated (by α, β, γ ). The tuberculum interpedunculare (of Hochstetter), which faces the mamillary region, is particularly evident in this embryo (dagger). The roof of the fourth ventricle leaves a part of the cerebellum free (ausserer¨ Kleinhirnwulst of Hochstetter, 1929). The ventricular cavity between the internal cerebellar swelling and the caudal part of the rhombencephalon has become very narrow. The cranial nerves have been cut short. A mesencephalic Blindsack (the site of the future inferior colliculus) is visible on each side of the mesencephalon and almost touches the cerebellum.
In a dorsal view in the left-hand inset, the cerebellar hemispheres are seen to be separated from each other. Dorsally, the left and right halves of the cerebellum are joined only in the area of the rostral (superior) medullary velum.
The right-hand inset is an end-on view showing the cerebral hemispheres, part of the diencephalon with the epiphysis, and the mesencephalon.
The neocortical development of an embryo belonging to stage 20 was illustrated by Mar´ın-Padilla (1983, 1988a). At this stage the intrinsic vascularization of the neocortex begins. “It evolves from perforating capillaries of the pia-vascular plexus, and progressively advances in ventro–dorso–lateral and antero–posterior [rostrocaudal] gradients, paralleling the differentiation and maturation of the neocortex” (Mar´ın-Padilla, 1988a). The intraneural vascular territories of the cerebral cortex in the embryo and in the adult are quite similar. Both are characterized by a short-link anastomotic network of intracortical capillaries established between adjacent perforating vessels (Mar´ın-Padilla, 1988b). Certain portions of the primordial plexiform layer are considered to be functionally active in the embryo by this stage (Mar´ın-Padilla and Mar´ın-Padilla, 1982).
Is has been found in the cat that corticipetal and some corticofugal fibers are present prior to the appearance of the cortical plate.
Cerebellar hemisphere
Plexus fold
Isthmic groove
X4
Lemniscal decussation
3
Globus pallidus ext.
N.-H.
Supraoptic
X
Ch.
Comm.
l.-v.f.
Olfactory buld
0.4
(a) |
Sulcus |
a |
limitans |
|
|
(b) |
Hypothalamic |
|
sulcus |
b |
|
(c) |
Marginal |
c |
ridge |
|
Post.
X
Ep.
Habenular X
Marginal ridge
Figure 20–2. Graphic reconstruction prepared from coronal sections to show a median view of the brain. The asterisk indicates the junction with the spinal cord.
The choroid plexuses of the lateral ventricles at stage 20 are at the “club-shaped” phase described by Shuangshoti and Netsky (1966). In the diencephalon, the position of the marginal ridge no longer corresponds to that of earlier stages. A ventral shift has occurred, whereby the dorsal thalamus has become larger towards the interventricular foramen. The earliest part of the globus pallidus, the globus pallidus externus, appears at the surface of the subthalamus in this and in the previous stage. Subsequently it moves into the hemispheric stalk. The optic chiasma lies rostroventral to the decussating fibers of the preoptico-hypothalamotegmental tract, and these form the supraoptic commissure. In most embryos of this stage, the epiphysis cerebri is immediately rostral to the posterior commissure. The caudal shift is probably caused by a growth gradient. It is likely that the habenular commissure, containing a few fibers, is present in most embryos of this stage. The habenulointerpeduncular tract is no longer at the surface but runs between the ventricular and intermediate layers of the diencephalon and arrives in the mesencephalon lateral to the interpeduncular nucleus (Fig. 20–12).
The commissural fibers of the oculomotor nerves (dagger) are joined by the crossing fibers of the dentatorubral tract. This is the beginning of the decussation of the superior cerebellar peduncles.
The basal plate of the rhombencephalon has increased greatly in thickness, especially in its caudal part. The lateral portions of the rhombencephalon are depressed in the region of the pontine flexure and hence are not visible in this view. The cerebellar hemispheres are conspicuous. The villi of the choroid plexus of the fourth ventricle extend from the choroid fold into the wall of the lateral recesses. The septum medullare (median raphe of His, 1890) begins at the isthmic recess. It is derived from the rhombencephalic floor plate and is important in the production of neurotransmitters.
Cuneate and gracile decussating fibers are present at the transition from rhombencephalon to spinal cord.
172 C h a p t e r 2 0 : THE CHOROID PLEXUS THE LATERAL VENTRICLES, THE OPTIC AND HABENULAR COMMISSURES
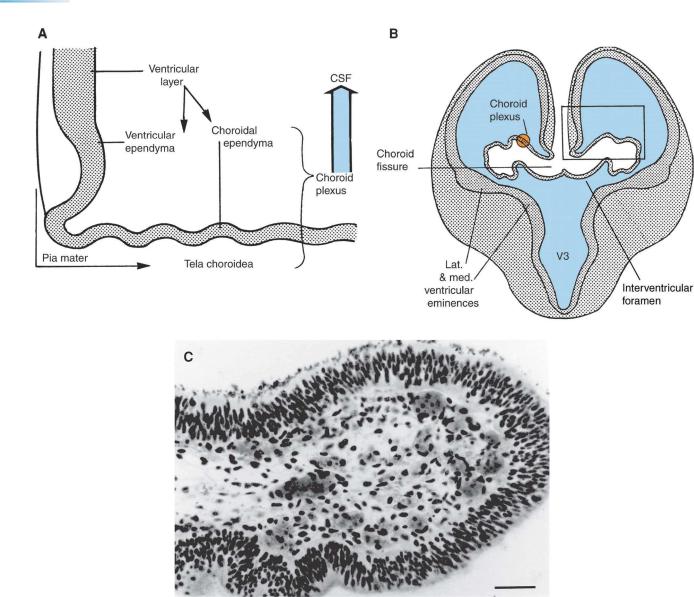
Figure 20–3. Sections through the prosencephalon. (A) A portion of the wall of the brain from the rectangle indicated in B. The ventricular layer gives origin to the ventricular ependyma (that lining the ventricle) and to the choroidal ependyma (of the choroid plexus), and the two are continuous with each other. The choroid plexus consists of choroidal ependyma and tela choroidea; the latter is a vascularized pia mater.
(B) Transverse section showing that the interventricular foramina are now narrow. The choroid plexus extends into the lateral ventricle on each side, and its line of invagination is the choroid fissure. The ependyma of the plexus is continuous with the medial wall of the lateral ventricle. (C) An enlargement of a portion of the choroid plexus at stage 19 from the rectangle indicated in B. The epithelium is the choroidal ependyma, the layers of which enclose the vascular pia mater known as the tela choroidea. Bar: 0.05 mm.
THE CHOROID PLEXUS THE LATERAL VENTRICLES, THE OPTIC AND HABENULAR COMMISSURES |
173 |
|
|
|
|
Dural |
|
|
|
|
limiting |
|
|
|
|
layer |
|
|
|
|
Pia |
|
|
Hippocampal |
Primordial |
|
|
|
plexiform |
||
|
|
primordium |
|
|
|
|
|
layer |
|
|
|
|
|
|
|
|
|
|
Ventricular |
|
|
Area |
|
layer |
|
|
Choroid |
|
|
|
|
epithelialis |
|
|
|
|
plexus |
|
|
|
|
|
|
|
|
|
|
|
Lat. ventricular |
|
|
|
|
eminence |
Roof |
Choroid |
|
|
|
of 3rd |
|
|
|
|
fissure |
|
|
Subventricular |
|
ventricle |
|
|
layer |
|
|
Med. |
|
|
|
ventricular |
|
Lat. forebrain |
|
|
eminence |
|
Subarachnoid |
||
|
bundle |
|||
|
|
|
space |
|
|
|
|
|
|
|
|
|
|
|
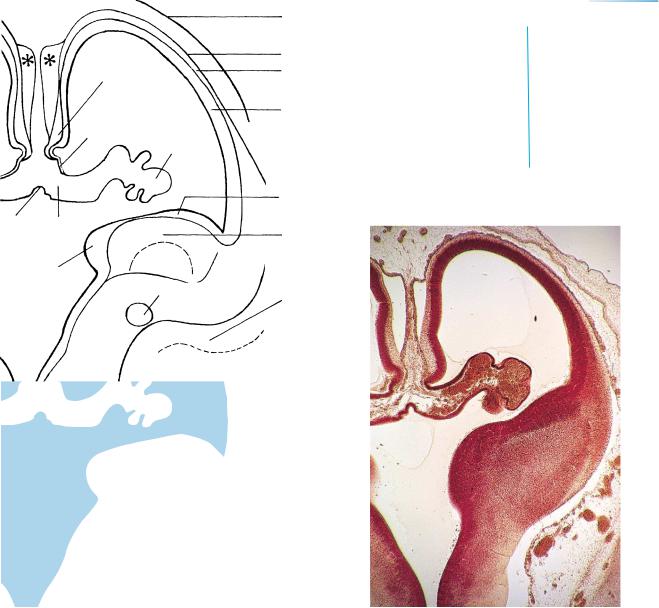
Figure 20–4. An oblique section through one cerebral hemisphere. The third and lateral ventricles are united by the interventricular foramen. The line of invagination of the prominent choroid plexus into the lateral ventricle is the choroid fisssure. Noteworthy is the absence of the corpus callosum, which has not yet developed. The asterisks are placed on artifactual spaces.
174 C h a p t e r 2 0 : THE CHOROID PLEXUS THE LATERAL VENTRICLES, THE OPTIC AND HABENULAR COMMISSURES
Figure 20–5. Section through the nasal epithelium, from which crest cells form cords that enter the mesenchyme and proceed towards the basal forebrain.
1 
5
3 
2
4
Figure 20–6. Telencephalic migration. Radial migration (1) to the cortex, (2) and (3) to the amygdaloid nuclei. Tangential migration (4) and (5) to the cortex.
|
|
|
Motor end |
|
Axon |
plate |
|
Oligodendrocyte |
|
|
|
|
|
|
|
Neural crest |
Neurilemma |
|
|
A
Pia mater
Germinal or |
blast |
cell |
plexus |
ventricular cells |
|
||
|
|
cell |
|
|
VENTRICLE |
|
|
|
|
|
plexus Choroid
Figure 20–7. A portion of the wall of the neural tube showing the main cells. (A) The germinal or ventricular cells in the ventricular or matrix zone (1) give rise to
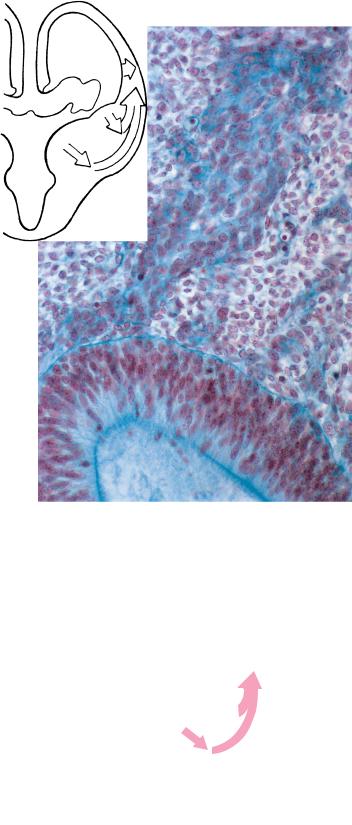
(a) immature and eventually mature neurons, (b) glioblasts (precursors of astrocytes, oligodendrocytes, and radial cells, which are essential for radial migration; Fields and Stevens-Graham,´ 2002), and (c) ependymoblasts, which develop into ependymal and choroid plexus cells. According to an alternative theory, neurons and glioblasts arise from different cells in the ventricular zone. Moreover, it is now maintained that glial cells can give rise to neurons. Above, an axon is shown surrounded by a neurilemmal sheath consisting of neurilemmal cells (derived from neural crest). 1, ventricular zone; 2, intermediate layer; 3, marginal layer.
(B) The choroid plexus consists of choroidal ependyma and a vascular pia termed the tela choroidea (cf Fig. 20–3A, C).
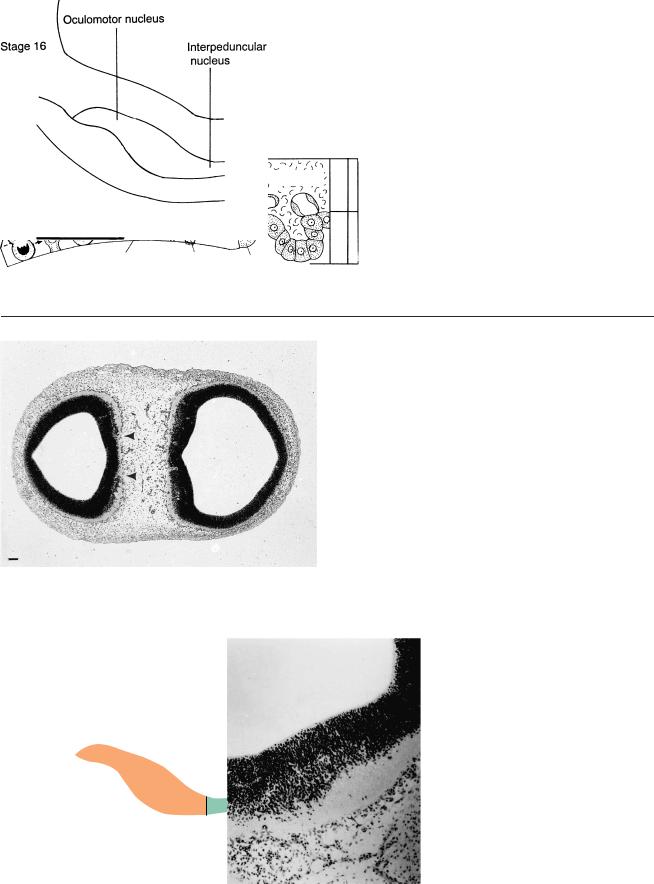
Stage 15
Figures 20–8 to 20–12 show the development of the interpeduncular nucleus from 5 to 7 weeks. This nucleus is important for neural grafts as a possible treatment for Parkinson disease (Freeman et al., 1991; Freed et al., 1992). The bars represent 0.1 mm.
M2 |
Di. |
Figure 20–8. At stage 15 a ventral proliferative area of the mesencephalon, between the right and left oculomotor nuclei, is already visible (arrowheads).
Figure 20–9. At stage 16 the loose material between the oculomotor nuclei has increased to several rows of cells.

176 C h a p t e r 2 0 : THE CHOROID PLEXUS THE LATERAL VENTRICLES, THE OPTIC AND HABENULAR COMMISSURES
Figure 20–10. At stage 17 transverse fibers ventral to the proliferative area represent the commissure of the oculomotor nerves.
20 – 12
20 – 11
Figure 20–11. At stage 20 the fibers passing through the tegmentum of the mesencephalon have increased to such an extent that the cerebral peduncles and the interpeduncular groove have begun to appear. The interpeduncular nucleus is visible, and the nucleus niger is lateral to it. The neurons of the substantia nigra are believed to be generated (in the rat) in the region of the isthmus rhombencephali. Transverse fibers, which are beginning to penetrate the loose cellular sheath, probably are rubrocerebellar fibers belonging to the superior cerebellar peduncles (Cooper, 1946a). The oculomotor nuclei are visible.
THE CHOROID PLEXUS THE LATERAL VENTRICLES, THE OPTIC AND HABENULAR COMMISSURES |
177 |
Stage 21 |
Figure 20–12. At stage 21 |
|
habenulo-interpeduncular fibers are seen to |
||
|
||
|
arrive in the lateral part of the |
|
|
interpeduncular nucleus. The substantia |
|
|
nigra is lateral to those fibers. The |
|
|
commissure of the superior colliculi is |
|
|
visible dorsally in the roof. Mesencephalic |
|
|
dopaminergic neurons have been detected at |
|
|
approximately this stage (Freeman et al., |
|
|
1991). The red nucleus, which is already |
|
|
distinguishable at stage 17, is lateral and |
|
|
slightly dorsal to the oculomotor nucleus. |
|
|
The ventral tegmental region is dorsal to the |
|
|
interpeduncular nucleus, and its fibers |
|
|
ascend in the medial forebrain bundle. |
|
|
Mesencephalic dopaminergic neurons, |
|
|
especially of the ventral tegmental area, |
|
|
project to the ventral striatum (Vernier et al., |
|
|
2004). |
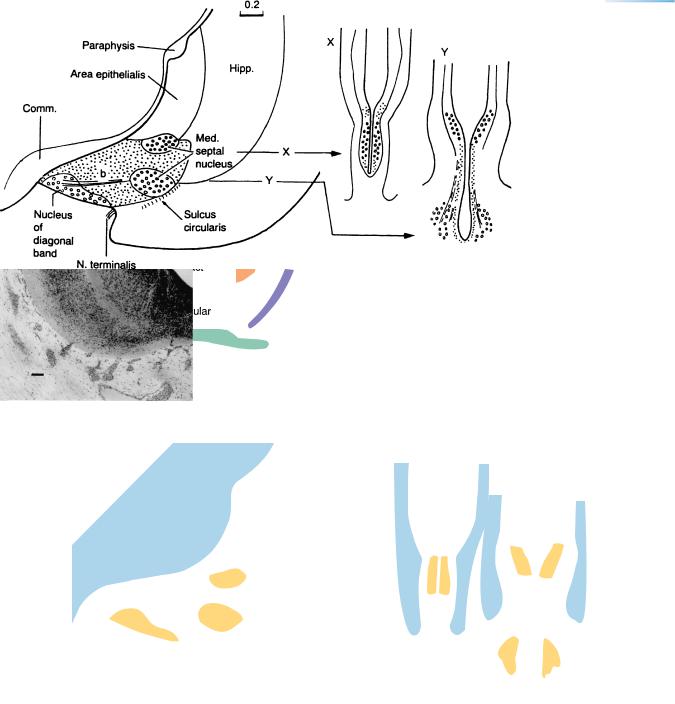
Figure 20–13. A graphic reconstruction and two transverse sections. The medial septal nucleus is near the hippocampus and includes two separate portions, X and Y. The nucleus of the diagonal band is ventrocaudal and occupies the middle third of the septum. Nerve fibers are present between the nucleus and the hippocampus.
The nucleus of the diagonal band is identified by its projecting fibers (b) to the hippocampus. The caudalmost part of this area, however, is less clear. In the monkey (studied by thymidine marking), the production of cells of the septal nuclei (medial and nucleus of the diagonal band) is almost simultaneous with that of the nucleus basalis (of Meynert). This would correspond approximately to Carnegie stage 14, followed by a second wave between stages 18 and 22. The three functional cholinergic groups of the basal forebrain (medial septal, nucleus of the diagonal band, and nucleus basalis) “are the most outstanding sets of neurons regulating cortical functions. Each group specifically ends in a part of the cortex,” and their number and function are “statistically diminished in AD {Alzheimer disease} patients” (Toledano, 1992).