

Книги по МРТ КТ на английском языке / Functional Neuroimaging in Child Psychiatry Ernst 1 ed 2000
.pdf144 D. M. Pascualvaca and G. Morote
Table 8.4. Characteristics of diVerent memory systems
Memory systems |
Other terminology |
DeWnition |
Examples of tasks |
|
|
|
|
Short-term memory |
Working memory |
Capacity to retain information for |
|
|
brief periods of time |
Repetition of information (e.g., digits,
letters, and words); immediate replication of spatial sequences; mental manipulation of stimuli for brief intervals
Episodic memory |
Autobiographical |
Recollection of past happenings |
Remembering speciWc personal events; |
|
memory |
|
recalling signiWcant events in the individual's |
|
|
|
experience |
Semantic memory |
Factual knowledge |
Ability to remember novel information; |
|
|
acquisition and retention of factual |
|
|
material |
Recalling stories, scenes, facts; answering questions of general knowledge
Declarative memory |
Explicit memory |
Includes memory for facts and events |
Same as semantic memory |
Nondeclarative |
Implicit memory, |
Mastery of skills and habits |
Priming tasks |
memory |
procedural memory |
unconsciously processed that facilitate |
|
|
|
performance |
|
|
|
|
|
|
|
|
|
ally achieved (Hulme et al., 1984). There is considerable debate as to whether these developmental changes reXect an increase in memory capacity itself (e.g., Pascual-Leone, 1970) or are secondary to improved processing strategies. The strategies that have received most support include a growth in the use of phonologic processes (e.g., internally generated phonologic or sound codes), an increase in lexical knowledge (e.g., a growth in vocabulary), and the emergence of rehearsal techniques (e.g., rehearsing items subvocally) (for a review see Gathercole, 1998). It is likely that the sharpening of these strategies also accounts for the improvement observed in visual working memory tasks. For example, preventing subvocal rehearsal disrupts visual pattern span (e.g., memory for geometric forms, shapes and objects) in 11-year-old children more dramatically than in 5-year-old children (Mills et al., 1996). Consequently, in addition to possible increases in the capacity to hold visual information in memory, the older children use a range of verbal strategies to retain information that are not accessible to the younger groups. Spatial memory tasks (i.e., those that tap memory for the locations of the objects in space) show a less dramatic developmental progression than visual memory tasks. The diVerence between these two developmental trajectories has been attributed to the greater diYculty in mediating spatial information verbally (Gathercole, 1998). Recent PET data in adults have provided support for this hypothesis. SpeciWcally, Jonides et al. (1993) reported activation in language regions of the left hemisphere when adults were completing object memory tasks (i.e., recognition of novel geometric forms) that were not clearly observed on spatial memory tasks (e.g., dots in various location).
As with short-term memory, several systems have been hypothesized to subserve long-term memory in adults. For example, investigators have distinguished between episodic memory, or recollection of personal events and experiences, and semantic memory, or accumulated factual knowledge (e.g., Vargha-Khadem et al., 1997; Fernandez et al., 1998). The distinction between episodic and semantic memory has received relatively little attention in the developmental literature, in part because of the diYculty in assessing personal memories during the early years of life. Because of limitations in the assessment of memory functions in children, many investigators have conceptually organized memory into declarative and procedural systems, combining memories for facts and events into an integrated component. Declarative or explicit memory refers to the capacity for conscious recall of information and captures what the layman typically thinks of as memory. Procedural or implicit memory, by comparison, reXects expressed abilities (perceptual or motor) for which there is no conscious recollection. It includes the skills and habits that facilitate performance in novel tasks. For example, recall of a task is facilitated by prior exposure to a similar or identical task, even when the individual does not remember having had such exposure, a phenomenon known as priming. Studies in human adults and with animal models indicate that the declarative and procedural long-term memories are mediated by distinct brain pathways. Lesions of the medial temporal lobe, including the hippocampus, in nonhuman primates result in impairments in declarative memory, leaving performance on procedural or implicit memory tasks intact. Implicit memory is, in contrast, disrupted by lesions in areas of the
Cognitive development from a neuropsychologic perspective |
145 |
|
|
|
|
striatum (e.g., Bachevalier, 1990). The results of animal studies also suggest that these memory systems emerge at diVerent times in development, and that the system presumed to mediate declarative or explicit memory matures later in nonhuman primates (Bachevalier and Mishkin, 1984).
It is diYcult to determine when these memory systems emerge in humans. The paucity of knowledge regarding developmental changes reXects limitations in the paradigms that can be used to assess memory in young children. Nonetheless, the available data suggest that tasks that measure implicit memory can be mastered by infants as young as 31¤2 months of age (for a review see Nelson, 1997). The early emergence of these functions is consistent with the early maturation of structures that comprise the striatum (caudate and putamen) (Hudspeth and Pribram, 1992). Infants between 6 and 12 months of age can also complete simple explicit memory tasks. However, more complex explicit memory tasks (or tasks with an explicit memory component), such as the delayed nonmatch-to- sample task, are not mastered until at least 18 months (Overman et al., 1992, 1993; Diamond, 1995), presumably because of their increased reliance on cortical structures (Malkova et al., 1995; Fernandez et al., 1998). The developmental trajectory of explicit memory continues throughout childhood, possibly reXecting further organization in the temporal lobe and neighboring cortical areas. In contrast, it is possible that less dramatic development occurs in implicit memory because the underlying neural system matures earlier in life.
It is clear that infants are capable of a rich repertoire of complex memories. By as early as 9 months of age, infants can recall speciWc events over delays of several weeks and even months (for reviews see Bauer, 1997; Rovee-Collier and Gerhardstein, 1997). The amount of information and type of material that children can remember increases steadily over the Wrst few years of life. This development in memory capacities reXects several factors including (i) an improvement in the child's ability to encode more features of a stimulus, (ii) a decrease in the child's tendency to encode irrelevant information, (iii) an increase in the use of mnemonic strategies that promote learning, and (iv) an enhancement in metamemory or knowledge about memory.
In general, children's memories improve as their knowledge base expands (Pressley and Schneider, 1997). When children come to associate new information with a larger fund of knowledge, their memories become enriched. There are stronger connections among diVerent types of material, which makes this material more accessible and easier to recall. As the knowledge base continues to grow, it
facilitates the learning of new concepts that are related to concepts the child has already mastered.
During the school years, children also become increasingly adept at encoding the essential features of new stimuli and at registering fewer distracting events (Lane and Pearson, 1982). There is also a gradual improvement in the use of mnemonic strategies during childhood (for a review see Pressley and Schneider, 1997). Even 5-year-old children can use simple rehearsal strategies (e.g., rote repetition), but they do not rely on them consistently and may not derive as much beneWt from their use. It is not until 8 years of age that children's use of rehearsal has a notable impact on their performance. Eight-year-old children rehearse single items spontaneously to learn new material, and, by 10 years of age, they can group items together and begin to organize information into categories. They can also plan speciWc strategies to facilitate their recall. Organizational strategies develop somewhat later than rehearsal techniques and are not seen consistently until approximately 10 years of age. With increasing age, mnemonic strategies also become more complex. For example, the use of elaboration becomes evident only in adolescence. These new techniques do not necessarily replace the old ones, but rather the older strategies continue to develop and become more eYcient.
Another important aspect in the developmental progression of memory is a remarkable growth in children's knowledge about memory processes (metamemory, or the ability to make judgments and predictions about one's memory capacities). Young children are not aware of their memory capabilities (Kreutzer et al., 1975). For example, they tend to miscalculate the amount of time they require to learn a particular task and are not familiar with speciWc strategies that would facilitate their learning. During the school years, children gradually acquire knowledge about their memory capacities. For instance, they learn to allocate more time to material they have not yet mastered and are in a better position to appraise their readiness for tests. This knowledge clearly inXuences memory and learning, which, in turn, leads to enhanced metamemory (for a review see Schneider, 1997).
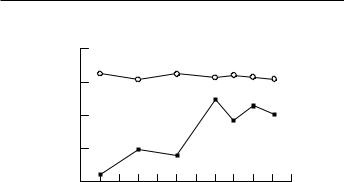
In addition to the processes that facilitate the encoding of new memories, there are also developmental diVerences in retrieval strategies, such as the use of categories and thematic cues to facilitate recall. The retrieval of memories grows steadily as other cognitive abilities mature. The diVerence between retrieval and recognition memories is a good example of how development in other cognitive systems inXuence recall. As shown in Fig. 8.3, 7-year-old children can recognize words in a list learning task as well as 16-year-olds, but they can spontaneously
146 D. M. Pascualvaca and G. Morote
|
16 |
|
|
|
|
|
|
|
|
|
|
|
of items |
14 |
|
|
|
|
|
|
|
|
|
|
|
12 |
|
|
|
|
|
|
|
|
|
|
|
|
Number |
|
|
|
|
|
|
|
|
|
|
|
|
10 |
|
|
|
|
|
|
|
|
|
|
|
|
|
8 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
17 |
|
6 |
Age (years)
Fig. 8.3. Comparison of developmental changes in free and recognition recall. Free recall, j; recognition recall, s. (Material from the Rey Auditory Verbal Learning Test: A Handbook copyright (1996) by Western Psychological Services. Reprinted by permission of the publisher. Western Psychological Services. 12031 Wilshire Boulevard, Los Angeles, California 90025, USA. All rights reserved.)
retrieve fewer items. The diVerence between performance in recognition and free-recall tasks has been attributed to an increase in language competency. It may well be, however, that the diVerence between these two paradigms also reXects the concurrent maturation of executive functions. This is evident by the improvement in organized search strategies (Jetter et al., 1986) and decrease in susceptibility to interference and intrusion errors (Fiducia and O'Leary, 1990) observed during the school-age years.
Language
Many critical events in neural development take place during the years of language acquisition.Yet, little is known about the underlying neural correlates of language development. Studies of children who have sustained trauma early in life have shown that the developing language system can undergo striking changes in neural organization. For example, the studies of Woods and Teuber (1973), Milner (Milner et al., 1964; Rasmussen and Milner, 1977), Bates (Reilly et al., 1998), Feldman (Feldman et al., 1992), and Kohn and Dennis (1971) all indicate that lesions and even resection of the left hemisphere do not preclude the development of language. The Wndings of these studies also suggest that the two hemispheres can mediate many aspects of language early in life. The bilateral representation of language early in life may be a natural phenomenon of the developing system rather than a reaction to trauma (Bates, 1992). This hypothesis is supported by the results of recent evoked response potential (ERP) studies with normal children (Mills et al., 1994; Neville, 1995; Mills and
Neville, 1997), which indicate that lateralization of language increases during childhood. Although there are clear structural hemispheric diVerences in language areas even in the fetus (Chi et al., 1977), the results of recent ERP studies suggest that functional lateralization of language continues to develop over the Wrst few months or years of life.
Language consists of six major components: the sound system (phonology), the content system (semantics), the rules of word formulation (morphology), the rules of sentence formation (syntax), the suprasegmental aspects of language (prosody), and the social use of language (pragmatics). The phonologic dimension reXects the ability to appreciate the distinct nature of individual speech sounds and sound patterns. Semantics refers to our knowledge of speciWc word meanings and understanding of relationships between words. Morphology describes the rules for combining morphemes (the smallest units of meaning) to form words and phrases. It also includes the use of inXection (as in the plural form) and derivation (creating new words by adding preWxes and suYxes). Syntax refers to the rules for joining words to make sentences. Prosody (also called the suprasegmental aspect of language) comprises vocal inXection, stress intonation, pausing, and other variables that contribute to the rhythmic aspects of language. Finally, pragmatics takes into account how variations in the intent of the speaker (e.g., goal or purpose) and the social milieu (e.g., speaker±listener relationship or situation) inXuence the linguistic output. Pragmatics includes the understanding and use of gestures as well as the social context in which language is used.
In the adult brain, these language dimensions are mediated by distinct neural systems. For example, after brain injury, understanding of words can be impaired while phonologic and syntactic processing remains intact (Allport and Funnell, 1981). Even diVerent types of semantic knowledge can be selectively impaired by brain lesions (e.g., Warrington and Shallice, 1984). Recent neuroimaging studies have provided detailed information about the representation of speciWc linguistic processes in normal adults (e.g., Martin et al., 1995, 1996; Damasio et al., 1996) and diVerences in the functional organization of language between males and females (Shaywitz et al., 1995). Although comparable developmental studies are limited, the available evidence suggests that diVerent dimensions of language can be dissociated in normal children (Neville, 1995), as well as in children with developmental language disorders (Korkman and Häkkinen-Rihu, 1994; Rapin and Allen, 1988). It is not known, however, how the neural systems that support speciWc language functions organize
Cognitive development from a neuropsychologic perspective |
147 |
|
|
|
|
and become more reWned during the course of normal development.
There are several principles of language development that are well documented. First, as noted above, language acquisition progresses at a remarkable pace during the Wrst 3 years of life and coincides with rapid changes in neural development. Second, there are critical periods in the acquisition of language. Support for the concept of critical periods comes from diVerent lines of investigation, including studies of children raised in grossly deprived environments (Curtiss, 1977), deaf isolates (Curtiss, 1989), and bilingual individuals (Johnson and Newport, 1989). Third, the emergence of language is an active process. Children generate their linguistic output and do not merely repeat what they have heard. Fourth, the course of language development is remarkably similar across individuals and cultures. Most children show a comparable progression in their acquisition of early language milestones.
The development of language begins well before the child speaks his Wrst words (see Fig. 8.4 for a description of the early language milestones). During the third trimester of pregnancy, for example, the fetus reacts to the maternal voice (de Casper et al., 1994). Shortly after birth, infants show a consistent response to sounds (Locke, 1996, 1997) and to variations in vocal emotion (Caron et al., 1988). Auditory orienting responses appear several months later, at approximately 4 months of age, when the infant orients to the source of sound, responds to the human voice, and learns to recognize the caregiver's vocal characteristics. These early behaviors provide the rudimentary basis for language and social development and are vital to the acquisition of linguistic proWciency. Subsequent receptive milestones have not been clearly mapped, but it seems that at all levels of development, children are able to comprehend more language than they are able to speak.
Expressive milestones have been studied extensively. These milestones show a clear pattern of development (for reviews see Locke, 1997; Owens, 1992), beginning with cooing and the indiscriminate use of certain sounds (dada) during the Wrst 4 months of life. These sounds give the infants a set of utterances that they can use in certain contexts and provides them with the opportunity to participate in social interactions. Babbling takes on speech-like characteristics and often overlaps with the emergence of the Wrst few words between 10 and 12 months of age. Already by the end of the Wrst year, babies use many single words to comment on their actions, the actions of others, and the environment around them. Most of the children's Wrst words are labels for objects (e.g., dada, mama, and cookie), action words (e.g., eat, come, and go), and social
terms (e.g., bye and hi). Language explodes at around 2 years of age (Plunkett, 1993) when children also begin to show symbolic capacities in play and elaborate social behaviors. The most evident explosion is seen in the child's vocabulary. For example, at 2 years of age, most children have a vocabulary of at least 50 words and combine words into two-word phrases or utterances (e.g., want cookie). By 3 years of age, the child's vocabulary has increased to approximately 250 words, and he or she is using threeword phrases. These short phrases are devoid of articles, prepositions, and auxiliary verbs and are referred to as telegraphic speech because nonessential words are omitted. In English, where word order conveys meaning, 2- and 3- year-old children demonstrate the subject±action±object order as their Wrst rule. By the end of the year, children begin to add grammatical inXections to their words (e.g., ª-edº, resulting in ªcomedº for ªcameº) and gradually expand the length of their sentences. They gradually begin to add elements to their speech, such as pronouns, prepositions, and grammatical morphemes, and elaborate the basic structure of the sentence to reXect more complex knowledge. Children possess perhaps as many as 400 words before they begin to apply the rules of linguistic morphology (Bates et al., 1994). By 4 years of age, the rapid development of language skills that characterize the Wrst years of life is no longer seen.
By the time children enter school, at age 6 in the USA, they can use language to initiate conversation, maintain a topic, express feelings and emotions, and interact socially. They continue to reWne their use of morphologic and syntactic rules and begin to view situations from the perspective of other people. By 8 years of age, children produce all phonemes correctly and exhibit the main characteristics of the adult grammar. Their language acquisition is not complete. In fact, language development continues through the school-age years and into adulthood (Johnson, 1996) in interaction with developing conceptual abilities, experience, and education. However, from this time on, additions to the linguistic system take the form of subtle reWnements in the way language is used. The child becomes increasingly skilled at adapting his language to the speciWc situation and partner and shows an increased understanding of metaphoric language (e.g., feeling blue).
The association between language and cognitive development has been studied extensively since the early 1970s. Brown (1973), for example, noted strong parallels between the child's acquisition of language and Piaget's model of cognitive development. SpeciWcally, Piaget emphasized the salience of objects and actions from the child's point of view, a salience that is supported by the Wrst utterances. These utterances typically describe objects and actions

Receptive |
0 months |
3 months |
6 months |
9 months |
1 year |
|
|
2 years |
3 years |
|
|||||||||||||||||||||||||||||||||||||||||||||
language |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Responds |
Discriminates |
|
Responds |
|
|
|
|
Knows body parts, |
|
|
|
Masters many |
|
|
|
Masters spatial |
|
||||||||||||||||||||||||||||||||||||
|
|
to sounds |
speech sounds |
differentially |
|
|
|
|
household objects |
|
|
|
single phonemes |
|
|
|
and temporal |
|
|||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
to sounds |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
and consonant blends |
|
|
|
words |
|
||||||||||||||||||||||
|
|
Discriminates |
Turns head in |
|
|
|
|
|
|
|
Understands |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
|
intonational |
direction of sound |
|
|
|
|
|
|
phonemic categories |
|
|
|
Recognizes semantic |
|
|
|
Masters other |
|
||||||||||||||||||||||||||||||||||||
|
|
patterns |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
classes |
|
|
|
consonant blends |
|
|||||||||||||||||||||||
Expressive |
0 months |
3 months |
6 months |
9 months |
|
1 year |
|
|
|
|
|
|
|
|
|
|
|
2 years |
3 years |
|||||||||||||||||||||||||||||||||||
language |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Undifferentiated |
Sounds |
|
Produces sounds |
Emergence of |
|
|
Uses about |
|
|
|
|
|
|
Emergence of |
|
|
Auxillary verbs, |
|||||||||||||||||||||||||||||||||||
|
|
crying and |
approximate |
|
with varying |
|
|
single words |
|
|
100 words |
|
|
|
|
|
|
morphemes |
|
|
irregular past |
|||||||||||||||||||||||||||||||||
|
|
cooing |
speech |
intonation patterns |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
and possessive |
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Uses two-word |
|
|
|
|
|
|
Uses telegraphic |
||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
utterances |
|
|
|
|
|
|
sentences |
|
|
Uses four-word |
|||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
utterances |
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Asks simple |
|
|
|
|
|
|
Asks what and |
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
questions |
|
|
|
|
|
|
where questions |
||||||||||||||||||||||||||||
Communicative 0 months |
3 months |
6 months |
9 months |
|
1 year |
|
|
|
|
|
|
|
|
|
|
|
2 years |
3 years |
|||||||||||||||||||||||||||||||||||
language |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Maintains |
Nonverbal |
|
Uses variations in |
Indicates |
|
|
Plays to elicit |
|
Uses household |
|
|
|
Engages in turn |
|||||||||||||||||||||||||||||||||||||||
|
eye contact |
turn taking |
|
pitch and intonation |
desires |
|
|
a specific |
|
objects in play |
|
|
|
taking |
|||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
response |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Communicates about |
|
|
|
Has imaginative |
|||||||||||||||||||||
|
|
|
|
Smiles with |
|
Imitation develops |
Uses simple |
|
|
|
|
|
|
|
|
|
|
|
|
here and now |
|
|
|
play |
|||||||||||||||||||||||||||||
|
|
|
|
parents |
|
|
|
|
|
gestures |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
Aware of own vocalizations
Fig. 8.4. Early language milestones.
Cognitive development from a neuropsychologic perspective |
149 |
|
|
|
|
with which the child is familiar (e.g., want milk). However, contrary to Piaget's prediction, more advanced language milestones do not seem to follow speciWc cognitive competencies but rather emerge at the same time. Children use words to learn new concepts as well as to express ideas that they understand. Therefore, it seems that children in the sensorimotor period (birth to 2 years of age) draw upon concepts to master language, whereas children in more advanced stages use language to learn new concepts and to communicate what they know.
Visuospatial skills
This section provides a concise review of visual processing abilities. The organization of visual areas has been mapped extensively in primates and human adults (for reviews see Ungerleider and Mishkin, 1982; Ungerleider and Haxby, 1994; Courtney and Ungerleider, 1997). These studies reveal that visual cortical areas are organized into two functionally specialized pathways for object and spatial vision. The dorsal stream or ªwhereº system provides information regarding the location of objects in space and the spatial relationship among objects. This pathway originates in area V1 and projects to the middle temporal area and to additional areas in the superior temporal and parietal cortex. The ventral stream or ªwhatº system, in turn, recognizes the identity or nature of objects (e.g., color, pattern, and shape) regardless of spatial location and is mediated by the inferior temporal cortex. Regions in the occipitotemporal and occipitoparietal pathways are organized hierarchically. In other words, early processing areas within both pathways are dedicated to the detection of speciWc features (e.g., angles, edges), while areas involved in later processing subserve increasingly integrated and general perceptions (e.g., objects). This loss in speciWcity is accompanied by increasing selectivity for complex stimuli. For example, within the occipitotemporal pathway, neurons in V1 respond to contours and edges, whereas neurons in the inferior temporal cortex respond to global features such as faces.
An anomaly in the dorsal stream, particularly in the magnocellular visual system, which subserves transient processing may account for the diYculties in temporal processing hypothesized in developmental dyslexia (Eden et al., 1996). Disordered perceptions of spatial relationships (e.g., Wnger agnosia, visuospatial neglect, constructional apraxia) have been attributed to anomalies in the occipitotemporal pathway (Newcombe and RatcliV, 1989). In addition, anomalies in areas of the ventral occipitotemporal cortex (e.g., collateral sulcus and lingual gyrus) can cause color-blindness (Damasio et al., 1980) and problems in visual discrimination. SpeciWc areas within the central
occipitotemporal cortex are sensitive to the perception of faces (Desimone, 1991; Clark et al., 1996) and have been implicated in disorders of facial recognition (Damasio et al., 1982, 1990).
The occipitotemporal and occipitoparietal pathways show diVerent developmental trajectories in nonhuman primates. Although both pathways are immature at birth, the occipitoparietal system matures at 2 months of age, approximately 1 month earlier than the occipitotemporal pathway (Distler et al., 1996). It is not known if the same Wnding applies to humans. The evidence suggests that these pathways are not fully mature at birth in human infants (for reviews see Cohen and Salapatek, 1975; Aslin et al., 1981). Neonates are sensitive to brightness and can discriminate between gross visual features but have diYculty identifying patterns and forms. Similarly, they can only detect gross movements in the visual Weld. At approximately 2 to 3 months of age, infants can track objects and discriminate simple forms. Between 2 and 12 months of age, the visual system matures rapidly and the infant becomes capable of making increasingly complex visual discriminations. The nature of their discriminations suggests that, in general, infants are attracted to high-contrast patterns and moving objects.
The infant's preference for movement and high-con- trast features likely makes faces among the most interesting of visual stimuli. The study of facial recognition has received signiWcant attention in recent years, not only because of the contribution of facial perception to the understanding of other perceptual abilities but also because of its role in the development of social competence. Studies of facial recognition have shown that, during the Wrst weeks of life, infants recognize their mother's face. Facial recognition shows rapid qualitative changes during the Wrst 7 months of life. In the early stages of development, infants rely on the external conWguration of the mother's face (e.g., outer contour of the head and hairline) (Bushnell, 1982; Bushnell et al., 1989; Pascalis et al., 1995). Facial recognition becomes more reWned in the Wrst few months and, by the age of 5 months, infants are able to recognize the mother's face from its internal details (de Schonen and Mathivet, 1990). By 7 months of age, babies can recognize faces even when shown in diVerent poses or from diVerent vantage points (Fagan, 1976). Accuracy of facial recognition increases steadily during the Wrst 5 years of life. By age 6, children can recognize faces as well as do adults but continue to have diYculty interpreting some facial expressions (Kolb et al., 1992). In fact, perception of facial expression continues to improve during childhood up until about 14 years of age. Kolb et al. (1992) argue that the neural

150 D. M. Pascualvaca and G. Morote
substrates of facial recognition diVer from those underlying the perception of facial expression, and that the latter ability reXects the integrity of the frontal lobes.
Motor and visuomotor
The development of motor functions is a remarkable process. In a matter of months, the crudely coordinated movements of the neonate are transformed into elegant and precise movements. Posture improves, coordination becomes more precise, and the infant begins to reach for and grasp objects. A review of motor development is beyond the scope of this chapter and only a cursory glance is presented here. Very brieXy, motor development proceeds from general to speciWc. General movements are followed by reWned actions and by subsequent integration of these movements. Motor development also progresses in cephalo±caudal and proximo±distal directions, with skilled manual movements appearing relatively late in development. Manipulatory dexterity shows greater precision with age. Skilled manual activities also become more automatic during childhood. For instance, increased speed in Wne motor tasks such as repetitive and successive Wnger movements represent an important aspect of developmental progression (Denckla, 1973). Similarly, there is a marked increase in grip strength between the ages of 6 and 11 years. This increase is more dramatically seen in boys (3.6-fold) than in girls (2.6-fold) (Espenschade and Eckert, 1980).
In addition to improved motor coordination, strength and speed, there is a gradual decrease in extraneous movements during childhood. Extraneous motor movements are not unusual before the age of 7 when the child performs certain activities, but they become increasingly rare as the child develops. Similarly, motor impersistence, normal in younger children, constitutes an atypical feature in adolescents (Benton et al., 1983). Motor impersistence, the inability to maintain an action, is universal among 5-year- olds, decreases signiWcantly between the ages of 5 and 7 and is relatively infrequent by age 10 (Benton et al., 1983). More complex motor behaviors such as inhibition of motor reactions are associated with executive functions and may not reach maturity until late adolescence.
Although visuospatial and visuomotor development have been covered separately in this chapter, integrative processes are equally important. A child's perceptual and motor functions can be well developed, but integration of visual perception with movements may continue to be problematic. Copying forms, for example, requires adequate interpretation of visual stimuli, execution of Wne motor responses, and the integration of these processes.
Copying complex designs also engages other cognitive processes, including planning and organization of response patterns as well as shifting of attention between what is seen and what is being reproduced.
Summary and future directions
In this chapter, cognitive development has been discussed as comprising distinct functional systems that undergo a process of reWnement with age. This process varies for the diVerent cognitive functions and coincides with the maturation of intricate interactions among the various functional systems. For example, we have seen how the emergence of speciWc language competencies impact upon the reWnement of executive functions and how these, in turn, aVect the modulation of attentional capacities.
To date, the assessment of cognitive functions has been limited by the array of clinical tests available. These procedures do not tap isolated functions free from the interactive eVects of other cognitive systems, and they may require skills that are beyond the child's developmental capabilities. Neuroimaging can overcome these limitations by providing an opportunity to localize the functional circuits that subserve performance on a task. By doing so, we can gain a better understanding of speciWc brain± behavior relationships in young children that have so far eluded us. Broadening the knowledge base of normal development from a neural perspective can also facilitate an understanding of aberrant development and consequent cognitive disorders.
Recent studies have already challenged traditional concepts of developmental trajectories. In some instances, they have shown that certain cognitive functions are present earlier than previously purported. For example, studies of infants (Casey and Richards, 1991) have shown that a rudimentary ability to sustain attention may be present far earlier than traditionally thought on the basis of clinical measures. At the same time, other studies have shown that cognitive functions may continue to undergo reWnement and changes well into the adult years. The ceilings for maturation of various cognitive functions have been identiWed to be around the ages of 10 to 12 years, but this may simply reXect the limitations of neuropsychologic tests, which may not capture reWnement at more advanced ages. Indeed, neuroimaging studies have shown that the brain continues to undergo striking reorganization even in adulthood (Courtney and Ungerleider, 1997). Therefore, functional neuroimaging holds great promise for mapping the brain maturational changes underlying neuropsychologic development in children and adolescents.

Cognitive development from a neuropsychologic perspective |
151 |
|
|
|
|
iReferencesi
Allport, A. and Funnell, E. (1981). Components of the mental lexicon. Philos. Trans. R. Soc. Lond., 295, 397±410.
Alvarez, P., Zola-Morgan, S. and Squire, L. R. (1994). The animal model of human amnesia: long-term memory impaired and short-term memory intact. Proc. Natl. Acad. Sci. USA, 91, 5637±41.
Aslin, R. N., Alberts, J. R. and Petersen, M. R. (eds.) (1981). The Development of Perception: Psychophysiological Perspectives. New York: Academic Press.
Bachevalier, J. (1990). Ontogenetic development of habit and memory formation in primates. In Development and Neural Basis of Higher Cognitive Functions, ed. A. Diamond, pp. 457±84. New York: New York Academy of Sciences Press.
Bachevalier, J. and Mishkin, M. (1984). An early and a late developing system for learning and retention in infant monkeys. Behav. Neurosci., 98, 770±8.
Baddeley, A. D. (1992). Working memory. Science, 255, 556±9. Baillargeon, R. (1995). Physical reasoning in infancy. In The
Cognitive Neurosciences, ed. M. S. Gazzaniga, pp. 181±204. Cambridge, MA: MIT Press.
Barkley, R. A., Grodzinsky, G. M. and DuPaul, G. J. (1992). Frontal lobe functions in attention deWcit disorder with and without hyperactivity: a review and research report. J. Abnormal Child Psychol., 20, 163±88.
Bates, E. A. (1992). Language development. Curr. Opin. Neurobiol.,
2, 180±5.
Bates, E. A., Dale, P. S. and Thal, D. (1994). Individual diVerences and their implications for theories of language development. In
Handbook of Child Language, eds. P. Fletcher and B. MacWhinney, pp. 96±151. Oxford: Blackwell.
Bauer, P. J. (1997). Development of memory in early childhood. In
The Development of Memory in Childhood, ed. N. Cowan, pp. 83±111. Hove, UK: Psychology Press.
Becker, M. G., Isaac, W. and Hynd, G. (1987). Neuropsychological development of non-verbal behaviors attributed to `frontal lobe' functioning. Dev. Neuropsychol., 3, 275±98.
Benton, A. L., de S. Hamsher, K.,Varney, N. R. and Spreen, O. (1983).
Contribution to Neuropsychological Assessment. New York: Oxford University Press.
Brown, R. (1973). A First Language: The early stages. Cambridge, MA: Harvard University Press.
Bushnell, I. W. (1982). Discrimination of faces by young infants. J. Exp. Child Psychol., 33, 298±308.
Bushnell, I. W., Sai., F. and Mullin, J. T. (1989). Neonatal recognition of the mother's face. Br. J. Devel. Psychol., 7, 3±15.
Caron, A. J., Caron, R. F. and MacLean, D. J. (1988). Infant discrimination of naturalistic emotional expressions: the role of face and voice. Child Dev., 59, 604±16.
Casey, B. J. and Richards, J. E. (1991). A refractory period for the heart rate response in infant visual attention. Dev. Psychobiol.,
24, 327±40.
Casey, B. J., Cohen, J. D., Jezzard, P., et al. (1995). Activation of prefrontal cortex in children during a nonspatial working memory task with functional MRI. Neuroimage, 2, 221±9.
Chelune, G. J. and Baer, R. L. (1986). Developmental norms for the Wisconsin Card Sorting Test. J. Clin. Exp. Neuropsychol., 8, 219±28.
Chi, J. G., Dooling, E. C. and Gilles, F. H. (1977). Left±right asymmetries of the temporal speech areas in the human fetus. Arch. Neurol., 34, 346±8.
Clark, V. P., Keil, K., Maisog, J. M., Courtney, S., Ungerleider, L. G. and Haxby, J. V. (1996). Functional magnetic resonance imaging of human visual cortex during face matching: comparison with positron emission tomography. Neuroimage, 4, 1±15.
Cohen, L. B. and Salapatek, P. (eds.) (1975). Infant Perception: From Sensation to Cognition. New York: Academic Press.
Cohen, R. M., Semple, W. E., Gross, M., Holcomb, H. H., Dowling, M. S. and Nordahl, T. E. (1988). Functional localization of sustained attention: comparison to sensory stimulation in the absence of instruction. Neuropsychiatr. Neuropsychol. Behav. Neurol., 1, 3±20.
Cooley, E. L. and Morris, R. D. (1990). Attention in children: a neuropsychologically based model for assessment. Dev. Neuropsychol., 6, 239±74.
Courchesne, E., AkshoomoV, N. A., Townsend, J. and Saitoh, O. (1995). A model system for the study of attention and the cerebellum: infantile autism. Electroencephalogr. Clin. Neurophysiol., 44
(Suppl.), 315±25.
Courtney, S. M. and Ungerleider, L. G. (1997).What fMRI has taught us about human vision. Curr. Opin. Neurobiol., 7, 554±61.
Courtney, S. M., Ungerleider, L. G., Keil, K. and Haxby, J. V. (1996). Object and spatial working memory activate separate neural systems in human cortex. Cerebr. Cortex, 6, 39±49.
Courtney, S. M., Petit, L., Maisog, J. M., Ungerleider, L. G. and Haxby, J. V. (1998). An area specialized for spatial working memory in human frontal cortex. Science, 279, 1347±51.
Cowan, N. (1997). The development of working memory. In The Development of Memory in Childhood, ed. N. Cowan, pp. 163±9. Hove, UK: Psychology Press.
Curtiss, S. (1977). Genie: A Psychological Study of a Modern Day ªWild Childº. New York: Academic Press.
Curtiss, S. (1989). The independence and task-speciWcity of language. In Interaction in Human Development, ed. A. Bornstein and J. Bruner, pp. 105±37. Hillsdale, NJ: Erlbaum.
Damasio, A. R., Yamada, T., Damasio, H., Corbett, J. and McKee, J. (1980). Central achromatopsia: Behavioral, anatomical and physiologic aspects. Neurology, 30, 1064±71.
Damasio, A. R., Damasio, H. and van Hoesen, G. W. (1982). Prosopagnosia: anatomic basis and behavioral mechanisms.
Neurology, 32, 331±41.
Damasio, A. R., Tranel, D. and Damasio, H. (1990). Face agnosia and the neural substrates of memory. Annu. Rev. Neurosci., 13, 89±109.
Damasio, H., Grabowski, T. J., Tranel, D., Hichwa, R. D. and Damasio, A. R. (1996). A neural basis for lexical retrieval. Nature,
380, 499±505.
Davies, D. R., Jones, D. M. and Taylor, A. (1984). Selectiveand sus- tained-attention tasks: individual and group diVerences. In
Varieties of Attention, eds. R. Parasuraman and D. R. Davies, pp. 395±446. Orlando, FL: Academic Press.
de Casper, A., Lecanuet, J. P., Busnel, M. C., Granier-Defere, C. and

152 D. M. Pascualvaca and G. Morote
Maugeais, R. (1994). Fetal reactions to recurrent maternal speech. Infant Behav. Dev., 17, 159±64.
D'Esposito, M., Detre, J. A., Alsop, D. C., Shin, R. K., Atlas, S. and Grossman, M. (1995). The neural basis of the central executive system of working memory. Nature, 378, 279±81.
Dempster, F. N. (1981). Memory span: sources of individual and developmental diVerences. Psychol. Bull., 89, 63±100.
Denckla, M. D. (1973). Development of speed in normal children.
Dev. Med. and Child Neurol., 15, 635±45.
de Schonen, S. and Mathivet, E. (1990). Hemispheric asymmetry in a face discrimination task in infants. Child Dev., 61, 1192±1205.
Desimone, R. (1991). Face-selective cells in the temporal cortex of monkeys. J. Cogn. Neurosci., 3, 1±8.
Deutsch, G., Papinicolaou, A. C., Bourbon, W. T. and Eisenberg, H. M. (1987). Cerebral blood Xow evidence of right frontal activation in attention demanding tasks. Int. J. Neurosci., 36, 23±8.
Diamond, A. (1990). Rate of maturation of the hippocampus and the developmental progression of children performance on the delayed non-matching to sample and visual paired comparison tasks. Anna. N.Y. Acad. Sci., 608, 394±426.
Diamond, A. (1995). Evidence of robust recognition memory early in life even when assessed by reaching behavior. J. Exp. Child Psychol., 59, 419±74.
Diamond, A. and Goldman-Rakic, P. S. (1989). Comparison of human infants and rhesus monkeys on Piaget's AB task: evidence for dependence on dorsolateral prefrontal cortex. Exp. Brain Res., 74, 24±40.
Distler, C., Bachevalier, J., Kennedy, C., Mishkin, M. and Ungerleider, L. G. (1996). Functional development of the corticocortical pathway for motion analysis in the macaque monkey: a
14C-2-deoxyglucose study. Cereb. Cortex, 6, 184±95.
Eden, G. F., van Meter, J. W., Rumsey, J. M., Maisog, J. M., Woods, R. P. and ZeYro, T. A. (1996). Abnormal processing of visual motion in dyslexia revealed by functional brain imaging. Nature, 382, 66±9.
Espenschade, A. S. and Eckert, H. M. (1980). Motor Development, 2nd edn. Columbus, OH: Charles E. Merrill.
Ewing-Cobbs, L., Prasad, M., Fletcher, J. M., Levin, H. S., Miner, M. E. and Eisenberg, H. M. (1998). Attention after pediatric traumatic brain injury: a multidimensional assessment. Child Neuropsychol., 4, 35±48.
Fagan, J. F. (1976). Infants' recognition of invariant features of faces. Child Dev., 47, 627±38.
Feldman, H. M., Holland, A. L., Kemp, S. S. and Janosky, J. E. (1992). Language development after unilateral brain injury. Brain Lang.,
42, 89±102.
Fernandez, G., Weyerts, H., Schrader-Bölsche, M. et al. (1998). Successful verbal encoding into episodic memory engages the posterior hippocampus: a parametrically analyzed functional magnetic resonance imaging study. J. Neurosci., 18, 1841±7.
Fiducia, D. and O'Leary, D. S. (1990). Development of a behavior attributed to the frontal lobes and the relationship to other cognitive functions. Dev. Neuropsychol., 6, 85±94.
Funahashi, S., Bruce, C. J. and Goldman-Rakic, P. S. (1993). Dorsolateral prefrontal lesions and oculomotor delayed-
response performance. Evidence for mnemonic ªscotomasº. J. Neurosci., 13, 1479±97.
Fuster, J. M. (1985). The preferred cortex: mediator of cross-tem- poral contingencies. Hum. Neurobiol., 4, 169±79.
Gathercole, S. E. (1998). The development of memory. J. Child Psychol. Psychiatr. Allied Disciplines, 39, 3±27.
Goldman-Rakic, P. S. (1987). Development of cortical circuitry and cognitive functions. Child Dev., 58, 601±22.
Goldman-Rakic, P. S. (1988). Topography of cognition: parallel distributed networks in primate association cortex. Annu. Rev. Neurosci., 11, 137±56.
Greenberg, L. M. and Waldman, I. D. (1993). Developmental normative data on the test of variables of attention (T.O.V.A.). J. Child Psychol. Psychiatr. Allied Disciplines, 34, 1019±30.
Hershey, T., Craft, S., Glauser, T. A. and Hale. (1998). Short-term and long-term memory in early temporal lobe dysfunction.
Neuropsychology, 12, 52±64.
Hudspeth, W. J. and Pribram, K. H. (1992). Psychophysiological indices of cerebral maturation. Int. J. Psychophysiol., 12, 19±29.
Hulme, C., Muir, C., Thomson, N. and Lawrence, A. (1984). Speech rate and the development of short-term memory span. J. Exp. Child Psychol., 38, 241±53.
Ito, Y., Teicher, M. H., Glod, C. A., Harper, D., Magnus, E. and Gelbard, H. A. (1993). Increased prevalence of electrophysiological abnormalities in children with psychological, physical and sexual abuse. J. Neuropsychiatr. Clin. Neurosci., 5, 401±8.
Jetter, W., Poser, U., Freeman, R. B. and Markowitsch, J. H. (1986). A verbal long term memory deWcit in frontal lobe damaged patients. Cortex, 22, 229±42.
Johnson, B. A. (1996). Language Disorders in Children. Albany:
Delmar.
Johnson, J. and Newport, E. (1989). Critical periods eVects in second language learning. The inXuence of maturational state on the acquisition of English as a second language. Cognit. Psychol., 21, 60±99.
Johnson, M. (1995). The development of visual attention: a cognitive neuroscience perspective. In The Cognitive Neurosciences, ed. M. S. Gazzaniga, pp. 737±47. Cambridge, MA: MIT Press.
Jonides, J., Smith, E. E., Koeppe, R. A., Awh, E., Minoshima, S. and Mintun, M. A. (1993). Spatial working memory in humans as revealed by PET. Nature, 363, 623±5.
Kohn, B. and Dennis, M. (1971). Patterns of hemispheric specialization after hemidecortication for infantile hemiplegia. In
Hemispheric Disconnection and Cerebral Function, ed. M. Kinsbourne and W. L. Smith, pp. 34±47. SpringWeld, IL: Thomas.
Kolb, B., Wilson, B. and Taylor, I. (1992). Developmental changes in the recognition and comprehension of facial expression: implications for frontal lobe function. Brain Cognit., 20, 74±84.
Korkman, M. and Häkkinen-Rihu, P. (1994). A new classiWcation of developmental language disorders (DLD). Brain Lang., 47, 96±116.
Kreutzer, M. A., Leonard, C. and Flavell, J. H. (1975). An interview study of children's knowledge about memory. Monogr. Soc. Res. Child Dev., 40, 1±60.
Lane, D. M. and Pearson, D. A. (1982). The development of selective attention. Merrill-Palmer Quart., 28, 317±27.
Cognitive development from a neuropsychologic perspective |
153 |
|
|
|
|
Levine, D. N., Warach, J. and Farah, M. J. (1985). Two visual systems in mental imagery: dissociation of ªwhatº and ªwhereº in imagery disorders due to bilateral posterior cerebral lesions.
Neurology, 35, 1010±18.
Locke, J. L. (1996). Why do infants begin to talk? Language as an unintended consequence. J. Child Lang., 23, 251±68.
Locke, J. L. (1997). Towards a biological science of language development. In The Development of Language, ed. M. Barrett, pp. 373±95. London: UCL Press.
Loss, N., Yeates, K. O. and Enrile, B. G. (1998). Attention in children with myelomeningocele. Child Neuropsychol., 4, 7±20.
Luria, A. R. (1959). The directive function of speech in development and dissolution. Word, 15, 341±52.
Luria, A. R. (1973). The Working Brain. New York: Basic Books. Macoby, E. E. and Jacklin, C. N. (1974). The Psychology of Sex
DiVerences. Stanford: Stanford University Press.
Malkova, L., Mishkin, M. and Bachevalier, J. (1995). Long-term eVects of selective neonatal temporal lobe lesions on learning and memory in monkeys. Behav. Neurosci., 109, 212±26.
Martin, A., Haxby, J. V., Lalonde, F. M., Wiggs, C. L. and Ungerleider, L. G. (1995). Discrete cortical regions associated with knowledge of color and knowledge of action. Science, 270, 102±5.
Martin, A., Wiggs, C. L., Ungerleider, L. G. and Haxby, J. V. (1996). Neural correlates of category-speciWc knowledge. Nature, 379, 649±52.
McKay, K. E., Halperin, J. M., Schwartz, S. T. and Sharma, V. (1994). Developmental analysis of three aspects of information processing: sustained attention, selective attention, and response organization. Dev. Neuropsychol., 10, 121±32.
Mills, C., Morgan, M. J., Milne, A. B. and Morris, D. M. (1996). Developmental and individual diVerences in visual memory span. Curr. Psychol., 15, 53±67.
Mills, D. L. and Neville, H. J. (1997). Electrophysiological studies of language and language impairment. Semin. Pediatr. Neurol., 4, 125±34.
Mills, D. M., CoVey, S. A. and Neville, H. J. (1994). Language acquisition and cerebral specialization in 20-month-old infants. J. Cognit. Neurosci., 5, 326±42.
Milner, B., Branch, C. and Rasmussen, R. (1964). Observations on cerebral dominance: In Disorders of Language, ed. A. V. S. de Reuck and M. O'Connor, pp. 200±14. London: Churchill.
Mirsky, A. F. (1987). Behavioral and psychophysiological markers of disordered attention. Environ. Health Perspect., 74, 191±9.
Mirsky, A. F., Anthony, B. J., Duncan, C. C., Ahearn, M. B. and Kellam, S. G. (1991). Analysis of the elements of attention: a neuropsychological approach. Neuropsychol. Rev., 2, 109±45.
Nelson, C. A. (1997). The neurobiological basis of early memory development. In The Development of Memory in Childhood, ed. N. Cowan, pp. 41±82. Hove, UK: Psychology Press.
Neville, H. J. (1995). Developmental speciWcity in neurocognitive development in humans. In The Cognitive Neurosciences, ed. M. S. Gazzaniga, pp. 219±31. Cambridge, MA: MIT Press.
Newcombe, R. and RatcliV, G. (1989). Disorders of visuospatial analysis. In Handbook of Neuropsychology, eds. F. Boller and F. Grafman, pp. 333±56. Amsterdam: Elsevier.
Overman, W. H., Bachevalier, J., Turner, M. and Peuster, A. (1992). Object recognition versus object discrimination: comparison between human infants and infant monkeys. Behav. Neurosci.,
106, 15±29.
Overman, W. H., Bachevalier, J., Sewell, F. and Drew, J. (1993). A comparison of children's performance on two recognition memory tasks: delayed nonmatch-to-sample vs. visual pairedcomparison. Dev. Psychobiol., 26, 345±57.
Owens, R. E. (1992). Language Development: An Introduction.
Columbus: Merrill.
Pardo, J. V., Fox, P. T. and Raichle, M. E. (1991). Localization of a human system for sustained attention by positron emission tomography. Nature, 349, 61±4.
Pascalis, O., de Schonen, S., Morton, J., Deruelle, C. and FabreGrenet, M. (1995). Mother's face recognition in neonates: a replication and an extension. Infant Behav. Dev., 18, 79±85.
Pascual-Leone, J. A. (1970). A mathematical model for the transition rule in Piaget's developmental stages. Acta Psychol., 32, 301±45.
Pascualvaca, D. M., Anthony, B. J., Arnold, L. E. et al. (1997). Attention performance in an epidemiological sample of urban children: the role of sex and verbal intelligence. Child Neuropsychol., 3, 13±27.
Pascualvaca, D. M., Fantie, B. D., Papageorgiou, M. and Mirsky, A. F. (1999). Attention capacities in children with autism: is there a general deWcit in shifting focus? J. Autism Dev. Dis. Disord., 28, 467±78.
Passler, M., Isaac, W. and Hynd, G. W. (1985). Neuropsychological development of behavior attributed to frontal lobe functioning in children. Dev. Neuropsychol., 1, 349±70.
Pea, R. (1982). What is planning development the development of? In Children's Planning Strategies, eds. D. Forbes and M. T. Greenberg, pp. 5±28. San Francisco, CA: Jossey-Bass.
Piaget, J. (1952). The Origins of Intelligence in Children. New York: International University Press.
Piaget, J. (1954). The Construction of Reality in the Child. New York: Basic Books.
Piaget, J. (1976). The Grasp of Consciousness. Cambridge: Harvard
University Press.
Plunkett, K. (1993). Lexical segmentation and vocabulary growth in early language acquisition. J. Child Lang., 20, 43±60.
Posner, M. I. (1993). Seeing the mind. Science, 262, 673±74. Posner, M. I. and Petersen, S. E. (1990). The attention system of the
human brain. Annu. Rev. Neurosci., 13, 25±42.
Posner, M. I., Petersen, S. E., Fox, P. T. and Raichle, M. E. (1988). Localization of cognitive operations in the human brain. Science,
240, 1627±31.
Pressley, M. and Schneider, W. (1997). Introduction to Memory Development During Childhood and Adolescence. New Jersey: Lawrence Erlbaum.
Pribram, K. H. and McGuinness, D. (1975). Arousal, activation, and eVort in the control of attention. Psychol. Rev., 82, 116±49.
Rabinowicz, T. (1976). Morphological features of the developing brain. In Brain dysfunction in infantile febrile convulsions, eds. M. A. B. Brazier and A. Coceani, pp. 1±23. New York: Raven Press.