

Радиобиология с основами радиоэкологии
.pdf6. Радиационно-химические и радиационно-биохимические процессы... |
191 |
ей. Разрыв полимерных цепей является основной реакцией для целлюлозы, облученной быстрыми электронами. Кроме прямой деполимеризации, например, в результате атаки кислорода гликозидных связей (как это происходит в крахмале), возникают повреждения мономерных единиц, которые лабилизируют полимерную цепь, и происходит разрыв цепи, проявляющийся по прошествии длительного времени после облучения. Процесс последействия может выражаться в снижении вязкости облученных растворов полимеров в течение многих дней после облучения. Таким образом, изменения физико-химических свойств высокомолекулярных углеводов связаны главным образом с деполимеризацией молекул, разрывом внешних эфирных мостиков между мономерами.
6.6.3. Действие ионизирующей радиации на белково-липидные мембраны
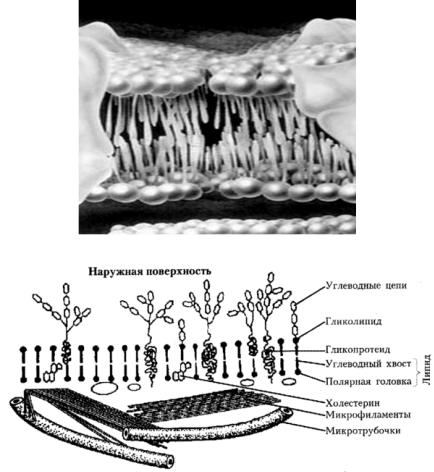
Липиды – вещества с молекулярной массой порядка 750– 1000 дальтон – являются производными трехатомного спирта (глицерина). Основу клеточной мембраны составляет двойной слой полярных липидов (фосфолипидов). Они могут служить мишенью при действии ионизирующего излучения, так как повреждение их даже в небольших количествах может изменить проницаемость мембран и нарушить обменные процессы в клетке. В состав фосфолипидов входят две жирные кислоты, одна из которых, как правило, насыщенная, другая – ненасыщенная. Они составляют гидрофобный «хвост» фосфолипидов, обращенных друг к другу, а гидрофильные «головки», в которые входят фосфат-ион и положительная заряженная группа, обращены наружу (рис. 6.13).
Примерно половина поверхности двойного липидного слоя покрыта белками, легко от него отделяемыми. В самом слое находятся внутренние (трудно отделяемые) белки, встроенные в него на некоторую глубину. Внутренние белки содержат значительно большее число полярных аминокислотных остатков, чем те, которые находятся на поверхности липидного слоя. Неполярные остатки обращены преимущественно к жирнокислотной части липидов, полярные – в окружающую среду. Мемб ранные белки имеют, как правило, глобулярную третичную структуру. Радиационно-химические превращения фосфолипидов начинаются с разрушения двойной связи в ненасыщенной жирной кислоте.
В гидрофобных (водоотталкивающих) органических структурах, в основном жировых и жироподобных (липидных), облучение в присутствии кислорода вслед за ионизацией и возбуждением вызывает образование свободных радикалов и перекисей. Еще одним превращением фосфолипидов является отщепление неорганического фосфата. В силу своей химической
192 |
Радиобиология с основами радиоэкологии |
|
|
А
Б
Рис. 6.13. Модель (А) и схема (Б) клеточной мембраны.
реактивности и при доступе кислорода эти активные продукты взаимодействуют с нейтральными молекулами липидов, постепенно их окисляя.
Развиваются, таким образом, цепные реакции окисления органических соединений, в которых инициаторами выступают образованные воздействием радиации ионы, возбужденные электронные состояния и радикалы. Гидроперекиси и переки-
си могут образоваться в присутствии кислорода и за счет непо- |
|
средственного, прямого действия радиации: RH → R. |
|
R. + O → ROO. |
. |
. 2 |
|
ROO + RH → ROOH + R |
|
ROO. + R. → ROOR. |
|

6. Радиационно-химические и радиационно-биохимические процессы... |
193 |
Более 60 лет назад академик Н.Н. Семенов установил, что
цепные реакции могут быть трех типов: неразветвленные (со временем они угасают), разветвленные (в итоге каждого тура взаимодействия вместо одного радикала возникают три и реакция протекает лавинообразно) и вырожденно-разветвленные. В последнем случае при взаимодействии молекулы и радикала образуются новый радикал и новая молекула, но периодически вновь возникшая молекула распадается на два радикала, обуславливающие разветвление цепи.
Как показали исследования его сотрудника академика Н.М. Эмануэля, вызванное радиацией окисление органических соединений, прежде всего липидов, происходит согласно кинетике вырожденно-разветвленных реакций с постепенным вовлечением в этот процесс все большей массы вещества и нарастанием количества активных продуктов. Свободнорадикальные цепные реакции окисления протекают, таким образом, самоускоряясь. При развитии этих реакций возникают конечные продукты: липоперекиси, ненасыщенные жирные кислоты, эпоксиды, альдегиды и кетоны, обладающие токсическим действием. Некоторые из них при введении в организм обусловливают появление симптомов отравления, характерных для лучевого поражения.
Приведенные примеры радиационно-химических превращений биологически значимых веществ показывают ведущую роль в этих процессах свободных радикалов, их взаимодействие с кислородом с образованием перекисей и гидроперекисей органических веществ. Свободные радикалы также участвуют в возникновении цепных реакций под влиянием облучения, реакций полимеризации и деградации сложных органических веществ. Все это необходимо учитывать при рассмотрении значительно более сложных процессов, возникающих в клетках и тканях облученного организма.
Контрольные вопросы к главе 6
1.Перечислите основные процессы развития радиационного поражения во времени.
2.Какие стадии включает первичный этап действия ионизирующего излучения?
3.Какие процессы происходят при прямом действии радиации?
4.Какие процессы происходят при косвенном (непрямом) действии радиации?
5.Что такое свободный радикал и как он образуется?
6.Какие радиационно-химические превращения происходят в молекулах водных растворов и безводных системах?
7.Какова роль кислорода в радиационно-химических превращениях молекул воды?
8.Основные продукты радиолиза воды. Прямой и обратный кислородный эффект.
194 |
Радиобиология с основами радиоэкологии |
|
|
9.Радиационно-химические повреждения ДНК и РНК.
10.Радиационно-химические изменения, происходящие в белках, аминокислотах, углеводах.
11.Строение клеточной мембраны, ее состав. Какие химические превращения происходят в клеточных мембранах?
12.Роль образования свободных радикалов и цепных реакций в процессах окисления липидов.

7. Особенности действия ионизирующих излучений... |
195 |
7. ОСОБЕННОСТИ ДЕЙСТВИЯ ИОНИЗИРУЮЩИХ ИЗЛУЧЕНИЙ НА КЛЕТКИ И ТКАНИ.
КРИТИЧЕСКИЕ ТКАНИ И ОРГАНЫ
Костный мозг животных и меристемы растений – идентичные системы клеточного обновления организма. Радиочувствительность клеток в различные фазы цикла деления. Радиоустойчивость покоящихся клеток. Гибель клеток при облучении. Количественная оценка выживаемости клеток. Молекулярно-биохимические процессы, лежащие в основе гибели клеток. Относительная радиочувствительность клеток млекопитающих. Тканевая радиочувствительность. Критические ткани и органы. Особенности поражения клеток и тканей под влиянием инкорпорированных радионуклидов.
Еще на заре развития радиобиологии в 1906 г. французские исследователи И. Бергонье и Л. Трибондо, изучая радиационное поражение процессов сперматогенеза в различные периоды, отметили, что радиочувствительность ткани прямо пропор циональна пролиферативной активности – способности к де лению составляющих ее клеток – и обратно пропорциональна степени их дифференциации. Это положение, сформулированное ими в виде правила, является фактически одним из первых и основных законов радиобиологии.
Таким образом, ткани, клетки которых находятся в состоянии деления, должны обладать более высокой радиочувствительностью, чем клетки дифференцированных и специализированных тканей. И чем более четко выражена способность к делению у одного типа клеток, тем выше их радиочувствительность. Эта закономерность, несмотря на немногие исключения, является априори принятой в радиобиологии.
Процесс постоянной или достаточно продолжительной пролиферации свойственен лишь популяциям стволовых клеток обновляющихся тканей организма. Только такая клеточная популяция характеризуется способностью к самоподдержанию, т.е. сохранению неопределенно долго в отсутствии поступления клеток, способностью продуцировать дифференцированные клетки и в результате – восстанавливать нормальное количество клеток после повреждения. Только стволовые клетки имеют неограниченный пролиферативный потенциал, в то время как нестволовые размножающиеся клетки после некоторых делений неизбежно теряют эту способность или даже погиба-
196 |
Радиобиология с основами радиоэкологии |
|
|
ют. Стволовые клетки, способные к дифференциации и специализации, являются источником всех тканей организма, формирующегося в процессе эмбрионального и постэмбрионального развития.
У взрослых млекопитающих к таким обновляющимся тканям относятся кроветворная ткань, эпителий тонкого кишечника, эндотелий сосудов, клетки печени, селезенки и некоторые другие, ответственные за клеточное обновление лишь определенных тканей и органов. У растений таковыми считаются инициальные клетки (инициали) различных меристем – образовательных тканей, состоящих из ювенильных, способных к неопределенно долгому, иногда периодически возобновляемому делению клеток. Именно меристемы – верхушечные (апикальные), боковые (латеральные), вставочные (интеркалярные), краевые (маргинальные) и др. обусловливают увеличение размеров корней и побегов, а также образование новых органов в течение всего онтогенеза растения. Однако только инициальные клетки, оставаясь в эмбриональной фазе развития и делясь, обеспечивают непрерывное поступление клеток в меристемы, которые, как правило, после пяти-семи делений переходят к дифференциации и специализации.
Несмотря на то, что в функциональном отношении стволовые клетки соответствующих тканей у животных и растений выполняют различную роль, процессы их размножения и целенаправленных превращений у обоих типов организмов подчиняются общим законам.
7.1. Костный мозг животных и меристемы растений – аналогичные системы клеточного обновления организма
Одно из главных различий между животными и высшими растениями состоит в том, что если у животного организма накопление основной клеточной массы происходит на начальных этапах развития еще в эмбриональном состоянии, а последующий рост и развитие осуществляются за счет сформированного пула клеток за исключением стволовых, то у растений увеличение количества клеток происходит на протяжении всего онтогенеза – всей жизни. Этот процесс возможен именно благодаря пролиферации меристем.
Неограниченная способность к делению у инициальных клеток меристем, наличие пролиферирующей субпопуляции
испособность пролиферирующих клеток к дифференциации
ипоследующей специализации ставят меристемы растений в один ряд со многими постоянно обновляющимися тканями животных и в первую очередь – с костным мозгом.
Основное назначение как костного мозга, так и меристем – производство дифференцированных клеток. На рис. 7.1 при-

7. Особенности действия ионизирующих излучений... |
197 |
ведена схема клеточного обновления костного мозга из книги американского радиобиолога В. Бондом (см. список рекомендуемой литературы), дополненная сравнительными данными по меристеме. В ней рассматривается несколько субпопуляций, или пулов клеток. Исходным пулом являются стволовые, или клоногенные клетки. В красном костном мозге это кроветворные клетки, в меристеме – инициали. Количество их в обоих пулах не превышает 0.1% всех делящихся клеток. При делении стволовых клеток часть их, в нормальных условиях около половины, переходит в пул деления и созревания. Остальные продолжают выполнять функции стволовых.
Разделившись несколько раз и сформировавшись, эти клетки теряют способность к пролиферации и переходят в последующие пулы, где окончательно формируются и приступают к выполнению предназначенных им функций.
Кинетика процессов превращений клеток во времени определяется продолжительностью клеточного цикла, а так как она практически одинакова для клеток животных и растений, варьируя в основном в пределах 10–30 ч, то и весь этот процесс в зависимости от формирующегося элемента крови или вида растения составляет 3–7 сут.
Аналогичным путем протекают процессы клеточного обновления в эпителии ворсинок кишечника, коже, быстрорастущих опухолях, клетках животных и растений в культуре.
При действии ионизирующей радиации в системах клеточного обновления могут происходить определенные нарушения гомеостаза, характерного для нормы. В соответствии с законом
Рис. 7.1. Схема клеточного обновления костного мозга млекопитаю - щих (В. Бонд и др., 1971) и меристемы растений (И.Н. Гудков, 1979)
198 |
Радиобиология с основами радиоэкологии |
|
|
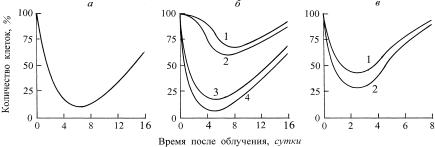
Бергонье и Трибондо, в первую очередь повреждаются активно делящиеся клетки. При относительно высоких дозах это обусловливает у части клеток задержку деления, нередко временную его остановку, полную потерю способности к делению, наконец, гибель клетки, что приводит к так называемому «опустошению» делящегося пула клеток. И в костном мозге, и в крови, и в меристеме клеточное опустошение начинается практически сразу же после облучения, и уже через несколько суток (несколько клеточных циклов) количество клеток в пуле (ткани, ее участке) достигает минимума (рис. 7.2).
В течение какого-то времени (1–2 сут.) это опустошение не сказывается на созревающих и функциональных клеточных пулах – организм в прежних количествах и соотношениях получает необходимые клетки, у животных не меняется формула крови, растения продолжают расти с прежней скоростью. Однако в последующем с продолжающимся истощением пула делящихся клеток у животных изменяется состав крови, у растений – замедляется рост органов. У большинства видов млекопитающих при дозах γ-излучения выше 10 Гр это опустошение приводит к прогрессирующей лейкопении, гранулопении, тромбоцитопении и гибели через одну-три недели. У растений в зависимости от радиочувствительности (у гороха – выше 12 Гр, кукурузы – 25 Гр, а бобов – всего 8 Гр) – к полной остановке роста, как правило, уже через неделю. Гибель всего растения может наблюдаться только через несколько недель.
Если доза облучения не оказалась фатальной, количество делящихся клеток начинает возрастать за счет деления клеток, сохранивших эту способность. При благоприятных условиях через некоторое время, зависящее от уровня радиационного повреждения, клеточная популяция данного пула может полностью восстановить свой первоначальный объем и нормализовать кинетику клеточного обновления всех пулов.
Рис. 7.2. Кинетика изменения количества делящихся клеток в пулах красного костного мозга (а) и периферической крови мышей (б: 1 – тромбоциты, 2 – нейтрофилы, 3 – лейкоциты, 4 – лимфоциты), а также апикальной меристемы корня (в: 1 – горох, 2 – кукуруза) после острого облучения в полулетальных дозах.

7. Особенности действия ионизирующих излучений... |
199 |
7.2. Радиочувствительность клеток в различные фазы цикла деления
Явление задержки деления клеток при действии ионизирующей радиации, оцениваемое по уменьшению количества митотических клеток в отдельных тканях, снижению скорости размножения одноклеточных организмов и другим признакам и показателям, известно давно. В сравнительно недавнем прошлом с разработкой методов оценки продолжительности отдельных фаз клеточного цикла (G1, S, G2 и M) и даже идентификации отдельных их периодов-субфаз (ранний, поздний) было показано, что такая задержка является следствием его блокирования на определенных стадиях, приуроченных, как правило, к периоду подготовки к синтезу ДНК (G1-, постмитотическая, или пресинтетическая фаза) либо митозу (G2-, постсинтетическая, или премитотическая фаза). Именно поэтому увеличение продолжительности клеточного цикла при облучении происходит преимущественно за счет возрастания длительности G1- и G2-фаз при относительной стабильности S- и М-фаз (рис. 7.3).
Все фазы деления клетки сопровождаются определенными метаболическими и структурными процессами ее целенаправленного превращения, которые обладают различной чувствительностью к повреждающим воздействиям, в том числе и ионизирующей радиации. Имеется довольно много экспериментальных данных о радиоустойчивости клеток, находящихся в различных фазах клеточного цикла, которая в зависимости не только от фазы, но и субфазы может изменяться в весьма широких пределах. Хотя эти данные, полученные на разных объектах, нередко существенно различаются и даже противоречат друг другу, в целом можно отметить, что по сведениям большинства исследователей, клетки, находящиеся в G2-фазе и митозе, обладают высокой радиочувствительностью, а находящиеся в S-фазе, как правило, более высокой радиоустойчивостью (рис. 7.4). Основные противоречия касаются G1-фазы, в которой клетки, по сведениям одних авторов, являются необычайно радиочувствительными, других – довольно радиоустойчивыми,
Рис. 7.3. Схематическое изображение увеличения продолжительности клеточного цикла при облучении за счет G1- и G2-фаз.

200 |
Радиобиология с основами радиоэкологии |
|
|
третьих – отдельные периоды G1-фазы значительно различаются между собой по чувствительности к радиации.
Считается, что причиной подобных противоречий является необычайно широкая вариабельность продолжительности этой фазы в клетках различных тканей и организмов по сравнению с другими. Так, если у клеток большинства организмов доля продолжительности S-фазы по отношению к длительности всего цикла составляет 30–60, митоза – 6–12, G2-фазы – 12–30%, то вклад в цикл G1-фазы варьирует от 1.5 до 50%. Показано, что если у объектов с короткой G1-фазой она, как правило, имеет высокую радиочувствительность, то с длинной G1-фазой ее начальный период обычно обладает высокой радиоустойчивостью, а последующий, по мере приближения к S-фазе, характеризуется последовательным повышением радиочувствительности. Маловероятно, что для относительно стабилизированного во времени синтеза ДНК требовалось бы столь разное время для его подготовки у различных клеток. Имеются все основания предполагать, что у клеток с длинной G1-фазой ее начальный период представляет состояние, близкое покою, обозначаемое G0-фазой, в которую клетка вступает после завершения митоза. На протяжении этой фазы решается ее последующая участь – деление или дифференциация. Иногда это состояние клетки называют «фазой выбора пути». Но иным клеткам с твердо предопределенной судьбой она просто не нужна.
Таким образом, по своей радиочувствительности пролиферирующие клетки самых различных организмов в зависимости от фазы деления могут различаться во много раз. Это является определяющим обстоятельством, обусловливающим наличие в асинхронно делящихся популяциях постоянно обновляющихся тканей в любой момент времени наряду с радиочувствительными, клеток, обладающих высокой радиоустойчивостью. Именно последние представляют собой главный источник восстановления пулов делящихся клеток в пострадиационный период.
Рис. 7.4. Радиочувствительность клеток на разных фазах митотического цикла:
а – человека (культура клеток НеLа), б – китайского хомячка (W.R. Sinclair, 1972) и в – корневой меристемы гороха (И.Н. Гудков, 1979).