

-
Нуклеиновые кислоты: рнк и днк, первичная структура.
Нуклеиновые кислоты занимают исключительное место в процессах жизнедеятельности живых организмов. Они осуществляют хранение и передачу генетической информации и являются инструментом, с помощью которого происходит управление биосинтезом белков.
Нуклеиновые кислоты представляют собой высокомолекулярные соединения (биополимеры), построенные из мономерных единиц — нуклеотидов, в связи с чем нуклеиновые кислоты называют также полинуклеотидами.
Структура каждого нуклеотида включает остатки углевода, гетероциклического основания и фосфорной кислоты. Углеводными компонентами нуклеотидов являются пентозы: D-рибоза и 2-дезокси-D-рибоза.
По этому признаку нуклеиновые кислоты делятся на две группы:
• рибонуклеиновые кислоты (РНК), содержащие рибозу;
• дезоксирибонуклеиновые кислоты (ДНК), содержащие дезоксирибозу.
ДНК содержатся в основном в ядрах клеток, РНК преимущественно находятся в рибосомах, а также протоплазме клеток. ДНК играют главную роль в передаче наследственных признаков (генетической информации). РНК участвуют в управлении процессом биосинтеза белка. Наиболее известны три вида клеточных РНК:
• матричные (мРНК);
• рибосомные (рРНК);
• транспортные (тРНК).
Первичная структура нуклеиновых кислот. ДНК и РНК имеют общие черты в структуре макромолекул:
• каркас их полинуклеотидных цепей состоит из чередующихся пентозных и фосфатных остатков;
• каждая фосфатная группа образует две сложноэфирные связи: с атомом С-З' предыдущего нуклеотидного звена и с атомом С-5' — последующего нуклеотидного звена;
• нуклеиновые основания образуют с пентозными остатками N-гликозидную связь.
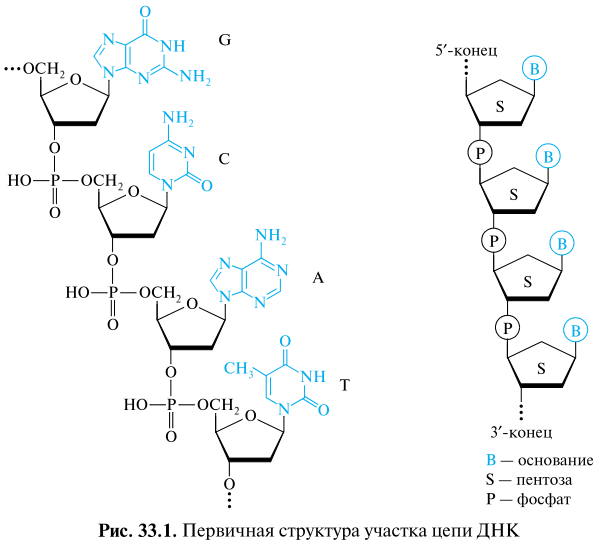
Приведено строение произвольного участка цепи ДНК, выбранного в качестве модели с включением в нее четырех основных нуклеиновых оснований — гуанина (G), цитозина (С), аденина (А), тимина (Т). Принцип построения полинуклеотидной цепи РНК такой же, как и у ДНК, но с двумя отличиями: пентозным остатком в РНК служит D-рибофураноза, а в наборе нуклеиновых оснований используется не тимин (как в ДНК), а урацил.
(!) Один конец полинуклеотидной цепи, на котором находится нуклеотид со свободной 5'-ОН-группой, называется 5'-концом. Другой конец цепи, на котором находится нуклеотид со свободной З'-ОН-группой, называется З'-концом.
Нуклеотидные звенья записываются слева направо, начиная с 5'-концевого нуклеотида. Запись строения цепи РНК осуществляется по таким же правилам, при этом буква «d» опускается.
С целью установления нуклеотидного состава нуклеиновых кислот проводят их гидролиз с последующей идентификацией полученных продуктов. ДНК и РНК ведут себя по-разному в условиях щелочного и кислотного гидролиза. ДНК устойчивы к гидролизу в щелочной среде, в то время как РНК очень быстро гидролизуются до нуклеотидов, которые, в свою очередь, способны отщеплять остаток фосфорной кислоты с образованием нуклеозидов. N-Гликозидные связи устойчивы в щелочной и нейтральной средах. Поэтому для их расщепления используется кислотный гидролиз. Оптимальные результаты дает ферментативный гидролиз с использованием нуклеаз, в том числе и фосфодиэстеразы змеиного яда, которые расщепляют сложноэфирные связи.
Наряду с нуклеотидным составом важнейшей характеристикой нуклеиновых кислот является нуклеотидная последовательность, т. е. порядок чередования нуклеотидных звеньев. Обе эти характеристики входят в понятие первичная структура нуклеиновых кислот.
Первичная структура нуклеиновых кислот определяется последовательностью нуклеотидных звеньев, связанных фосфодиэфирными связями в непрерывную цепь полинуклеотида.
Общий подход к установлению последовательности нуклеотидных звеньев заключается в использовании блочного метода. Сначала полинуклеотидную цепь направленно расщепляют с помощью ферментов и химических реагентов на более мелкие фрагменты (олигонуклеотиды), которые расшифровывают специфическими методами и по полученным данным воспроизводят последовательность строения всей полинуклеотидной цепи.
Знание первичной структуры нуклеиновых кислот необходимо для выявления связи между их строением и биологической функцией, а также для понимания механизма их биологического действия.
Комплементарность оснований лежит в основе закономерностей, которым подчиняется нуклеотидный состав ДНК. Эти закономерности сформулированы Э. Чаргаффом:
• количество пуриновых оснований равно количеству пиримидиновых оснований;
• количество аденина равно количеству тимина, а количество гуанина равно количеству цитозина;
• количество оснований, содержащих аминогруппу в положениях 4 пиримидинового и 6 пуринового ядер, равно количеству оснований, содержащих в этих же положениях оксогруппу. Это означает, что сумма аденина и цитозина равна сумме гуанина и тимина.
Для РНК эти правила либо не выполняются, либо выполняются с некоторым приближением, поскольку в РНК содержится много минорных оснований.
Комплементарность цепей составляет химическую основу важнейшей функции ДНК — хранения и передачи наследственных признаков. Сохранность нуклеотидной последовательности является залогом безошибочной передачи генетической информации. Изменение последовательности оснований в любой цепи ДНК приводит к устойчивым наследственным изменениям, а следовательно, и к изменениям в строении кодируемого белка. Такие изменения называют мутациями. Мутации могут происходить в результате замены какой-либо комплементарной пары оснований на другую. Причиной такой замены может служить сдвиг таутомерного равновесия.
Например, в случае гуанина сдвиг равновесия в сторону лактимной формы обусловливает возможность образования водородных связей с необычным для гуанина основанием — тимином и возникновение новой пары гуанин—тимин вместо традиционной пары гуанин—цитозин.
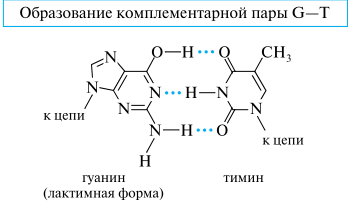
Замена «нормальных» пар оснований передается затем при «переписывании» (транскрипции) генетического кода с ДНК на РНК и приводит в итоге к изменению аминокислотной последовательности в синтезируемом белке.