

Генетика – наука о наследственности и изменчивости организма, лежит в сущности живого, т.к. связана с основными процессами жизнедеятельности.
Имеет теоретическое значение в философии и естествознании, практика – научная основа селекции (генмодифицмрованные продукты), причины возникновения, механизмы формирования и лечения заболеваний.
До ХХ века были попытки объяснить наследование признаков; накоплены сведения.
Германия, Франция, Англия – наследование признаков у внутривидовых и межвидовых гибридов → имеет место явление преобладания у потомков признаков одного из родителей.
Исключением стала работа Грегора Менделя 1865 г. «Опыты над растительными гибридами» (осталась без внимания).
1859 г. – Ч. Дарвин «Эволюционная теория…»: эволюция организмов возможна только на основе наличия изменчивости и её сохранения (наследственности).
Концепция слитной наследственности: наследственность определяется смешением кровей предковых форм. Из неё исходил Дарвин.
Теория пангенеза (Дарвин): полагал, что каждая клетка вырабатывает особые частицы – геммулы (пангены), которые током крови переносятся в половые клетки, включаются в них, передаются потомкам.
Теория зародышевой плазмы: 1883 г. Август Вейсман, в клетках организма имеется особая зародышевая плазма – носитель наследственности и сома, которая развивается на основе зародышевой плазмы. С целью примирить теоретические требования большого запаса изменчивости популяции со своими наблюдениями о том, что приобретенные признаки не наследуются. Отрезал мышам хвосты. Сома смертна, а зародышевая плазма живет.
Корпускулярная (дискретная) теория наследственности (Мендель):
наследственность корпускулярна и каждая из родительских особей передает своему потомку одинаковое число генов, за исключением тех организмов, у которых имеются половые хромосомы, у таких родителей вклады могут быть не одинаковы.
хотя некоторые гены могут подавлять или изменять действие других генов, наследственные факторы непосредственно не влияют друг на друга, т.е. фундаментальная структура гена не изменяется при ассоциации с другими генами и они передаются следующему поколению в той же форме в какой были получены предыдущими
гены исключительно стабильны по своему составу, но тем не менее они могут изменяться, что сопровождается изменением функции.
Законы Менделя.
В своих работах Мендель использовал гибридологический метод: скрещивание с получением гибридов и анализом результатов расщепления признаков по фенотипу потомков.
Особенности:
статистичность: большое количество материала, т.к. анализ косвенный, необходимо равновероятное соединение всех сортов гамет, равновероятная жизнеспособность зигот, полное проявление признаков
аналитичность: анализ от простого к сложному
использование альтернативных признаков, т.е. особи для скрещивания четко различаются по фенотипу
использование самоопылителей дает возможность убедится в чистоте линий
использование плодовитых линий, не вызывает регулярную потерю
проведение реципрокных скрещиваний, когда меняется пол обладателя признака, для выявления признаков, сцепленных с полом
принцип принудительного перекрестного опыления
Закон единообразия гибридов первого поколения.
В первом поколении все потомки были с желтыми горошинами. Тот признак, который проявляется в первом поколении – доминантный, признак, который исчез – рецессивный.
А – желтые горошины (доминантный признак)
а – зеленые горошины (рецессивный признак)
Р: ♀ АА х ♂ аа
F1: Aa – желтые
При скрещивании особей, отличающихся по одной паре признаков, в F1 наблюдается единообразие по фенотипу и генотипу.
Закон расщепления.
Р: ♀ Аа х ♂ Аа
F1: AA : Aa : aa
ж ж з
При скрещивании гибридов первого поколения во втором поколении наблюдается расщепление особей по фенотипу 3 : 1, по генотипу 1 : 2 : 1.
Закон независимого наследования.
При анализе дигибридного скрещивания.
А – желтые горошины
а – зеленые горошины
В – гладкая кожура
b – морщинистая кожура
P: ♀ AABB x ♂ aabb
F1: AaBb – желтые гладкие
Р2: ♀ АаВb x ♂ AaBb
F2: 9/16 – желтые гладкие А_В_
3/16 – зеленые гладкие ааВ_
3/16 – желтые морщинистые A_bb
1/16 – зеленые морщинистые ааbb
При скрещивании особей, отличающихся по 2 и более парам альтернативных признаков, наследование по каждой паре идет независимо от других.
Промежуточное наследование.
При скрещивании растений ночная красавица с белым и красным венчиком в первом поколении имели розовую окраску венчика. Имеет место неполное доминирование, т.е. подавление одного аллеля другим. При скрещивании гибридов первого поколения во втором поколении расщепление по генотипу и фенотипу совпадают.
Кодоминирование – оба аллеля проявляют себя в гетерозиготном состоянии.
Пр.: наследование групп крови системы MN
P: ♀ MM x ♂ NN
F1: MN
Ни один из аллелей не подавляет другой.
Аллельные гены – гены, расположенные в гомологичных локусах гомологичных хромосом.
При анализе был обнаружен ряд случаев, когда получающиеся фенотипические соотношения не укладывались в классическую схему.
При скрещивании желтых и черных мышей между собой в первом поколении получались желтые мыши, а во втором поколении получалось соотношение 2 : 1, т.е. часть эмбрионов гибнет (АА), желтые Аа – гетерозиготы, гомозиготы гибнут.
Было обнаружено, что некоторые признаки могут контролироваться не одним геном, а 2 и более. Характер взаимоотношения неаллельных генов могут быть различными, наиболее изучены:
комплиментарность
эпистаз
полимерия
Комплементарность – взаимодействие неаллельных, неэквивалентных или эквивалентных генов, которое сопровождается появлением нового признака. При комплиментарном взаимодействии может быть разное расщепление, в зависимости от того эквивалентные гены или нет, одинаковое или разное фенотипическое проявление.
окраска перьев у попугаев
А – желтые
а – белые
В – голубые
b - белые
Р: ♀ ААbb х ♂ ааВВ
F1: AaBb – зеленая окраска
P2: ♀ AaBb x ♂ AaBb
F2: 9/16 A_B_ - зеленые
3/16 A_bb – желтые
3/16 aaB_ - голубые
1/16 aabb – белые
форма плода у тыквы (эквивалентные гены)
А – сферическая форма
а – удлиненная форма
В – сферическая форма
b - удлиненная форма
Р: ♀ ААbb х ♂ ааВВ
F1: AaBb – дисковидная форма
P2: ♀ AaBb x ♂ AaBb
F2: 9/16 A_B_ - дисковидная форма
3/16 A_bb – сферическая форма
3/16 aaB_ - сферическая форма
1/16 aabb – удлиненная форма
Соотношение: 9:6:1
окраска шерсти у грызунов (один аллель имеет собственное фенотипическое проявление, а другой доминантный аллель выполняет регуляторную функцию, не имеет собственного фенотипического проявления)
А – черная окраска
а – белая окраска
В – распределение пигмента по волосу
b – белая окраска
Р: ♀ ААbb х ♂ ааВВ
F1: AaBb – окраска агути
P2: ♀ AaBb x ♂ AaBb
F2: 9/16 A_B_ - агути
3/16 A_bb – черный
3/16 aaB_ - белый
1/16 aabb – белый
Соотношение: 9 : 3 : 4
доминантные аллели не имеют фенотипического проявления, формирование признака только в присутствии 2 доминантных аллелей
А – пропигмент 1
а – нет синтеза
В – пропигмент 2
b – нет синтеза
Р: ♀ ААbb х ♂ ааВВ
F1: AaBb – окраска (есть пигмент)
P2: ♀ AaBb x ♂ AaBb
F2: 9/16 A_B_ - окраска (есть пигмент)
3/16 A_bb – нет пигмента
3/16 aaB_ - нет пигмента
1/16 aabb – нет пигмента
Соотношение: 9 : 7
Биохимические модели комплементарного взаимодействия, предложенные Вагнером и Митчеллом:
Последовательная модель – последовательность химических реакций при взаимодействии неаллельных генов (окраска у мышей).

Параллельная модель взаимодействия неаллельных генов (окраска попугаев)

Эпистаз – это тип взаимодействия неаллельных генов, когда один ген подавляет действие другого гена. Ген, который подавляет, называется эпистатическим или супрессором, подавляемый ген – гипостатический.
В зависимости от вида гена – супрессора выделяют:
Доминантный эпистаз
окраска шерсти у лошадей
А – вороная окраска
а – рыжая окраска
В – ранние поседение (серые)
b – нет поседения
Р: ♀ ААbb х ♂ ааВВ
F1: AaBb – серые
P2: ♀ AaBb x ♂ AaBb
F2: 9/16 A_B_ - серые
3/16 A_bb – вороная окраска
3/16 aaB_ - серые
1/16 aabb – рыжая окраска
Соотношение: 12 : 3 : 1
Может быть 13 : 3, когда фенотипическое проявление гена – супрессора совпадает с фенотипическим проявлением одного из аллелей, контролирующих признак
окраска кур
А – пестрая
а – белая
В – супрессор
b – не подавляет
Р: ♀ ААbb х ♂ ааВВ
F1: AaBb – белые
P2: ♀ AaBb x ♂ AaBb
F2: 9/16 A_B_ - белые
3/16 A_bb – пестрые
3/16 aaB_ - белые
1/16 aabb – белые
Соотношение: 13 : 3
Рецессивный эпистаз – ген – супрессор рецессивен
окраска плода у тыквы
А – желтые
а – зеленые
В – проявление
b – супрессор
Р: ♀ ААbb х ♂ ааВВ
F1: AaBb – желтые
P2: ♀ AaBb x ♂ AaBb
F2: 9/16 A_B_ - желтые
3/16 A_bb – белые
3/16 aaB_ - зеленые
1/16 aabb – белые
Соотношение: 9 : 3 : 4
бомбейский феномен – явление, когда на локус имеется более 2 аллелей называется множественным аллелизмом (группы крови). Эволюция – это увеличение разнообразия, увеличение изменчивости → повышение адаптивных возможностей.
H – проявление, h – супрессор
Р: ♀ IBI0hh x ♂ IAI0HH
F1: IAIBHh
Предположили, что имеется редкий рецессивный ген – супрессор, который подавляет синтез антигенов А и В.
Модель эпистаза по Вагнеру и Митчеллу: значение генов – супрессоров в том, что они обуславливают развитие нормального фенотипа не смотря на присутствие неаллельного мутантного гена.
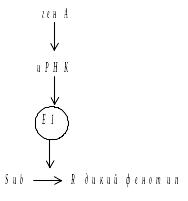

Взаимодействие с продуктом мутантного гена восстанавливает дикий фенотип
Включение запасного метаболического пути:
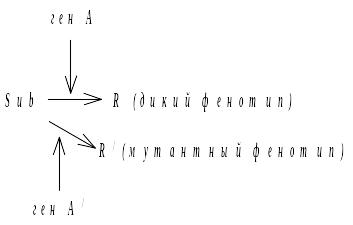
Ген – супрессор, действуя на другой субстрат, может приводить к формированию дикого фенотипа. Фермент работает на другом субстрате.
Полимерия – это взаимодействие неаллельных эквивалентных генов, действие которых может взаимно усиливаться.
Пр.: пигментация кожи у человека контролируется 2 парами эквивалентных неаллельных генов, действие которых суммируется кумулятивная полимерия
А1, А2 – выработка пигмента
а1, а2 – нет пигмента
Р: ♀ А1А1А2А2 х ♂ а1а1а2а2
F1: А1a1А2a2
P2: ♀ А1a1А2a2 x ♂ А1a1А2a2
F2: 1/16 А1A1А2A2 – негр
4/16 А1A1А2a2 + А1a1А2A2 – темный мулат (3 доминантных аллеля)
6/16 А1a1А2a2 + А1А1а2a2 + а1a1А2А2 – мулат (2 доминантных аллеля)
4/16 А1a1а2a2 + а1a1А2a2 – светлый мулат (1 доминантный аллель)
1/16 а1а1а2а2 – белый (рецессивные аллели)
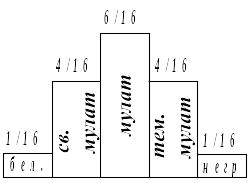
Признак варьирует от дозы гена, совместное действие аллелей усиливает проявление признака. Так наследуются преимущественно количественные признаки, чем больше генов контролирует признак, тем сложнее выделить отдельные фенотипические классы. Начиная с 5 пар, формируется непрерывный фенотипический ряд. Количественные признаки определяются по типу кумулятивной, накопительной полимерии.
Существуют качественные признаки, которые тоже определяются полимерными генами, но в этом случае признак не варьирует от дозы гена и для полного развития признака достаточно одного аллеля.
Пр.: форма плода у пастушьей сумки (некумулятивная полимерия).
А1, А2 – треугольная
а1, а2 – овальная
Р: ♀ А1А1А2А2 х ♂ а1а1а2а2
F1: А1a1А2a2
P2: ♀ А1a1А2a2 x ♂ А1a1А2a2
F2: 1/16 А1A1А2A2 – треугольная
4/16 А1A1А2a2 + А1a1А2A2 – треугольная
6/16 А1a1А2a2 + А1А1а2a2 + а1a1А2А2 – треугольная
4/16 А1a1а2a2 + а1a1А2a2 – треугольная
1/16 а1а1а2а2 – овальная
Соотношение: 15 : 1
Гипотезы возникновения полимерии:
происхождение полимерии возможно при скрещивании 2 близких форм, имеющих одинаковые доминантные гены с одним и тем же действием. При таком скрещивании у гибрида могло произойти удвоение набора хромосом.
путем различных хромосомных перестроек – неравный кроссинговер

Биологическое значение некумулятивной полимерии – это резервирование изменчивости. Данные, полученные в результате изучения наследования количественных признаков, показали, что большая часть признаков организма обусловлены множественными генами, анализ их весьма сложен, т.к.
число полимерных генов может быть различным
сила действия и значение гена может быть специфично
каждый из генов может иметь разную степень доминирования