

67 Стадійний розвиток. (див попереднє питання) Стадія яровизації.
Залежність переходу рослин до цвітіння від температури – яровизація. Яровизація (верналізація) – процес, що протікає в озимих форм однорічних і дворічних рослин під дією низьких позитивних температур певної тривалості і який сприяє прискореному розвитку цих рослин.Рослини, які потребують яровизації, називають озимими, а ті, які розвиваються без неї – ярими.Озимі види без яровизації не зацвітають (якісна реакція); у деяких при дії низьких температур лише вкорочується час переходу до цвітіння (кількісна реакція). У рослин із кількісною реакцією яровизація може відбуватися в накільченому насінні. Рослини з якісною реакцією повинні досягнути певних розмірів (певного етапу розвитку) до її початку.Яровизація звичайно продовжується 1-3 місяці. Найефективнішими є температури від 0 до +70 С; для теплолюбних рослин - +10-120 С.Для деяких видів (жито, капуста) періодичне переривання низькотемпературної дії знижує або і нівелює (усуває) ефект яровизації.Необхідна умова яровизації – наявність клітин, що діляться. Даний процес протікає в зародку, або в апікальних меристемах стебла і молодих листків. Яровизаційні зміни передаються лише шляхом поділу клітин. Фізіолого-біохімічна суть яровизації до кінця не вивчена.Після яровизації для більшості рослин необхідні довгоденні фотоперіоди. В інших рослин яровизація послаблює потребу у фотоперіодичній дії. Для деяких рослин безперервне освітлення сприяє переходові рослин від озимої поведінки до типово ярової. Т.Д.Лисенко широко впроваджував метод яровизації і вважається одним з його авторів. Після "розгрому лисенковщини" метод поступово використався все рідше та рідше, бо був дуже трудомістким.
68. Субмікроскопічна будова рослинної клітини. Будова та ф- ї основних органел.
Будова рослинної КЛІТИНИ Рослинна клітина складається з більш-менш жорсткої клітинної оболонки і протопласта. Клітинна оболонка - це клітинна стінка і цитоплазматична мембрана. Термін протопласт походить від слова протоплазма, яке довгий час використовувалася для позначення всього живого. Протопласт - це протоплазма індивідуальної клітини. Протопласт складається з цитоплазми і ядра. У цитоплазмі знаходяться органели (рибосоми, мікротрубочки, пластиди, мітохондрії) та мембранні системи (ендоплазматичний ретикулум, діктіосоми). Цитоплазма включає в себе ще цитоплазматичний матрикс (основна речовина) в яке занурені органели і мембранні системи. Від клітинної стінки цитоплазма відокремлена плазматичною мембраною, яка представляє собою елементарну мембрану. На відміну від більшості тварин клітин рослинні клітини містять одну або кілька вакуоль. Це бульбашки, заповнені рідиною і оточені елементарної мембраною (тонопластом). Плазматична мембрана. Являє собою біслойную вфосфоліпідну структуру. Для рослинних клітин властиві впячивания плазматичної мембрани. Плазматична мембрана виконує наступні функції: -Бере участь в обміні речовин між клітиною та навколишнім середовищем; -Координує синтез і збірку целюлозних мікрофібрил клітинної стінки; -Передає гормональні та зовнішні сигнали, які контролюють ріст і диференціювання клітин. Ядро. Це найбільш помітна структура в цитоплазмі еукаріотичної клітини. Ядро виконує дві важливі функції: -Контролює життєдіяльність клітки, визначаючи, які білки, і в який час має поєднуватися; -Зберігає генетичну інформацію і передає її дочірнім клітинам в процесі клітинного поділу. Зовнішня мембрана ядерної оболонки в деяких місцях об'єднується з ендоплазматичним ретикулумом. У забарвленому спеціальними барвниками ядрі можна розрізнити тонкі нитки і грудочки хроматину і нуклеоплазми (основна речовина ядра). Хроматин складається з ДНК, пов'язаної із спеціальними білками - гістонами. У процесі клітинного ділення хроматин все більше ущільнюється і збирається в хромосоми. У ДНК закодована генетична інформація. Організми розрізняються за кількістю хромосом у соматичних клітинах. Наприклад, капуста має - 20 хромосом; соняшник - 34, пшениця - 42; людина - 46, а один з видів папороті Ophioglossum – 1250 Під світловим мікроскопом можна розглянути сферичні структури - ядерця. У кожному ядрі є одне або кілька ядерець, які помітні в неподільних ядрах. Біохімічно ядерця характеризуються високою концентрацією РНК, яка тут пов'язана з фосфопротеідамі.
Пластиди. Вакуолі, целюлозна клітинна стінка і пластиди - характерні компоненти рослинних клітин. Кожна пластиду має власну оболонку, що складається з двох елементарних мембран. Усередині пластиди розрізняють мембранну систему і різного ступеня гомогенне речовина - строму. Зрілі пластиди класифікують на підставі містяться в них пігментів. Хромопласти - пігментовані пластиди. Різноманітні за формою вони не мають хлорофілу, але синтезують і накопичують каротиноїди, які надають жовтого, помаранчеву, червоне забарвлення квіток, старим листям, плодів і коріння. хромопласти можуть розвиватися з хлоропластів, які при цьому втрачають хлорофіл і внутрішні мембранні структури, накопичують каротиноїди. Це відбувається при дозріванні багатьох плодів. Хромопласти приваблюють комах та інших тварин, з якими вони разом еволюціонували. Лейкопласти - непігментовані пластиди. Деякі з них синтезують крохмаль (амілопласти), інші здатні до утворення різних речовин, у тому числі ліпідів і білків. На світлі лейкопласти перетворюються у хлоропласти. Пропластид - дрібні безбарвні або блідо-зелені недиференційовані пластиди, які знаходяться в меристематичних (діляться) клітинах коренів і пагонів. Вони є попередниками інших, більш диференційованих пластид - хлоропластів, хромопластів і амінопласти.
Мітохондрії. Як і хлоропласти, мітохондрії оточені двома елементарними мембранами. Внутрішня мембрана утворює безліч складок і виступів - крист, які значно збільшують внутрішню поверхню мітохондрії. Вони значно менше, ніж пластиди, мають близько 0,5 мкм в діаметрі і різноманітні по довжині і формі. У мітохондріях здійснюється процес дихання, в результаті якого органічні молекули розщеплюються з вивільненням енергії і передачею її молекул АТФ, основного резерву енергії всіх еукаріотичних клітин. Більшість рослинних клітин містить сотні й тисячі мітохондрій. Їх кількість в одній клітці визначається потребою клітини в АТФ. Мікротільця. На відміну від пластид і мітохондрій, які відмежовані двома мембранами, мікротільця представляють собою сферичні органели, оточені однієї мембраною. Мікротільця мають гранулярний (зернисте) вміст, іноді в них зустрічаються і кристалічні білкові включення. Мікротільця пов'язані з одним або двома ділянками ендоплазматичного ретикулуму. Деякі мікротільця, звані проксісомамі, відіграють важливу роль у метаболізмі гліколевої кислоти, що має безпосереднє відношення до фотодихання. У зеленому листі вони пов'язані з мітохондріями і хлоропластами. Інші мікротільця, звані, гліоксісомамі, містять ферменти, необхідні для перетворення жирів у вуглеводи. Це відбувається в багатьох насінні під час проростання. Вакуолі - це відмежовані мембраною ділянки клітини, заповнені рідиною - клітинним соком. Вони оточені тонопластом (вакуолярної мембраною). Молода рослинна клітина містить численні дрібні вакуолі, які в міру старіння клітини зливаються в одну велику. У зрілій клітці вакуолью може бути зайнято до 90% її обсягу. При цьому цитоплазма притиснута у вигляді тонкого шару периферичного до клітинної оболонки. Збільшення розміру клітини в основному відбувається за рахунок зростання вакуолі. У результаті цього виникає тургорное тиск і підтримується пружність тканини. У цьому полягає одна з основних функцій вакуолі і тонопластом. Основний компонент соку - вода, інші варіюють залежно від типу рослини і його фізіологічного стану. Вакуолі містять солі, цукру, рідше білки. Тонопласт відіграє активну роль у транспорті і накопиченні в вакуолі деяких іонів. Концентрація іонів у клітинному соку може значно перевищувати її концентрацію у навколишньому середовищі. При високому вмісті деяких речовин у вакуолях утворюються кристали. Найчастіше зустрічаються кристали оксалату кальцію, що мають різну форму. Вакуолі - місця накопичення продуктів обміну речовин (метаболізму). Це можуть бути білки, кислоти і навіть отруйні для людини речовини (алкалоїди). Часто відкладаються пігменти.
Рибосоми. Маленькі частинки (17 - 23нм), що складаються приблизно з рівної кількості білка і РНК. У рибосомах амінокислоти з'єднуються з утворенням білків. Їх більше в клітинах з активним обміном речовин. Рибосоми розташовуються в цитоплазмі клітини вільно або ж прикріплюються до грубого ендоплазматичного ретикулуму (80S). Їх виявляють і в ядрі (80S), мітохондріях (70S), пластидах (70S). Рибосоми можуть утворювати комплекс, на яких відбувається одночасний синтез однакових поліпептидів, інформація про яких знімається з однієї молекули і РНК. Такий комплекс називається полірібосомамі (полісомах). Клітини, які синтезують білки у великих кількостях, мають велику систему полісом, які часто прикріплюються до зовнішньої поверхні оболонки ядра. Ендоплазматичний ретикулум. Це складна тривимірна мембранна система невизначеною протяжності. У розрізі ЕР виглядає як дві елементарні мембрани з вузьким прозорим простором між ними. Форма і довжина ЕР залежать від типу клітини, її метаболічної активності та стадії диференціювання. У клітинах, які секретують або запасающих білки, ЕР має форму плоских мішечків або цистерн, з численними рибосомами, пов'язаними з його зовнішньою поверхнею. Такий ретикулум називається шорстким ендоплазматичним ретикулумом. Гладкий ЕР зазвичай має трубчасту форму. Шорсткий і гладкий ендоплазматичний ретикулум можуть бути присутніми в одній і тій же клітині. Як правило, між ними є багато чисельні зв'язку. Ендоплазматичний ретикулум функціонує як комунікаційна система клітини. Він пов'язаний із зовнішньою оболонкою ядра. Фактично ці дві структури утворюють єдину мембранну систему. Коли ядерна оболонка під час поділу клітини розривається, її обривки нагадують фрагменти ЕР. Ендоплазматичний ретикулум - це система транспортування речовин: білків, ліпідів, вуглеводів, в різні частини клітини. ендоплазматичний ретикулум сусідніх клітин з'єднуються через цитоплазматичні тяжі - плазмодесми - які проходять крізь клітинні оболонки. Ендоплазматичний ретикулум - основне місце синтезу клітинних мембран. У деяких рослинних клітинах тут утворюються мембрани вакуолей і мікротелец, цистерни діктіосом. Апарат Гольджі. Цей термін використовується для позначення всіх діктіосом, або тілець Гольджі, у клітці. Діктіосоми - це групи плоских, дисковидні бульбашок, або цистерн, які по краях розгалужуються в складну систему трубочок. Діктіосоми у вищих рослин складаються з 4 - 8 цистерн, зібраних разом. Зазвичай в пачці цистерн розрізняють формується і дозрівають сторони. мембрани формуються цистерн по структурі нагадують мембрани ЕР, а мембрани дозріваючих цистерн - плазматичну мембрану. Мікротрубочки виявлені практично у всіх еукаріотичних клітинах. Представляють собою циліндричні структури діаметром близько 24 нм. Довжина їх варіює. Кожна трубочка складається з субодиниць білка, званого тубуліну. Мікротрубочки - це динамічні структури, вони регулярно руйнуються і утворюються на певних стадіях клітинного циклу. Їх складання відбувається в особливих місцях, які називаються центрами організації мікротрубочок. У рослинних клітинах вони мають слабко виражені аморфну структуру. Функції мікротрубочок: беруть участь в утворенні клітинної оболонки; направляють бульбашки діктіосом до формується оболонці, подібно ниток веретена, які утворюються в делящейся клітці; відіграють певну роль у формуванні клітинної пластинки (первісної кордону між дочірніми клітинами). Крім того, мікротрубочки - важливий компонент джгутиків і війок, в русі яких, грають важливу роль. Мікрофіламенти, подібно мікротрубочки, знайдені практично у всіх еукаріотичних клітинах. Являють собою довгі нитки товщиною 5 - 7 нм, що складаються з скорочувального білка актину. Пучки мікрофіламентів зустрічаються в багатьох клітинах вищих рослин. Мабуть, грають важливу роль в струмах цитоплазми. Мікрофіламенти разом з мікротрубочками утворюють гнучку мережу, звану цитоскелетом. Ліпідні краплі - структури сферичної форми, що надають гранулярність цитоплазмі рослинної клітини під світловим мікроскопом. На електронних мікрофотографіях вони виглядають аморфними. Дуже схожі, але більш дрібні краплі зустрічаються в пластидах. Ліпідні краплі, приймаючи за органели, називали їх сферосомамі і вважали, що вони оточені одно-або двошарової мембраною. Проте останні дані показують, що у ліпідних крапель мембран немає, але вони можуть бути покриті білком. Клітинна стінка. Клітинна стінка відмежовує розмір протопласта і оберігає його розрив за рахунок поглинання води вакуолью. Клітинна стінка має специфічні функції, які важливі не тільки для клітини і тканини, в якій клітина знаходиться, але і для всієї рослини. Клітинні стінки грають істотну роль у поглинанні, транспорті і виділення речовин, а, крім того, в них може бути зосереджена лізосомальних, або переваривающая активність. Компоненти клітинної стінки. Найбільш типовим компонентом клітинної стінки є целюлоза, яка значною мірою визначає її архітектуру. молекули целюлози складаються з повторюваних молекул глюкози, з'єднаних кінець до кінця. Довгі тонкі молекули целюлози об'єднані в мікрофібрил товщиною 10 - 25 нм. Целюлозний каркас клітинної стінки заповнений переплітаються з ним целюлозними молекулами матриксу. До його складу входять полісахариди, звані геміцелюлозами, і пектинові речовини, або пектини, хімічно дуже близькі до геміцелюлози. Інший компонент клітинної стінки - лігнін - є найпоширенішим після целюлози полімером рослинних клітин. Лігнін збільшує жорсткість стінки і зазвичай міститься в клітинах, що виконують опорну або механічну, функцію. Кутін, суберіном, воску - зазвичай відкладаються в оболонках захисних тканин рослин.
69. Темнова фаза фотосинтезу. Фаза карбоксилювання ,відновлення, регенерації.
Темнова фаза фотосинтезу або цикл Кальвіна (С-3 шлях фотосинтезу).
Запасена в світловий фазі фотосинтезу енергія витрачається на процес перетворення неорганічної форми вуглецю (вуглекислого газу або бікарбонат-іона) в органічну, тобто фактично витрачається на створення простих цукрів, з яких потім формуються полісахариди. Практично у всіх рослин відбувається процес, званий циклом Кальвіна, в якому йде перетворення неорганічного вуглецю в органічний. Процес поділяється на три етапи:
карбоксилювання, коли СО 2 сполучається з рибулезо-1-5-дифосфат, утворює нестійке шестіуглеродное з'єднання, яке розпадається на дві молекули фосфогліцеринової кислоти - 3-вуглецеві молекули,
фазу відновлення, коли утворилася фосфогліцеринової кислота відновлюється в фосфогліцеринової альдегід - це центральна ланка циклу, так як ФГА за рівнем восстановленности вуглецю відповідає вуглеводи з загальною формулою (СН 2 О) 3, фазу регенерації, коли знову утворюється первинний акцептор СО 2 - рібулезодіфосфат - і , одночасно, відбувається синтез кінцевих продуктів фотосинтезу - глюкози і крохмалю.
Оскільки в циклі Кальвіна первинними продуктами включення неорганічного вуглецю в органічний є трьохвуглецевих з'єднання, цей процес носить назву С-3 шлях фотосинтезу.
Для синтезу однієї молекули глюкози має відбутися шість обертів циклу Кальвіна. У кожному обороті використовуються три молекули АТФ (дві для активування двох молекул фосфогліцеринової кислоти і одна при регенерації рібулезодіфосфата) і дві молекули НАДФ. Н 2 для відновлення кислоти в альдегід. Таким чином для синтезу однієї молекули глюкози необхідно витратити 12 молекул НАДФ. Н 2 і 18 молекул АТФ.
Важливо відзначити, що фізіологічне значення циклу Кальвіна полягає не тільки в акцепцію вуглекислого газу, а й у створенні маси вуглеводних сполук, які йдуть як на синтез запасних речовин, так і на створення компонентів хлоропласта та поточний метаболізм клітини. .
З-4 шлях фотосинтезу.
Більшість рослин засвоює неорганічний вуглець саме шляхом циклу Кальвіна.
70. теорія біологічного окислення Баха та Полудіна
Відповідно до теорії радянського біохіміка А. Н. Баха, процес дихання, тобто окислення вуглеводів, жирів, білків, здійснюється за допомогою окисної системи клітини в два етапи[1]:
активація кисню (O2) повітря шляхом його приєднання до ненасичених, здатних мимоволі окислюватися сполукам, що містяться в живій клітині (оксигенази) з утворенням перекисів,
активація перекисів із звільненням атомарного кисню, здатного окислювати важкоокислювані органічні речовини.
За теорією дегідрування російського ботаніка В. І. Палладіна, найважливіша ланка дихання — активація водню субстрату, здійснювана дегідрогеназою. Обов'язковий учасник складного ланцюга процесів дихання — вода, водень якої разом з воднем субстрату використовується для відновлення самоокислюваних сполук — так званих дихальних пігментів. Вуглекислий газ (CO2), що виділяється при диханні, утворюється без участі кисню повітря, тобто анаеробно. Кисень повітря йде на окислення дихальних хромогенів, що перетворюються при цьому в дихальні пігменти. Подальший розвиток теорія дихання отримала в дослідженнях радянського ботаніка С. П. Костичева, згідно з якими перші етапи аеробного дихання аналогічні процесам, властивим анаеробам. Перетворення утворюваного при цьому проміжного продукту можуть йти з участю кисню, що властиво для аеробів. У анаеробів ж ці перетворення йдуть без участі молекулярного кисню.
За сучасними уявленнями, процес окислення, який становить хімічну основу дихання, полягає у втраті речовиною електрона. Здатність приєднувати або віддавати електрони залежить від величини окисного потенціалу з'єднання.
71 типи вуглецевого живлення рослин. Фотосинтез, загальне рівняння
Фотосинтез — єдиний процес у біосфері, який веде до збільшення вільної енергії біосфери за рахунок зовнішнього джерела — Сонця і забезпечує існування як рослин, так і всіх гетеротрофних організмів, у тому числі й людини.
Загальне рівняння фотосинтезу не досить точно передає суть цього процесу, адже, згідно з цим рівнянням, можна припустити, що частина виділеного О2 походить від СО2, тоді як насправді весь кисень утворюється з води в ході фотолізу:
![]()
Тому доцільніше розділити його на складові частини:
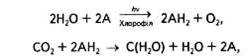
де А — акцептор електронів і протонів.
Згідно з цим рівнянням, в основі суті фотосинтезу лежить окисно-відновна реакція. За участі хлорофілу та енергії сонячних квантів вода фотоокиснюється, в результаті чого виділяються кисень та водень, останній і відновлює СО2 до рівня вуглеводів. Ці реакції відбуваються відповідно в світлову та темпову стадії фотосинтезу.
Типи живлення. Залежно від джерела вуглецю, що поглинається, розрізняють декілька типів П. р. Частина нижчих рослин (всі гриби і велика частина бактерій) може використовувати вуглець лише з органічних сполук, в яких він міститься у відновленій формі. При окисленні таких з'єднань в процесі дихання звільняється запасена в них хімічна енергія, яка потім може витрачатися на різні ендергонічеськие (тобто енергії, що вимагають витрат) процеси: синтез складніших з'єднань, пересування речовин в рослині і ін. Живлення цього типа називається гетеротрофним, а рослини, споживаючі органічні джерела вуглецю, - гетеротрофними (див. Гетеротрофні організми ) ; живлення за рахунок мертвих органічних залишків називається сапрофітним, а рослини, що харчуються мертвими органічними залишками, - сапрофітами . Цей тип живлення властивий всім гнильним грибам і бактеріям. Гетеротрофи, що живуть за рахунок органічних сполук ін. живих організмів, називаються паразитами . До них відносяться всі гриби і бактерії — збудники хвороб тварин і рослин, а також деякі вищі рослини, наприклад вовчок, що висмоктує за допомогою спеціальних присосків соки ін. рослин. Паразитичне П. р. відрізняється від симбіозу, при якому відбувається постійний обмін продуктами життєдіяльності корисний для обох партнерів. Симбіотичний П. р. спостерігається, наприклад, у азотфіксуючих бактерій, поселяющихся в бульбах на корінні бобових рослин (див. Азотфіксация ) , в капелюшних грибів, гіфи яких проникають в кореневі тканини деревних рослин (див. Мікориза ) , а також в лишайників, що є групою грибів що знаходяться в постійному співжитті з водоростями. Велика частина рослин здатна засвоювати вуглець з вуглекислого газу, відновлюючи його до органічних сполук. Цей тип живлення називається автотрофним (див. Автотрофні організми ) . Він властивий всім вищим зеленим рослинам, а також водоростям, деяким бактеріям. Відновлення Co 2 до органічних сполук вимагає витрат енергії або за рахунок сонячного світла (фотосинтетики), що поглинається, або за рахунок окислення відновлених з'єднань, що поглинаються із зовнішнього середовища (хемосинтетіки).
В процесі фотосинтезу рослини не лише поглинають речовини, але і нагромаджують енергію. Один з первинних продуктів фотосинтезу — цукру. При з'єднанні 6 грам-молекул Co 2 і такої ж кількості H 2 O утворюється 1 грам-молекула глюкози (180 г ) . Цей процес відбувається з поглинанням 674 ккал (1 ккал = 4,19 кдж ) енергії сонячного світла, яка і запасається в хімічних зв'язках цукру. Разом з молекулами цукру ця запасена хімічна енергія може потім переміститися в інших, нефотосинтезуючі частини рослин, наприклад в корінь. Тут в процесі дихання вона може звільнятися для синтезу складніших з'єднань і для ін. процесів життєдіяльності рослинних кліток. Хоча у фотосинтезі безпосередньо беруть участь лише Co 2 і H 2 O, для його здійснення і особливо для подальших перетворень його первинних продуктів необхідні всі ін. елементи П. р., в яких би нікчемних кількостях вони не містилися в рослині.
72.історія відкриття і значення фотосинтезу
Фотоси́нтез (від грец. φωτο- — світло та грец. σύνθεσις — синтез, сукупність) — процес синтезу органічних сполук з вуглекислого газу та води з використанням енергії світла й за участю фотосинтетичних пігментів: (хлорофіл у рослин, хлорофіл, бактеріохлорофіл і бактеріородопсин у бактерій), часто з виділенням кисню як побічного продукту. Це надзвичайно складний процес, що включає довгу послідовність координованих біохімічних реакцій. Він відбувається у вищих рослинах, водоростях, багатьох бактеріях, деяких археях і найпростіших — організмах, відомих разом як фототрофи. Сам процес відіграє важливу роль у кругообігу вуглецю у природі.
Фотосинтез – єдиний процес у біосфері, який призводить до засвоєння енергії Сонця і забезпечує існування як рослин, так і всіх гетеротрофних організмів.[1]
Узагальнене рівняння фотосинтезу (брутто-формула) має вигляд:
6СО2 + 6Н2О = С6Н12О6 + 6О2
На початку XVII в. фламандський лікар Ван Гельмонт виростив в кадці із землею дерево, яке він поливав тільки дощовою водою. Він відмітив, що через п'ять років дерево виросло до великих розмірів, хоча кількість землі в кадці практично не зменшилась. Ван Гельмонт, природно, зробив висновок, що матеріал, з якого утворилося дерево був отриманий з води, що використовувалась для поливу. У 1777 році англійський ботанік Стівен Хейлс опублікував книгу, в якій повідомлялось, що в якості поживної речовини, необхідної для зростання, рослини використовують головним чином повітря. У той же період знаменитий англійський хімік Джозеф Прістлі (він був одним з першовідкривачів кисню) провів серію дослідів по горінню і диханню і прийшов до висновку про те, що зелені рослини здатні здійснювати всі ті дихальні процеси, які були виявлені в тканинах тварин. Через декілька років голландський лікар Інгенхауз виявив, що рослини окисляють кисень лише на сонячному світлі і що тільки їх зелені частини забезпечують виділення кисню. Жан Сенебье, що займав пост міністра, підтвердив дані Інгенхауза і продовжив дослідження, показавши, що в якості поживної речовини рослини використовують двоокис вуглецю, розчинений у воді.
У 1817 р. два французькі хіміки, Пельтьє і Каванту, виділили з листя зелену речовину і назвали її хлорофілом. Наступною важливою віхою в історії вивчення фотосинтезу було зроблене в 1845 р. німецьким фізиком Робертом Майєром твердження про те, що зелені рослини перетворюють енергію сонячного світла в хімічну енергію.
Типии росту рослин

Транспирація Види транспірації,показники транспірації


75. Ферменти реакції дихання






76. Фізичні та оптичні властивості хлорофілу





77. Фізіологічна роль мікроелементів



