

Роль термогенина в снижении протонного градиента
Вещества, которые снижают величину электрохимического градиента, приводя к увеличению скорости движения электронов по ферментам дыхательной цепи, уменьшению синтеза АТФ и возрастанию катаболизма называются разобщители. Они разобщают (разъединяют) процессы окисления и фосфорилирования внутренней мембраны митохондрий. К разобщителям в первую очередь относят "протонофоры" – вещества переносящие ионы водорода. При этом одновременно уменьшаются оба компонента электрохимического градиента – электрический и химический, и энергия градиента рассеивается в виде тепла. Следствием эффекта протонофоров является возрастание катаболизма жиров и углеводов в клетке и во всем организме.
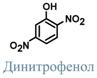
Классическим протонофором является динитрофенол, жирорастворимое соединение, присоединяющие ионы водорода на внешней поверхности внутренней митохондриальной мембраны и отдающие их на внутренней поверхности. Белок термогенин является физиологическим протонофором.
Кроме динитрофенола и термогенина протонофорами, к примеру, являются салицилаты, жирные кислоты итрийодтиронин.
Ингибиторы ферментов дыхательной цепи
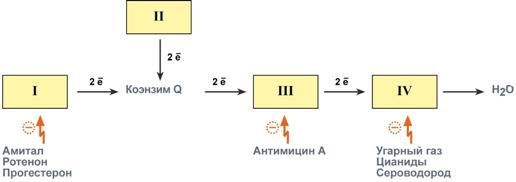
Ряд веществ может ингибировать ферменты дыхательной цепи и блокировать движение электронов от НАДН и ФАДН2 на кислород. Они называются ингибиторы. В результате прекращается движение электронов, выкачивание ионов Н+ и работа АТФ-синтазы. Синтез АТФ отсутствует и клетка погибает. Выделяют три основных группы ингибиторов:
действующие на I комплекс, например, амитал (производное барбитуровой кислоты), ротенон,прогестерон,
действующие на III комплекс, например, экспериментальный антибиотик антимицин А,
действующие на IV комплекс, например, сероводород (H2S), угарный газ (СО), цианиды (-CN).
Ингибиторы ферментов дыхательной цепи Как подсчитать эффективность окисления? Коэффициент p/o
Энергетическую ценность и выгодность окисления вещества можно подсчитать. Количество запасенной энергии при окислении того или иного соединения характеризует количество АТФ, а эффективность использования энергии вещества и ее захвата – коэффициент P/O.
Коэффициент Р/О – это отношение количества неорганического фосфата, включенного в молекулу АТФ АТФ-синтазой к количеству атомов кислорода, включенного в молекулу Н2О, при переносе одной пары электронов по дыхательной цепи.
Экспериментально установлено, что при окислении веществ с участием НАД-зависимых дегидрогеназ, соотношение количества включенного в АТФ фосфата относится к количеству использованных атомов кислорода как 3 к 1, т.е. коэффициент P/O равен трем. Аналогично для ФАД-зависимых дегидрогеназ коэффициент P/O равен двум.
Расчет энергетической ценности и коэффициента р/о
При расчете энергетической ценности, т.е. количества АТФ, образующейся при окислении вещества, и коэффициента Р/О необходимо представлять себе весь путь этого вещества до полного окисления его углеродных атомов в СО2. При этом необходимо учитывать число атомов углерода в молекуле.
Для расчета Р/О при окислении какой-либо молекулы необходимо учитывать следующее:
восстановленный эквивалент (молекула НАДН или ФАДН2) передает в цепь переноса электронов по 2 электрона.
для восстановления кислорода в воду необходима 1 пара электронов.
при прохождении пары электронов через всю дыхательную цепь (т.е. через I, III, IV комплексы) выкачивается столько ионов Н+, сколько необходимо для синтеза 3 молекул АТФ или, по-другому, в состав АТФ включается три атома неорганического фосфата.
при прохождении пары электронов через III и IV комплексы дыхательных ферментов ионов Н+выкачивается столько, сколько необходимо для синтеза 2 молекул АТФ или, по-другому, в состав АТФ включается два атома неорганического фосфата.
В качестве самых простых примеров рассмотрим окисление аланина и аспарагиновой кислоты.
Окисление аспарагиновой кислоты
За точку начала отсчета принимаем оксалоацетат, который образуется при трансаминировании из аспартата и считаем, что все должно закончиться также на этом уровне.
Оксалоацетат содержит 4 атома углерода и поэтому ему необходимо пройти 2 оборота ЦТК, прежде чем эти углероды выделятся в виде СО2. Подсчитывая число восстановленных НАДН, ФАДН2 и ГТФ, образуемых вдвух оборотах ЦТК, определяем конечную сумму АТФ – 24 молекулы.
При расчете коэффициента Р/О учитываем только фосфат, включенный в АТФ ферментом АТФ-синтазой, т.е. в процессе окислительного фосфорилирования. Это значит, что фосфат, входящий в ГТФ, не учитывается!
И, наконец, помним, что каждая молекула восстановленного эквивалента (т.е. НАДН или ФАДН2) передает на дыхательную цепь по одной паре электронов, которые проходят разное расстояние: от НАДН – трикомплекса ферментов, от ФАДН2 – два комплекса ферментов, но в любом случае восстанавливают до воды по одному атому кислорода. Иначе говоря, при окислении НАДН образуется 3 молекулы АТФ, при окислении ФАДН2 – молекулы АТФ.
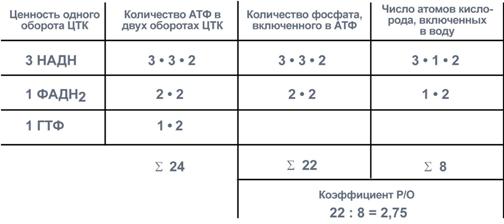
Соотношение количества АТФ, неорганического фосфата и кислорода при окислении аспартата
Окисление аланина
Два атома углерода аланина попадают в ЦТК после того, как безазотистый остаток аланина – пируват – окислится в пируват-дегидрогеназном комплексе с образованием НАДН. Образованный ацетил-S-КоА входит в ЦТК и полностью окисляется в одном его обороте. В общей сложности в дыхательную цепь направляется 4 молекулы НАДН (3 из ЦТК и 1 из ПВК-дегидрогеназы) и 1 молекула ФАДН2.
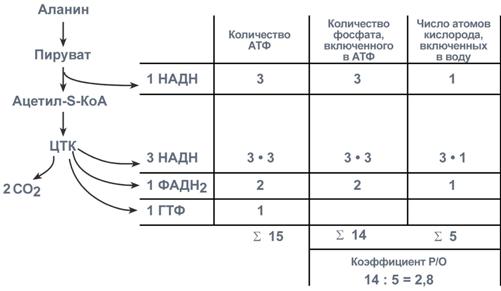
Соотношение количества АТФ, неорганического фосфата и кислорода при окислении аланина
Классифицируют гормоны по-разному
Гормоны (греч. hormao – привожу в движение) – это вещества, вырабатываемые специализированными клетками и регулирующие обмен веществ в отдельных органах и во всем организме в целом. Для всех гормонов характерна большая специфичность действия и высокая биологическая активность.
С нарушением гормонального обмена связан ряд наследственных и приобретенных заболеваний, сопровождающихся серьезными проблемами в развитии и жизнедеятельности организма (карликовость игигантизм, сахарный и несахарный диабет, микседема, бронзовая болезнь и др).
Гормоны можно классифицировать по химическому строению, растворимости, локализации их рецепторов и влиянию на обмен веществ.
Классификация по строению
Гормоны – производные аминокислот
Адреналин, норадреналин, тироксин, трийодтиронин.
Пептидные гормоны
Адренокортикотропный гормон (АКТГ), соматотропный гормон (СТГ), тиреотропный гормон (ТТГ), лактотропный гормон (пролактин, ПЛГ), лютеинизирующий гормон (ЛГ), фолликулостимулирующий гормон (ФСГ), меланоцитстимулирующий гормон (МСГ), антидиуретический гормон (АДГ, вазопрессин), окситоцин, кальцитонин, паратгормон, инсулин, глюкагон.
Стероидные гормоны
Кортизол, альдостерон, эстрадиол, прогестерон, тестостерон, кальцитриол.
Классификация по влиянию на обмен веществ
Белковый обмен
Инсулин, СТГ, АКТГ и кортизол, ТТГ и тироксин,
Липидно-углеводный обмен
Инсулин, СТГ, АКТГ и кортизол, ТТГ и тироксин, адреналин, глюкагон,
Водно-солевой обмен
Альдостерон, АДГ
Обмен кальция и фосфора
Кальцитонин, паратгормон, кальцитриол
Репродуктивная функция
Гонадотропные гормоны и эстрадиол, эстриол, прогестерон, тестостерон, пролактин, окситоцин
Классификация по месту синтеза
Гипоталамус
Кортиколиберин, тиреолиберин, гонадолиберин, соматолиберин, меланолиберин.
Пролактостатин, соматостатин, меланостатин.
Гипофиз
СТГ, АКТГ, ЛТГ, ТТГ, АДГ, МСГ, ФСГ, ЛГ, окситоцин.
Периферические железы
Инсулин, глюкагон, кортизол, тироксин, адреналин, альдостерон, эстрадиол, эстриол, тестостерон, кальцитонин, паратгормон, кальцитриол.
Гормональный сигнал должен пройти через мембрану
Для регуляции деятельности клетки с помощью гормонов, находящихся в плазме крови, необходимо обеспечить возможность клетки воспринимать и обрабатывать этот сигнал. Эта задача усложняется тем, что сигнальные молекулы (нейромедиаторы, гормоны, эйкозаноиды) имеют разную химическую природу, реакция клеток на сигналы должна быть различной по направленности и адекватной по величине.
В связи с этим, эволюционно сформировались два основных механизма действия сигнальных молекул по локализации рецептора:
1. Мембранный – рецептор расположен на мембране. Для этих рецепторов в зависимости от способа передачи гормонального сигнала в клетку выделяют три вида мембраносвязанных рецепторов и, соответственно, три механизма передачи сигнала. По данному механизму работают пептидные и белковые гормоны, катехоламины, эйкозаноиды.
2. Цитозольный – рецептор расположен в цитозоле.