

2 курс / Гистология / Основы_гистологии_УЧЕБНО_МЕТОДИЧЕСКОЕ_ПОСОБИЕ_1
.pdf2.Барьерно-защитная: защищают внутреннюю среду клетки от действия вредных внешних факторов.
3.Рецепторная (см. рис. 3.2).
4.Транспортная: транспорт веществ в клетку — эндоцитоз, и из клетки — экзоцитоз.
5.Участие в межклеточных взаимодействиях: формирование межклеточных контактов, дистантные взаимодействия между клетками.
Второй частью клеточной оболочки является гликокаликс (Рис. 3.2). Он представлен углеводными концами сложных белков (гликопротеинов) и сложных липидов (липопротеинов), входящих в состав цитомембраны. В гликокаликсе располагаются также поверхностные белки мембран, полуинтегральные белки. Их функциональные участки находятся в гликокаликсе. Эти белки могут играть роль ферментов. В гликокаликсе находятся рецепторы гистосовместимости, иммуноглобулины, могут адсорбироваться ферменты, рецепторы гормонов.
Функции гликокаликса: 1. Рецепторная (распознавание молекул соседних клеток и межклеточного вещества); 2. Межклеточные (адгезивные) контакты и взаимодействия; 3. Ориентация белков в мембране; 4. Участие в транспорте веществ.
Третий компонент клеточной оболочки — подмембранный слой опорно-сократительных структур. В его состав входят сократительные структуры — актиновые филаменты, а также опорный аппарат — кера-тиновые филаменты, микротрубочки. Подмембранный слой тесно
связан с цитоскелетом с одной стороны, и рецепторами гликокаликса — с другой.
Функции подмембранного слоя: поддержание формы клетки, создание ее упругости, изменения клеточной поверхности, за счет чего клетка участвует в эндо- и экзоцитозе, фагоцитозе, движении, секреции. С другой стороны, подмембранный слой связывает клеточную поверхность с компонентами цитоплазмы, поддерживает их упорядоченное расположение.
ПОНЯТИЕ О ЦИТОРЕЦЕПТОРАХ
Рецепторы представляют собой белковые молекулы на поверхности клетки, в ее цитоплазме или ядре, которые специфически реагируют с лигандами (гормонами, нейромедиаторами, факторами роста, цитокинами) или другими клетками. В соответствии со своей локализацией рецепторы делятся на поверхностные и внутриклеточные, а внутриклеточные подразделяются на цитоплазматические и ядерные.
Поверхностные рецепторы образованы поверхностными белками цито-мембран, а также гликокаликсом. Они предназначаются для полярных лиганд, т.е. веществ, которые не могут проникнуть через клеточную мембрану внутрь клетки и оказывают свое действие на нее через систему внешних рецепторов и вторичных посредников. Подразделяются на каталитические рецепторы, рецепторы, связанные с ионными каналами, рецепторы, связанные с G-белками, и рецепторы, связывающие молекулы внеклеточного матрикса с цитоскелетом.
Гликокаликс образует своеобразные "антенны", которые состоят из нескольких моно(олиго)сахаридных участков. Эти участки имеют разную конфигурацию, благодаря чему могут связываться с самыми различными химическими веществами. "Антенны" распознают различные внешние сигналы: молекулы гормонов, нейромедиаторов, факторов роста, цитокинов, генетически чуждые вещества и др. Рецепторные белки и углеводные участки часто связаны с
https://t.me/medicina_free
ферментами (каталитические рецепторы). Такие рецепторные белки являются трансмембранными и состоят из рецепторного и каталитического участков.
В качестве примера можно привести протеинкиназы (например, тирозин-киназа). Эти ферменты активируют внутриклеточные белки, что ведет к образованию второго посредника (мессенджера), передающего внешние сигналы в клетку, изменяя ее метаболизм, усиливая или ослабляя обмен веществ, синтез секрета. Так построены рецепторы инсулина, факторов роста и др.
Мембранные рецепторы могут изменять проницаемость мембран для ионов, что ведет к формированию электрического импульса (рецепторы к нейромедиаторам). Это так называемые рецепторы, связанные с ионными каналами. Рецепторы также контролируют поступление в клетку различных молекул, связывают молекулы внеклеточного матрикса с компонентами цитоске-лета
(рецепторы, связывающие молекулы внеклеточного матрикса с цитоскелетом). К таким рецепторам относят, например, интегрины. Они относятся к молекулам адгезии клеток (МАК). Интегрины — трансмембранные белки, воспринимающие молекулы внеклеточного матрикса, в частности, фибронектина и ламинина. В свою очередь, фибронек-тин связывается с другими молекулами внеклеточного матрикса (фибрином, коллагеном, гепарином и др.), а интегрин при помощи ряда других белков — с цитоскелетом. Таким образом, влияние молекул внеклеточного матрикса может передаваться на компоненты цитоскелета. Под влиянием раздражения этого вида рецепторов может изменяться состояние подмембранного слоя, и клетка может начать движение, а также экзоцитоз, эндоцитоз и другие виды деятельности. Особый вид поверхностных рецепторов — рецепторы, связанные с G-белками. Это трансмембранные белки, которые могут быть связаны либо с ионным каналом, либо с ферментом. Состоят из двух частей: рецепторной, взаимодействующей с сигнальными молекулами, и субъединиц G-белка ос, Р, у. G-белки — белки, связывающие гуанозинтрифосфат (ГТФ). После связывания с сигнальной молекулой комплекс G- белков передает сигнал на ассоциированный с цитолеммой фермент аденилатциклазу, которая синтезирует вторичный посредник циклический аденозинмонофосфат (цАМФ). В качестве вторичного посредника могут выступать и молекулы кальция. Через рецепторы, связанные G- белками, опосредуется действие на клетку подавляющего большинства гормонов и нейромедиаторов.
Внутриклеточные рецепторы находятся внутри клетки — в гиалоплазме, на мембранах органелл (цитоплазматические рецепторы), в ядре (ядерные рецепторы). Они предназначены для гормонов и других биологически активных веществ, которые в силу неполярности своих молекул могут легко проникать внутрь клетки (стероидные и тиреоидные гормоны и др.). Особый интерес представляют ядерные рецепторы. С этими рецепторами связываются такие гормоны, как стероидные, тиреоидные, витамин D,.
Молекулы таких рецепторов состоят из 2 участков: участок для связывания с гормоном и участок, взаимодействующий со специфическими участками ДНК в ядре. Ядерные рецепторы являются факторами транскрипции. Некоторые из них относятся к протоонкогенам — генам нормального генома, регулирующим пролиферацию клеток органов-мишеией, их дифференцировку и межклеточные взаимодействия. В результате соматических мутаций в протоонкогенах может происходить злокачественное перерождение клеток.
Внутриклеточные рецепторы могут находиться также на мембранах органелл, например, на митохондриях содержатся рецепторы к тиреоидным гормонам и т.д.
https://t.me/medicina_free

https://t.me/medicina_free
МОЛЕКУЛЫ АДГЕЗИИ КЛЕТОК (МАК)
С деятельностью поверхностных рецепторов клеток связан такой феномен, как клеточная адгезия.
Адгезия — процесс взаимодействия специфических гликопротеинов соприкасающихся
плазматических мембран распознающих друг друга клеток или клеток и внеклеточного матрикса. В том случае, если гликоиротеины при этом образуют связи, происходит адгезия, а затем формирование прочных межклеточных контактов или контактов клетки и межклеточного матрикса.
Все молекулы клеточной адгезии подразделяются на 5 классов.
1.Кадгерины. Это трансмембранные гликопротеины, использующие для адгезии ионы кальция. Отвечают за организацию цитоскелета, взаимодействие клеток с другими клетками.
2.Интегрины. Как уже отмечалось, интегрины представляют собой мембранные рецепторы для белковых молекул внеклеточного матрикса — фибронектина, ламинина и др. Связывают внеклеточный матрикс с цитос-келетом при помощи внутриклеточных белков талина, винкулина, а-акти-нина. Функционируют как клеточно-виеклеточные, так и межклеточные адгезионные молекулы.
3.Селектины. Обеспечивают прилипание лейкоцитов к эндотелию сосудов и тем самым — лейкоцитарно-эндотелиальпые взаимодействия, миграцию лейкоцитов через стенки сосудов в ткани.
4.Семейство иммуноглобулинов. Эти молекулы играют важную роль в иммунном ответе, а также в эмбриогенезе, заживлении ран и др.
5.Гоминговые молекулы. Обеспечивают взаимодействие лимфоцитов с эндотелием, их миграцию и заселение специфических зон иммунокомпс-тептных органов.
Таким образом, адгезия является важным звеном клеточной рецепции, играет большую роль в межклеточных взаимодействиях и взаимодействиях клеток с внеклеточным матриксом. Адгезионные процессы абсолютно необходимы при таких общебиологических процессах, как эмбриогенез, иммунный ответ, рост, регенерация и др. Они участвуют также в регуляции внутриклеточного и тканевого гомеостаза.
ЦИТОПЛАЗМА
ГИАЛОПЛАЗМА. Гиалоплазму называют также клеточным соком, цитозолем, или клеточным матриксом. Это основная часть цитоплазмы, составляющая около 55% объема клетки. В ней осуществляются основные клеточные обменные процессы. Гиалонлазма является сложной коллоидной системой и состоит из гомогенного мелкозернистого вещества с низ-кой электронной плотностью. Она состоит из воды, белков, нуклеиновых кислот, полисахаридов, липидов, неорганических веществ. Гиалоплазма может менять свое агрегатное состояние: переходить из состояния жидкого (золь) в более плотное — гель. При этом может изменяться форма клетки, ее подвижность и обмен веществ. Функции гиалонлазмы:
1.Метаболическая — метаболизм жиров, белков, углеводов.
2.Формирование жидкой микросреды (матрикса клетки).
https://t.me/medicina_free
3. Участие в движении клетки, обмене веществ и энергии. ОРГАНЕЛЛЫ. Органеллы — это второй важнейший обязательный
компонент клетки. Важным признаком органелл является то, что они имеют постоянное строго определенное строение и функции. По функциональному признаку все органеллы делятся на 2 группы:
1.Органеллы общего значения. Содержатся во всех клетках, поскольку необходимы для их жизнедеятельности. Такими органеллами являются: митохондрии, эндоплазматическая сеть (ЭПС) двух видов, комплекс Голь-джи (КГ), центриоли, рибосомы, лизосомы, пероксисомы, микротрубочки и микрофиламенты.
2.Органеллы специального значения. Есть только в тех клетках, которые выполняют специальные функции. Такими органеллами являются миофибриллы в мышечных волокнах и клетках, нейрофибриллы в нейронах, жгутики и реснички.
По структурному признаку все органеллы делятся на: 1) органеллы мембранного типа и 2)
органеллы немембранного типа. Кроме того, немембранные органеллы могут быть построены по
фибриллярному и гранулярному принципу.
В органеллах мембранного типа основным компонентом являются внутриклеточные мембраны. К таким органеллам относятся митохондрии, ЭПС, КГ, лизосомы, пероксисомы. К немембранным органеллам фибриллярного типа относятся микротрубочки, микрофиламенты, реснички, жгутики, центриоли. К немембранным гранулярным органеллам относят рибосомы, полисомы.
МЕМБРАННЫЕ ОРГАНЕЛЛЫ
ЭНДОПЛАЗМАТИЧЕСКАЯ СЕТЬ (ЭПС) - мембранная органелла, описанная в 1945 году К. Портером. Ее описание стало возможно благодаря электронному микроскопу. ЭПС — это система мелких каналов, вакуолей, мешочков, образующих в клетке непрерывную сложную сеть, элементы которой часто могут формировать кажущиеся на ультратонких срезах изолированными вакуоли. ЭПС построена из мембран, более тонких, чем цитолемма, и содержащих больше белка из-за находящихся в ней многочисленных ферментных систем. Есть 2 вида ЭПС: гранулярная (шероховатая) и агранулярная, или гладкая. Оба вида ЭПС могут взаимно переходить друг в друга и функционально связаны между собой так называемой переходной, или транзиторной, зоной.
Гранулярная ЭПС (рис. 3.3) содержит на своей поверхности рибосомы (полисомы) и является органеллой биосинтеза белка. Полисомы или рибосомы связываются с ЭПС при помощи так называемого причального белка (docking protein). При этом в мембране ЭПС имеются специальные интегральные белки рибофорины, также связывающие рибосомы и формирующие гидрофобные трапемембранные каналы для транспорта синтезированной полипентидной цени в просвет гранулярной ЭПС.
Гранулярная ЭПС видна только в электронном микроскопе. В световом микроскопе признаком развитой гранулярной ЭПС служит базофилия цитоплазмы. Гранулярная ЭПС имеется в каждой клетке, но степень ее развития различна. Она максимально развита в клетках, синтезирующих белок на экспорт, т.е. в секреторных клетках. Максимального развития гранулярная ЭПС достигает в нейроцитах, в которых ее цистерны приобретают упорядоченное расположение. В этом случае на светомикроскопическом уровне она выявляется в виде закономерно расположенных участков базофилии цитоплазмы, называемых базофилыюй субстанцией Ниссля.
https://t.me/medicina_free

Функция гранулярной ЭПС — синтез белка на экспорт. Кроме того, в ней происходят начальные посттрансляционные изменения полипептидной цепочки: гидроксилирование, сульфатирование и фосфорилиро-вание, гликозилирование. Последняя реакция особенно важна, т.к. приводит к образованию гликопротеинов — наиболее частого продукта клеточной секреции.
Агранулярная (гладкая) ЭПС представляет собой трехмерную сеть канальцев, не содержащих рибосомы. Гранулярная ЭПС может без перерыва переходить в гладкую ЭПС, но может существовать как самостоятельная органелла. Место перехода гранулярной ЭПС в агранулярную называется переходной (промежуточной, транзиторной) частью. От нее происходит отделение пузырьков с синтезированным белком и транспорт их к комплексу Гольджи.
Функции гладкой ЭПС:
1. Разделение цитоплазмы клетки на отделы — компартменты, в каждом из которых идет своя группа биохимических реакций.
https://t.me/medicina_free
2.Биосинтез жиров, углеводов.
3.Образование пероксисом;
4.Биосинтез стероидных гормонов;
5.Дезинтоксикация экзо- и эндогенных ядов, гормонов, биогенных аминов, лекарств за счет деятельности специальных ферментов.
6.Депонирование ионов кальция (в мышечных волокнах и миоцитах);
7.Источник мембран для восстановления кариолеммы в телофазе митоза.
ПЛАСТИНЧАТЫЙ КОМПЛЕКС ГОЛЬДЖИ. Это мембранная органелла, описанная в 1898 г. итальянским нейрогистологом К.Гольджи. Он назвал эту органеллу внутриклеточным сетчатым аппаратом из-за того, что в световом микроскопе она имеет сетчатый вид (рис. 3.4, а). Световая микроскопия не дает полного представления о строении этой органеллы. В световом микроскопе комплекс Гольджи имеет вид сложной сети, в которой ячейки могут быть связаны друг с другом или лежать независимо друг от друга (диктиосомы) в виде отдельных темных участков, палочек, зерен, вогнутых дисков. Между сетчатой и диффузной формой комплекса Гольд-жи нет принципиального различия, может наблюдаться смена форм этой оргамеллы. Еще в эпоху световой микроскопии было отмечено, что морфология комплекса Гольджи зависит от стадии секреторного цикла. Это позволило Д.Н.Насонову предположить, что комплекс Гольджи обеспечивает накопление синтезируемых веществ в клетке. По данным электронной микроскопии, комплекс Гольджи состоит из мембранных структур: плоских мембранных мешков с ампулярными расширениями на концах, а также крупных и мелких вакуолей (рис. 3.4, б, в). Совокупность этих образований называют диктиосомой. В диктиосоме находятся 5—10 мешковидных цистерн. Число диктиосом в клетке может достигать нескольких десятков. При этом каждая диктиосома связана с соседней при помощи вакуолей. В каждой диктиосоме есть проксимальная, незрелая, формирующаяся, или ЦИС-зона, -повернутая к ядру, и дистальная, ТРАНС-зона. Последняя, в отличие от выпуклой цис-поверхности, вогнутая, зрелая, обращена к цитолемме клетки. С цис-стороны происходит присоединение пузырьков, отделяющихся от переходной зоны ЭПС и содержащих вновь синтезированный и частично процессированный белок. При этом мембраны пузырьков встраиваются в мембрану цис-поверхности. С трансстороны отделяются секреторные пузырьки и лизосомы. Таким образом, в комплексе Гольджи существует постоянный поток клеточных мембран и их созревание. Функции комплекса Гольджи:
1.Накопление, созревание и конденсация продуктов биосинтеза белка (происходящего в гранулярной ЭПС).
2.Синтез полисахаридов и превращение простых белков в гликопротеины.
3.Образование липонротеидов.
4.Формирование секреторных включений и выделение их из клетки (упаковка и секреция).
5.Образование первичных лизосом.
6.Формирование клеточных мембран.
https://t.me/medicina_free
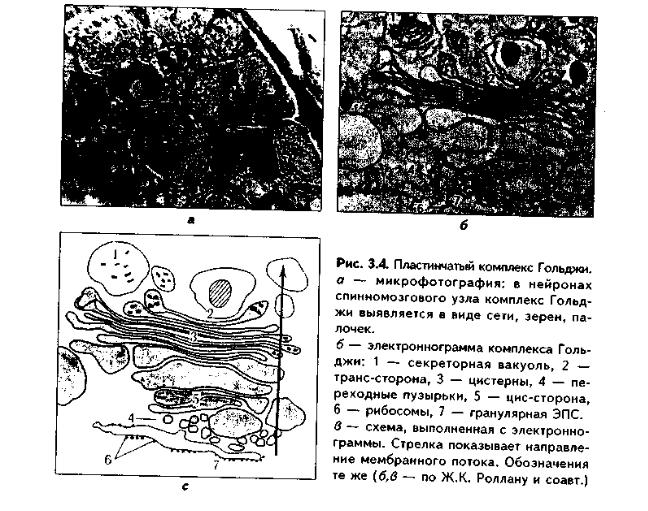
7. Образование акросомы — структуры, содержащей ферменты, находящейся на переднем конце сперматозоида и необходимой для оплодотворения яйцеклетки, разрушения ее оболочек.
МИТОХОНДРИИ. Эти органеллы обеспечивают окисление органических соединений и синтез АТФ. Были открыты в 1890 году немецким ученым Р. Альтманом при помощи предложенного им метода окрашивания кислым фуксином, при этом выглядели в виде нитей и зерен (отсюда их название). Интересно, что сам Р. Альтман считал митохондрии бактериями, внедрившимися в клетку с паразитической целью, а в последующем ставшими симбионтами (симбиотическая теория происхождения митохондрий в настоящее время поддерживается многими исследователями).
Размеры митохондрий составляют от 0,5 до 7 мкм, а их общее число в клетке — от 50 до 5000. Эти органеллы хорошо видны в световом микроскопе, однако информация об их строении, получаемая при этом, скудна (рис. 3.5, а). Электронный микроскоп показал, что митохондрии состоят из двух мембран — наружной и внутренней, каждая из которых имеет толщину 7 нм (рис. 3.5, б, в, 3.6, а). Между наружной и внутренней мембранами имеется щель размером до 20 нм.
Внутренняя мембрана неровная, образует много складок, или крист. Эти кристы идут перпендикулярно поверхности митохондрии. На поверхности крист имеются грибовидные образования (оксисомы, АТФсомы или F,-частицы), представляющие собой АТФ-синтетазный
https://t.me/medicina_free
комплекс (рис. 3.6) Внутренняя мембрана отграничивает матрикс митохондрии. Он содержит многочисленные ферменты для окисления пирувата и жирных кислот, а также ферменты цикла Кребса. Кроме того, в матриксе находятся мито-хондриальная ДНК, митохондриальные рибосомы, т-РНК и ферменты активации митохондриального генома. Внутренняя мембрана содержит белки трех типов: ферменты, катализирующие окислительные реакции; АТФ-син-тезатный комплекс, синтезирующий в матриксе АТФ; транспортные белки. Наружная мембрана содержит ферменты, превращающие липиды в реакционные соединения, участвующие затем в метаболических процессах матрикса. Межмембранное пространство содержит ферменты, необходимые для окислительного фосфорилирования. Т.к. митохондрии имеют свой геном, то они обладают автономной системой синтеза белка и могут частично строить собственные белки мембран.
Функции.
1.Обеспечение клетки энергией в виде АТФ.
2.Участие в биосинтезе стероидных гормонов (некоторые звенья биосинтеза этих гормонов протекают в митохондриях). Клетки-продуценты сте
роидных гормонов имеют крупные митохондрии со сложными крупными трубчатыми кристами.
3.Депонирование кальция.
4.Участие в синтезе нуклеиновых кислот. В некоторых случаях в результате мутаций митохондриалыюй ДНК возникают так называемые митохондриальные болезни, проявляющиеся широкой и тяжелой симптоматикой. ЛИЗОСОМЫ. Это мембранные органеллы, не видимые в световом микроскопе. Были открыты в 1955 году К. де Дювом при помощи электронного микроскопа (рис. 3.7). Представляют собой мембранные пузырьки, содержащие гидролитические ферменты: кислую фосфатазу, липазу, протеазы, нуклеазы и др., всего более 50 ферментов. Различают лизосомы 5 типов:
1.Первичные лизосомы, только что отделившиеся от транс-поверхности комплекса Гольджи.
2.Вторичные лизосомы, или фаголизосомы. Это лизосомы, которые соединились с фагосомой
— фагоцитированной частицей, окруженной мембраной.
3.Остаточные тельца — это слоистые образования, формирующиеся в том случае, если процесс расщепления фагоцитированных частиц прошел не до конца. Примером остаточных телец могут быть липофусциновые включения, которые появляются в некоторых клетках при их старении, содержат эндогенный пигмент липофусцин.
4.Первичные лизосомы могут сливаться с погибающими и старыми органеллами, которые они разрушают. Такие лизосомы называются ауто-фагосомами.
5.Мультивезикулярные тельца. Представляют собой крупную вакуоль, в которой, в свою очередь, находятся несколько так называемых внутренних пузырьков. Внутренние пузырьки, очевидно, образуются путем отпочковывания внутрь от мембраны вакуоли. Внутренние пузырьки могут постепенно растворяться содержащимися в матриксе тельца ферментами.
Функции лизосом: 1. Внутриклеточное пищеварение. 2. Участие в фагоцитозе. 3. Участие в митозе — разрушении ядерной оболочки. 4. Участие во внутриклеточной регенерации.5. Участие в аутолизс — саморазрушении клетки после ее гибели.
https://t.me/medicina_free

Существует большая группа болезней, называемых лизосомными болезнями, или болезнями накопления. Они являются наследственными болезнями, проявляются дефицитом определенного лизосомального пигмента. При этом в цитоплазме клетки накапливаются непереваренные продукты
обмена веществ (гликоген, гликолиниды, белки, рис. 3.7, б,в), что ведет к постепенной гибели клетки. ПЕРОКСИСОМЫ. Пероксисомы — это оргаиеллы, напоминающие ли-зосомы, но содержащие ферменты, необходимые для синтеза и разрушения эндогенных перекисей — нероксидазу, каталазу и другие, всего до 15. В электронном микроскопе представляют сферические или эллипсоидные пузырьки с умеренно плотной сердцевиной (рис. 3.8). Образуются пероксисомы путем отделения пузырьков от гладкой ЭПС. В эти пузырьки затем миг-рируют ферменты, которые синтезируются отдельно в цитозоле или в гранулярной ЭПС
https://t.me/medicina_free