

Глава I
АНАТОМИЯ И ФИЗИОЛОГИЯ ОРГАНОВ РАЗМНОЖЕНИЯ
ОПРЕДЕЛЕНИЕ И ДИФФЕРЕНЦИРОВКА ПОЛА
Одним из основных свойств всех живых существ является их способность к размножению. Способ размножения зависит от степени совершенства вида. Для многих групп организмов характерно размножение при помощи отдельных клеток, или цитогония.
Наиболее распространенная форма цитогонии у млекопитающих - половое размножение (гамогония или амфигония), - характеризующаяся образованием дифференцированных половых клеток (гамет) и их последующим соединением в зиготу в процессе оплодотворения. Дифференциация гамет определяется существованием двух полов - мужского и женского. У каждого пола свои специфические железы (гонады), которые продуцируют не только половые клетки, но и особые вещества внутренней секреции - гормоны. Женские гаметы - ооциты (яйцеклетки) - крупные, богаты питательным материалом, неподвижные. У многих видов они незначительно отличаются по своей структуре и в то же время сильно различаются по величине, что связано с различной степенью накопления питательного материала. Мужские гаметы - спермин (сперматозоиды) - имеют меньшую величину, состоят почти исключительно из ядра и обладают подвижностью, которая позволяет им в соответствующей среде достигнуть яйцеклетки, проникнуть внутрь и оплодотворить ее.
В результате оплодотворения образуется зигота. Хромосомный набор ее диплоидный, в отличие от гаплоидного у обеих половых клеток. Такой хромосомный набор у гамет создается благодаря редукционному делению (мейозу), который происходит в семеннике и яичнике во время фазы созревания. Таким образом, два важнейших процесса: оплодотворение (сингамия) и мейоз взаимно дополняют друг друга и делают возможным наследование родительских признаков.
После оплодотворения из зиготы развивается новый организм мужского или женского пола. В формировании полов следует выделить два этапа - определение и дифференцировку пола.
Определение пола – это механизм, по которому распределяются половые хромосомы согласно первому закону Менделя. У животных решающим фактором в определении пола является спермий. Если яйцеклетка будет оплодотворена спермием, несущим Х-половую хромосому, то зигота получит две такие хромосомы и из нее станет развиваться самка. При участии в оплодотворении спермия с У-половой хромосомой зигота получит X и Y хромосомы, что обусловит развитие самца. Определение пола заканчивается при оплодотворении.
Дифференцировка пола - это цепь процессов развития, в результате которых генетически обусловленный пол находит свое выражение в виде характерных структур, функций, систем поведения. Дифференцировка происходит на протяжении всего внутриутробного периода и в первое время после рождения.
Следует отметить, что половой аппарат самцов и самок развивается из полового зачатка, одинакового у эмбрионов обоих полов. В состав зачатка входят две недифференцированные (нейтральные) гонады, которые находятся в верхней части брюшной полости, две первичные почки с их выделительными Волъфовыми протоками и два Мюллеровых протока. Первичные почки расположены рядом с гонадами (рис. 1).

В нейтральные гонады на ранних стадиях развития внедряются первичные половые клетки. Они представляют собой малодифференцированные клетки, которые вначале размещаются в области внезародышевой энтодермы, а на более поздних стадиях развития мигрируют по кровеносным сосудам или перемещаются в мезенхиме и принимают участие в образовании зачатков гонад. Здесь первичные половые клетки вступают в контакт с опорными (фолликулярными) клетками, которые снабжают их питательными веществами. Вокруг каждой половой клетки в яичнике фолликулярные клетки формируют эпителиальный пузырек (фолликул), а в семеннике они вместе с половыми клетками образуют стенку семенных канальцев.
Эндокринная функция зачатков половых желез начинается до морфологической дифференцировки. У крупного рогатого скота прогестерон синтезируется 13-14-дневными бластоцистами и затем недифференцированными железами обоих полов. У эмбрионов с полуторамесячного возраста половые железы дифференцируются морфологически и функционально и продуцируют тестостерон или 17β-эстрадиол. У эмбрионов свиней дифференцировка половых желез начинается с 27-дневного возраста и завершается к 30-му дню.
Дифференцировка половых органов самцов. Только пройдя нейтральную стадию развития половых зачатков, наследственная половая конституция зародыша начинает проявлять заметное влияние на половую дифференциацию. У эмбриона, имеющего половые хромосомы XY, внутренняя (медулярная) часть гонады расширяется и дифференцируется, корковая часть почти исчезает и нейтральная часть гонады превращается в семенник. Остатки корковой части сморщиваются и преобразуются в систему связок, которые прикрепляются на передней и задней части семенников. Задняя (направляющая) связка соединяет гонаду с брюшной стенкой. Начальный участок связки (до Вольфова протока) превращается в специальную связку, конечный участок — в паховую связку. Остатки первичных почек дают начало придаткам семенников, а Вольфовы протоки преобразуются в спермиопроводы. Позднее в каждом спермиопроводе происходит выпячивание стенки, т. е. образуется семенной пузырек.
Мюллеровы каналы дегенерируют. Но нередко нижняя часть их остается между ампулами спермиопрово-дов в виде тяжа или кистоподобного образования, а верхняя часть прикрепляется к семеннику.
В ходе эмбрионального развития паховая связка семенника сжимается, укорачивается и оттягивает семенник вместе с серозной оболочкой вниз в брюшную полость и через отверстие в брюшной стенке (паховый канал) – в мошонку.
У различных видов животных опускание семенников происходит в различные периоды развития. У собак при рождении семенники находятся в брюшной полости и опускаются в мошонку через 5 дней. У хрячков опускание семенников начинается при длине туловища более 20 см и завершается к моменту рождения, иногда за 20-25 дней до него. У жвачных этот процесс происходит на более ранних стадиях внутриутробного развития: у баранчиков он заканчивается к 80-му дню, у бычков - в 3-3,5 мес. Андрогены ускоряют опускание семенников в мошонку, эстрогены задерживают этот процесс. У кобыл на 5-6-м мес. беременности отмечается повышенное содержание в крови эстрогенов и заметное увеличение половых желез у плодов. Очевидно, поэтому у жеребчиков опускание семенников происходит только к моменту рождения. Если семенники остаются в брюшной полости или паховом канале, развитие их замедляется, сперматогенез не может осуществляться полностью, и это приводит к уродству – крипторхизму. В зависимости от месторасположения семенников различают абдоминальных и инвагинальных крипторхов. Эта патология чаще встречается у лошадей и свиней, реже – у жвачных.
В процессе дифференцировки половых органов самца соединение семенника и придатка семенника, формирующихся из нейтральной гонады и первичной почки, происходит за счет прорастания соединительнотканных клеточных тяжей. Эти тяжи растут навстречу друг другу и постепенно становятся полыми. Они соединяются в единую систему, состоящую из извитых и прямых канальцев, сети семенника и семявыносящих канальцев. Если в тяжах образование каналов не происходит, это может послужить причиной застоя спермиев. Причиной этому может явиться и обеднение кислородом тканей придатка семенника вследствие замедления тока крови, что приводит к повреждению эпителия семявыносящих канальцев, парализует движение спермиев и вызывает образование очагов застоя спермиев. В конечном итоге вся система канальцев становится непроходимой.
Дифференцировка половых органов самок. У эмбриона с половыми хромосомами XX корковая часть гонады развивается очень сильно, медулярная часть становится малозаметной и нейтральная гонада превращается в яичник. Мюллеровы каналы преобразуются в трубчатые половые органы: яйцепроводы, рога, тело и шейку матки и влагалище. Вольфовы каналы дегенерируют. Остатки их у половозрелого животного находятся между яичником и яйцепроводом. Очень часто, особенно у крупного рогатого скота, в стенке влагалища встречаются рудименты дистальных отделов вольфовых протоков – Гартнеровы ходы. Наружные отверстия их находятся в преддверии влагалища латерально от отверстия мочеиспускательного канала.
Параллельно с внутренними из специального зачатка развиваются наружные половые органы. Зачаток возникает из элементов мезобласта и состоит из полового бугорка (возвышение возле клоачного отверстия) и полового валика (выпуклость мезодермы вокруг полового бугорка). Внутри полового валика расположены две половые складки, между которыми открывается отверстие мочеполового синуса (мочеиспускательного канала). Мюллеровы и Вольфовы каналы у зародышей сначала открываются в клоачный отрезок прямой кишки. В последующем в результате продвижения мезенхимы в пространство между прямой кишкой и мочеполовым синусом клоака разделяется на прямую кишку и мочеполовой синус. Половой бугорок у самцов превращается в половой член, у самок – в клитор, а из половых валиков образуется соответственно кожа мошонки или вульва.
Дифференцировка половых органов у самок различных видов происходит по-разному. У птиц правый Мюллеров канал и нейтральная гонада атрофируются. Левый канал преобразуется в примитивную матку и влагалище, которое открывается в прямую кишку. Вместе с ней влагалище образует клоаку. У млекопитающих в дифференцировке участвует весь половой зачаток.
У грызунов Мюллеровы каналы образуют два яйцепровода, две матки с двумя шейками, а задние участки каналов сливаются в общее влагалище. Матка грызунов относится к разновидности двойных – uterus duplex.
У крупного и мелкого рогатого скота, буйволиц и всеядных Мюллеровы каналы сливаются на большем протяжении, образуя влагалище, небольшой величины тело матки и два рога. Основания рогов матки сращены между собой на более или менее значительном расстоянии. Такая матка называется двураздельной – лuterus biparticus. У кобылы и ослицы основания рогов матки сливаются в единую полость (тело матки) и только их передние участки раздваиваются, образуя рога матки. Такую матку рассматривают как типично двурогую – uterus bicornis.
У приматов сращение Мюллеровых каналов еще более полное. Матка представляет собой единый мускульный мешок, имеющий самостоятельные ответвления Мюллеровых каналов в виде маточных труб – яйцепро-водов. Это простая матка – uterus simplex.
В отдельных случаях необходимое сращение
мюллеровых каналов не происходит, и тип
матки оказывается нехарактерным для
данного вида. Так, вместо простой матки
у женщины образуется двурогая матка, а
у животных с двурогой маткой развивается
двойная матка с двумя шейками, которые
открываются в общее влагалище, а иногда
- и в разделенное в передней части
вертикальной перегородкой. Такие
аномалии встречаются у всех
сельскохозяйственных животных. У
крупного рогатого скота степень сращения
Мюллеровых каналов сильно различается
и это приводит к развитию следующих
аномалий (рис. 2, а, б, в, г):
отдельных случаях необходимое сращение
мюллеровых каналов не происходит, и тип
матки оказывается нехарактерным для
данного вида. Так, вместо простой матки
у женщины образуется двурогая матка, а
у животных с двурогой маткой развивается
двойная матка с двумя шейками, которые
открываются в общее влагалище, а иногда
- и в разделенное в передней части
вертикальной перегородкой. Такие
аномалии встречаются у всех
сельскохозяйственных животных. У
крупного рогатого скота степень сращения
Мюллеровых каналов сильно различается
и это приводит к развитию следующих
аномалий (рис. 2, а, б, в, г):
двойная матка (uterus didelphys) - два яйцепровода, два маточных рога, два тела и две шейки матки;
двойная шейка матки (complete double cervix) - два яйцепровода, два маточных рога, одно тело матки и две шейки;
неполная двойная шейка матки (incomplete double cervix) -два яйцепровода, два маточных рога, одно тело матки, один цервикальный канал в передней части и два канала в задней части шейки; иногда один канал слепо заканчивается, но чаще, спустя одну треть длины шейки, сливается в общий канал;
двойное наружное отверстие (double os uteri externum) -два яйцепровода, два маточных рога, одно тело и одна шейка матки с вертикально расположенной полоской ткани у наружного отверстия; иногда в передней стенке влагалища толщиной 3—4 мм на расстоянии 4-6 см друг от друга находятся два отверстия; за стенкой имеется сплошное пространство и шейка матки начинается без характерного для нее выступа.
Частота удвоения шейки матки у коров достигает 3,7% (Ван Лоэн). Аномалия обусловлена генетическими факторами и не снижает существенным образом плодовитости таких особей. Число осеменений на стельность у них несколько выше (1,75), чем у нормальных животных (1,59), что может быть связано с трудностью попадания спермиев в соответствующий рог матки.
У кобыл также наблюдаются случаи полного
разделения матки и две шейки матки.
Плодовитость таких животных не нарушена.
кобыл также наблюдаются случаи полного
разделения матки и две шейки матки.
Плодовитость таких животных не нарушена.
Мочеполовой клапан эмбриологически является остатком раздельного существования Мюллеровых каналов. Недоразвитие этих каналов отмечается не так часто, как их раздельное существование, и это приводит к серьезным аномалиям половых органов. Случаи полного недоразвития протоков (аплазии) редки. У телок при клиническом исследовании вместо матки находят два тяжа толщиной с вязальную спицу. При частичной (сегментальной) аплазии отсутствует какой-либо участок половых путей (рис. 3). Чаще это проявляется отсутствием одного из рогов матки. Обнаруживается у свиней и крупного рогатого скота, более часто у телок шортгорнской породы (болезнь белых телок). Выявляются телки (редко – коровы) с однорогой маткой и среди животных черно-пестрой породы.
У таких особей только один рог матки имеет полость, а второй очень узкий, в виде ленты с отложением жировой ткани. Другие органы нормальные и животные могут оплодотвориться в случае овуляции в яичнике на стороне нормального рога матки. Иногда рог матки с полостью очень короткий или же отсутствует какой-либо его участок. При недоразвитии обоих рогов матки особи бесплодны.
АНАТОМИЯ И ТОПОГРАФИЯ ПОЛОВЫХ ОРГАНОВ САМОК
Половые органы самок вырабатывают женские половые клетки, необходимые для возникновения нового организма, и являются средой, в которой новый организм развивается в первые дни и месяцы своей жизни. Различают половые органы внутренние и наружные. К внутренним органам (genitalia interna) относят яичники, яйцепроводы, матку и влагалище; к наружным (vulva) – половые губы, преддверие влагалища и клитор.
В зависимости от вида животных и физиологического состояния внутренние половые органы расположены в тазовой или брюшной полости. Удерживаются они с помощью широких маточных связок. Связки представляют собой удвоенную брюшину, которая идет от поясничной области. В связках проходят сосуды и нервы.
Яичники, яйцепроводы и матка снабжаются кровью парных передней, средней и задней маточных артерий. Передняя маточная артерия отходит от аорты, средняя – от подвздошной у кобыл и от пупочной – у коров, задняя – от геморроидальной у кобыл и от мочеполовой у коров. Отток крови осуществляется по одноименным венам. У овец средние маточные вены отсутствуют, а кровь оттекает по передним пузырным и задним маточным венам.
Иннервируются половые органы симпатическими и парасимпатическими нервными стволами. У кобыл, коров, свиней и сук источником симпатических нервных стволов, идущих к половым органам, является каудальный брыжеечный узел. Он соединен со спинномозговыми нервами большими соединительными ветвями, и, следовательно, через спинной мозг поясничной области и через нервные сплетения брюшной полости проводящими путями – с центральными отделами нервной системы. Парасимпатические нервные ветви отходят от крестцовых нервов.
Лимфатическая система гениталий представлена капиллярами, которые залегают во всех оболочках половых органов, интраорганными и афферентными экстраорганными сосудами, крупнопетлистыми сосудистыми сплетениями, расположенными в маточных связках, крупными магистральными афферентными экстраорганными сосудами, впадающими в региональные лимфатические узлы, а также афферентными лимфатическими сосудами и протоками, которые соединяются и впадают в поясничный проток.
ПОЛОВЫЕ ОРГАНЫ КОРОВ И ТЕЛОК
Половые губы (labium pudendi) представляют собой два валиковидных выпячивания, расположенных над седалищными буграми. Смыкаясь, половые губы образуют верхний и нижний углы половой щели. Верхний угол закругленный, нижний - острый, снабжен пучком длинных волос. Наружная поверхность половых губ покрыта нежной пигментированной или непигментированной кожей, в толще которой имеются потовые и сальные железы. Внутренняя поверхность покрыта плоским многослойным эпителием. В толще губ находятся мышечные волокна суживателя половой щели и соединительная ткань. В верхней части волокна переходят в промежность и сливаются со сфинктером ануса, внизу окружают клитор, впереди продолжаются как суживатель мочеполового преддверия (рис. 4).

Клитор расположен внизу преддверия влагалища. Начинается в виде двух пещеристых тел на седалищных буграх. Вместе они образуют тело клитора, которое покрыто плотной фиброзного типа оболочкой. Заканчивается клитор заостренной головкой в нижнем углу половой щели. По бокам головки расположены складки слизистой оболочки, которые переходят на тело клитора и оканчиваются возле головки уздечкой.
Преддверие влагалища (vestibulum vaginae) - представляет собой узкий участок вагинальной трубки длиной 4-8 см. В норме оно сжато с боков и на разрезе представляет вертикальную щель. От половой щели по направлению к влагалищу канал преддверия направлен снизу вверх, вперед. На границе с влагалищем открывается отверстие мочеиспускательного канала (ostium urethraeexternum), нижняя стенка которого имеет слепой мешок. По бокам от отверстия и сзади от него находятся выводные протоки Бартолиниевых желез. По форме и величине они напоминают бульбо-уретральные железы быка. Степень развития их неодинакова, иногда они представлены островками ткани. На нижней стенке преддверия возле клитора расположены отверстия слабо развитых малых желез преддверия. Стенка преддверия влагалища состоит из слизистой, мышечной и соединительнотканной оболочек. Мышечная оболочка представлена, главным образом, гладкими мускульными волокнами, но в заднем участке его имеется значительное количество пучков поперечно-полосатых мышц. В боковой стенке преддверия ближе к половой щели расположено венозное сплетение, напоминающее пещеристое тело уретры самцов. Наружная оболочка преддверия состоит из прослоек рыхлой соединительной ткани, которая переходит в ткань промежности и прямой кишки.
Между анусом и половой щелью находится промежность, представленная рыхлой соединительной тканью. Она простирается вглубь между прямой кишкой и вагинальной трубкой и сходит на клин. От анального отверстия до верхнего угла половой щели кожа промежности образует небольшое валиковидное возвышение - шов промежности. Он хорошо выражен у молодых животных.
Влагалище (vaginae, colpos) – дистальная часть внутренних половых органов, представляет собой широкую мясистую трубку длиной 8-22 см. Начинается от шейки матки и сзади переходит в более узкое преддверие влагалища. Расположено влагалище в тазу и является органом совокупления. В норме влагалище сжато сверху вниз и на разрезе представляет собой горизонтальную щель. Передняя часть влагалища снаружи покрыта серозной оболочкой. Простирается она по верней стенке на 7-12 см и затем с влагалища переходит на прямую кишку, образуя в этом месте глубокое вагино-ректальное углубление брюшины. Внизу влагалище почти по всей длине прочно сращено с мочеиспускательным каналом. Серозная оболочка покрывает только небольшую часть стенки влагалища, переходит на мочевой пузырь и в этом месте образует слабо заметное влагалищно-пузырное углубление в брюшине. С боков между стенками влагалища и каналом таза имеется рыхлая соединительная ткань. Мышечная оболочка влагалища состоит из двух слоев гладких мышц: наружного слоя продольных волокон и внутреннего слоя, представленного поперечными волокнами. Оба слоя богаты эластическими волокнами. Слизистая влагалища покрыта плоским многослойным эпителием и не имеет желез. Она образует большое количество продольных глубоких и слабо выраженных поперечных складок. Наличие таких складок наряду с эластичностью мускульной оболочки обеспечивает высокую растяжимость стенок влагалища во время полового акта и в период родов. Растяжимость ограничена лишь стенками тазовой полости.
Возле шейки матки слизистая влагалища состоит из многослойных, вырабатывающих слизь клеток и тонких эпителиальных клеток. Интенсивное образование слизи в этой части влагалища наблюдается в период течки и охоты. По направлению к мочеполовому преддверию число слизеобразующих клеток резко уменьшается. В этом участке слизистая оболочка толстая и ее поверхностный эпителий может ороговевать. У телок в месте перехода влагалища в преддверие влагалища отмечается сужение полового канала. Примерно на 1 см впереди отверстия мочеиспускательного канала находится перегородка – мочеполовой клапан. Перегородка имеет вид связки различной ширины и толщины. У взрослых телок толщина перегородки варьирует от 1 до 2,5 мм, ширина – от 2 до 4-6 мм. В месте прикрепления перегородки к верхней и нижней стенкам влагалища ширина ее значительно увеличивается. Здесь она плавно переходит в продольные складки влагалища. Высота перегородки не превышает 10 мм (рис. 4).
Впереди влагалище ампуловидно расширяется и обхватывает со всех сторон влагалищную часть шейки матки. Здесь между шейкой матки и стенками влагалища образуются боковые, верхний и нижний карманы. Наиболее хорошо выраженный верхний карман называется сводом влагалища.
В стенке влагалища по бокам от отверстия мочеиспускательного канала в краниальном направлении у многих животных идут два слепо заканчивающихся канала – остатки вольфовых протоков (Гартнеровы ходы). Нередко имеется только один канал.
М атка
(uterus, metra) – служит вместилищем для
плода, обеспечивает его рост и развитие,
а затем - и выведение через родовые пути.
У телок и небеременных молодых коров
вся или почти вся матка находится в
тазовой полости, в ложбине лонного
сращения. У многорожавших коров в
расслабленном состоянии она частично
смещается в брюшную полость. Шейка
матки длиной 5-10 см, толщиной 3-4,5 см,
обычно расположена в тазу и лишь в
середине беременности перемещается в
брюшную полость. Она плотная, как бы
хрящеватая, с толстыми стенками, четко
ограничена и хорошо прощупывается рукой
через прямую кишку. Задняя часть шейки
матки на 2—3 см вдается во влагалище,
образуя ясно выраженную влагалищную
часть, в которой находится наружное
отверстие – устье шейки матки (ostium
uteri). Тело матки сравнительно
мягкое, длиной 2-5 см, впереди разделяется
на два рога. На протяжении 10-15 см рога
сращены между собой. В этом месте между
ними снаружи хорошо заметна межроговая
борозда (желоб). После раздвоения
(бифуркации) рога расходятся несколько
в стороны, затем загибаются вниз и назад,
а конечная часть их приподнимается
вверх к яичникам и переходит в яйцепроводы.
В месте расхождения рога имеют диаметр
2-3,5 см, но по направлению к верхушке
сильно истончаются. От точки разделения
они имеют длину 15-20 см. Общая длина
каждого рога – 30-55 см. У телок длина
рогов матки изменяется от 15 см в возрасте
12 мес. до 30 см в 2 года. С возрастом толщина
стенок матки и ее мышечной оболочки
имеет тенденцию к увеличению.
атка
(uterus, metra) – служит вместилищем для
плода, обеспечивает его рост и развитие,
а затем - и выведение через родовые пути.
У телок и небеременных молодых коров
вся или почти вся матка находится в
тазовой полости, в ложбине лонного
сращения. У многорожавших коров в
расслабленном состоянии она частично
смещается в брюшную полость. Шейка
матки длиной 5-10 см, толщиной 3-4,5 см,
обычно расположена в тазу и лишь в
середине беременности перемещается в
брюшную полость. Она плотная, как бы
хрящеватая, с толстыми стенками, четко
ограничена и хорошо прощупывается рукой
через прямую кишку. Задняя часть шейки
матки на 2—3 см вдается во влагалище,
образуя ясно выраженную влагалищную
часть, в которой находится наружное
отверстие – устье шейки матки (ostium
uteri). Тело матки сравнительно
мягкое, длиной 2-5 см, впереди разделяется
на два рога. На протяжении 10-15 см рога
сращены между собой. В этом месте между
ними снаружи хорошо заметна межроговая
борозда (желоб). После раздвоения
(бифуркации) рога расходятся несколько
в стороны, затем загибаются вниз и назад,
а конечная часть их приподнимается
вверх к яичникам и переходит в яйцепроводы.
В месте расхождения рога имеют диаметр
2-3,5 см, но по направлению к верхушке
сильно истончаются. От точки разделения
они имеют длину 15-20 см. Общая длина
каждого рога – 30-55 см. У телок длина
рогов матки изменяется от 15 см в возрасте
12 мес. до 30 см в 2 года. С возрастом толщина
стенок матки и ее мышечной оболочки
имеет тенденцию к увеличению.
Стенка матки состоит из трех оболочек: серозной, мышечной и слизистой (рис. 5). Серозная оболочка (perimetrium) покрывает матку снаружи. С боков тела и шейки и по малой кривизне рогов матки серозная оболочка переходит в широкие маточные связки, на которых она и подвешена. Мышечная оболочка (myometrium) состоит из трех слоев гладких мышц. Один слой наружных продольных волокон отделен сосудистым слоем от двух других слоев циркулярных и продольных волокон. В области шейки матки мышечные слои толще, особенно циркулярный слой. Слизистая оболочка матки (endometrium) от бледно-розового до буровато-красного цвета имеет
трубчатые маточные железы, которых насчитывается около 1 млн.; изнутри покрыта псевдомногорядным призматическим реснитчатым эпителием. В области тела и рогов матки в эндометрии имеются особые образования – карункулы. В каждом роге 4 ряда карункулов, по 10-14 в ряду, всего 80-120. Они выпуклы, длиной 15-17 мм, шириной 6-9 мм и 2-4 мм в высоту. При одноплодной беременности число карункулов колеблется от 45 до 144, при двуплодной – достигает 170. Во время беременности железистая часть карункулов разрастается, в ней образуются углубления, в которые внедряются ворсинки хориона. У небеременных животных карункулы на своей поверхности не имеют маточных желез.
Слизистая оболочка шейки матки бледная, образует многочисленные мелкие продольные и толстые поперечные складки или кольца. Противоположные складки заходят одна за другую, образуя извилистый церви-калъный канал, который открывается отверстием в тело матки. В области влагалищной части шейки мощно развитые поперечные складки слизистой оболочки (верхняя и нижняя) образуют отчетливо выраженную розетку. У молодых животных складки небольшие, ровные, у старых коров они могут быть сильно гипертрофированы и по форме напоминать цветную капусту. Поверхностный эпителий слизистой оболочки шейки матки имеет многочисленные слизеобразующие (бокаловидные) клетки. Они постоянно выделяют слизь, которая закупоривает цервикальный канал. Количество и консистенция этой слизи в различные фазы полового цикла и в период беременности различны.
Цервикальный канал, соединяющий полость матки с влагалищем, у нерожавших животных плотно закрыт и только во время течки происходит его расслабление, что позволяет провести через него пипетку диаметром 5-7 мм. У многорожавших животных полное закрытие канала не происходит и такой толщины пипетку можно провести через шейку матки во все периоды полового цикла.
Яйцепроводы (oviductus, tuba uterina, salpinx) – две извитые трубочки, служат местом оплодотворения яйцеклеток и.обеспечивают проведение их в рога матки. Располагаются яйцепроводы в складках брюшины (mesosalpinx), простирающихся от верхушки рогов матки до яичника. Канал яйцепровода начинается от верхушки соответствующего рога матки узким маточным отверстием. В зависимости от возраста животных длина яйцепроводов колеблется от 20 до 35 см. Наименьший диаметр (1-3 мм) яйцепроводы имеют в области перешейка (istmus), который примыкает к рогу матки и составляет около половины всей длины. Средняя часть –ампула, имеет диаметр 3-5 мм, конечная часть – воронка – 5-7 мм. Воронка открывается отверстием в брюшную полость. Свободный длинный край воронки называют бахромкой (fimbriae). Благодаря своеобразному расположению яичника в яичниковом кармане, в складке которого проходит яйцепровод, бахромка тесно прилегает к поверхности яичника, и это обеспечивает попадание овулировавших яиц в отверстие яйцепрово-да. Стенка яйцепровода состоит из трех оболочек: серозной, мышечной и слизистой. Серозная оболочка представлена в основном эпителиальным слоем брюшины и двумя слоями широкой маточной связки. Мышечная оболочка состоит из трех слоев гладких мышц: мощного внутреннего кольцевого и двух более тонких продольных поверхностных слоев. Наиболее толстая мышечная оболочка в месте соединения яйцепровода с маткой. Слизистая оболочка покрыта псевдомногорядным реснитчатым (мерцательным) эпителием, который в узкой части яйцепровода образует 4-8 мелких продольных складок, а в воронке – до 20-40 более крупных складок. Движение ресничек эпителия направлено в сторону матки.
Яичники (ovaria) – половые железы, продуцируют яйцеклетки и половые гормоны. Расположены на границе тазовой и брюшной полости. У небеременных коров они чаще находятся у края подвздошной ямки на границе с лонным сращением, реже – между подвздошной ямкой и концом маточного рога. К верхушкам рогов матки яичники прикреплены с помощью яичниковой связки, которая является частью широкой маточной связки. Яичниковая связка образует две складки: яйцепроводную, в которую заключен яйцепровод, и яичниковую, более толстую и содержащую пучки гладких мышечных волокон. Складки, соединяясь, образуют открытый яичниковый карман. В нем располагается яичник.
Типичная форма яичников яйцевидная или округлая, но иногда они могут быть совершенно плоскими или иметь неправильную форму, что зависит от наличия в них фолликулов или желтых тел. Длина яичника – наивысшее расстояние между его краями вдоль прикрепленной части – 32-42 мм, ширина – расстояние между свободным и прикрепленным краем – 19-32 мм и толщина – расстояние между боковыми поверхностями – 13-19 мм. Масса каждого яичника 10-19 г. У новорожденных телочек левый яичник немного больше правого, у взрослых животных чаще бывает наоборот. Вся поверхность яичника, за исключением прикрепленного края, покрыта однослойным кубическим эпителием. Под ним располагается собственная белочная оболочка. Она придает яичнику соответствующую форму. Вся остальная масса яичника состоит из двух слоев, между которыми не существует резкой границы: периферического коркового и центрального мозгового. Мозговой слой компактнее коркового. Он содержит кровеносные сосуды, нервы и рыхлую соединительную ткань. Корковый (паренхиматозный) слой занимает большую часть яичника и состоит из клеток соединительной ткани и фолликулов различных стадий развития. Примордиальных (первичных) фолликулов, которые закладываются в период эмбрионального развития, у трехмесячной телочки более 75 тыс., в 1,5-3-летнем возрасте – до 20 тыс., в 9-летнем – около 2,5 тыс., и к 20-летнему возрасту уменьшается до минимума (от 1-2 тыс. до 0). У некоторых видов (приматов, плотоядных, грызунов) примордиальные фолликулы образуют хорошо заметную зону в поверхностной части яичника. У коров это не так выражено.
ПОЛОВЫЕ ОРГАНЫ ОВЦЫ
П реддверие
влагалища относительно короткое – 4-5
см. В стенке его заложены две Бартолиниевы
железы и малые железы преддверия. Длина
влагалища 8-12 см. В каудальной части его
возле отверстия мочеиспускательного
канала возвышается небольшой валик, у
молодых ярочек обнаруживается в этом
месте кольцевая складка. Матка двурогая
двураздельная (рис. 6), массой около 100
г. Длина шейки матки 5-7 см, тела – 3-5 см
и рогов матки – 10-20 см. В основании рога
сращены между собой. Межроговый желоб
хорошо выражен. Слизистая оболочка тела
и рогов матки имеет 76-108 карункулов.
Форма их округлая, на поверхности они
имеют углубления. Слизистая оболочка
шейки матки образует мелкие продольные
и 7-8 крупных поперечных складок, высота
которых в каудальной части цервикального
канала достигает 1 см; вершины складок
направлены в сторону влагалища. Задняя
складка сильно выдается в полость
влагалища, образуя устье шейки матки.
Яйцепроводы длиной 10-15 см. Яичники
расположены в поясничной части брюшной
полости, несколько позади и ниже почек.
Они имеют овальную или яйцевидную форму,
относительно крупные (в период диэструса
1,3x1,1x0,8 см) массой от 0,6 до 3 г. В период
половой охоты яичники заметно
увеличиваются.
реддверие
влагалища относительно короткое – 4-5
см. В стенке его заложены две Бартолиниевы
железы и малые железы преддверия. Длина
влагалища 8-12 см. В каудальной части его
возле отверстия мочеиспускательного
канала возвышается небольшой валик, у
молодых ярочек обнаруживается в этом
месте кольцевая складка. Матка двурогая
двураздельная (рис. 6), массой около 100
г. Длина шейки матки 5-7 см, тела – 3-5 см
и рогов матки – 10-20 см. В основании рога
сращены между собой. Межроговый желоб
хорошо выражен. Слизистая оболочка тела
и рогов матки имеет 76-108 карункулов.
Форма их округлая, на поверхности они
имеют углубления. Слизистая оболочка
шейки матки образует мелкие продольные
и 7-8 крупных поперечных складок, высота
которых в каудальной части цервикального
канала достигает 1 см; вершины складок
направлены в сторону влагалища. Задняя
складка сильно выдается в полость
влагалища, образуя устье шейки матки.
Яйцепроводы длиной 10-15 см. Яичники
расположены в поясничной части брюшной
полости, несколько позади и ниже почек.
Они имеют овальную или яйцевидную форму,
относительно крупные (в период диэструса
1,3x1,1x0,8 см) массой от 0,6 до 3 г. В период
половой охоты яичники заметно
увеличиваются.
Половой аппарат козы по строению и топографии аналогичен половому аппарату овцы.
ПОЛОВЫЕ ОРГАНЫ СВИНЬИ
Половые губы выделяются в виде треугольника, нижний угол которого образован их заостренной спайкой. Клитор тонкий, длинный, оканчивается продолговатой головкой.
Преддверие влагалища длиной 5-8 см, имеет хорошо выраженные продольные складки слизистой оболочки. В толще ее продольными рядами заложены малые вестибулярные железы. В нижней части боковых стенок преддверия имеются пещеристые образования.
Влагалище с преддверием влагалища имеет длину 15-23 см, при соотношении их 2:1. Влагалище узкое, длиной 10-15 см. На границе с преддверием у интактных свинок заметно сужение полового канала, что создает впечатление наличия кольцевой складки, хотя на самом деле она развита слабо. Впереди отверстия мочеиспускательного канала обнаруживается такая же перегородка, как и у телок, но толщина ее варьирует более сильно. Иногда она настолько тонка, что с трудом удается рассмотреть, не повредив ее, а в других случаях хорошо выражена. Иногда обнаруживаются две складочки, на расстоянии 1 см друг от друга.
М атка
двурогая двураздельная. Шейка матки
длиной 10-15 см, без резких границ переходит
сзади во влагалище, впереди – в короткое
(2-3 см) тело матки. Рога матки в правой
половине брюшной полости расходятся
на обе стороны от тела матки и образуют
большое число петель (рис. 7, 6), которые
у молодых свинок располагаются недалеко
от тазовой полости, а у приносивших
приплод – глубоко в брюшной полости.
Длина рогов матки у половозрастных
свинок – 50-75 см, у взрослых свиноматок
90-200 см. Левый рог на 3-8 см длиннее правого.
Задние участки рогов срастаются своими
стенками на протяжении 4-6 см. Диаметр
рогов достигает 6 см, толщина стенок 2-4
мм. Слизистая оболочка матки темно-красного
цвета, сильно складчатая. В области
шейки складки в виде многочисленных
(15-20) треугольных выступов расположены
с боков. Верхушки их не совпадают, в
результате чего канал шейки матки
образует кривую штопорообразную линию.
По направлению к телу матки и особенно
к влагалищу складки сглаживаются.
Величина матки обусловливает вместимость
ее в период супоросности и влияет на
величину помета и массу поросят при
рождении.
атка
двурогая двураздельная. Шейка матки
длиной 10-15 см, без резких границ переходит
сзади во влагалище, впереди – в короткое
(2-3 см) тело матки. Рога матки в правой
половине брюшной полости расходятся
на обе стороны от тела матки и образуют
большое число петель (рис. 7, 6), которые
у молодых свинок располагаются недалеко
от тазовой полости, а у приносивших
приплод – глубоко в брюшной полости.
Длина рогов матки у половозрастных
свинок – 50-75 см, у взрослых свиноматок
90-200 см. Левый рог на 3-8 см длиннее правого.
Задние участки рогов срастаются своими
стенками на протяжении 4-6 см. Диаметр
рогов достигает 6 см, толщина стенок 2-4
мм. Слизистая оболочка матки темно-красного
цвета, сильно складчатая. В области
шейки складки в виде многочисленных
(15-20) треугольных выступов расположены
с боков. Верхушки их не совпадают, в
результате чего канал шейки матки
образует кривую штопорообразную линию.
По направлению к телу матки и особенно
к влагалищу складки сглаживаются.
Величина матки обусловливает вместимость
ее в период супоросности и влияет на
величину помета и массу поросят при
рождении.
Яйцепроводы у свиньи выделяются отчетливо, длина их 15-28 см. Истмическая часть узкая, диаметром 2-3 мм, составляет четвертую часть длины яйцепровода, средняя часть – ампула, толщиной 3-5 мм и воронка – 5-6 мм. Конечная часть воронки очень расширена.
Яичники у свинок вскоре после рождения – овальной формы, гладкие, массой около 0,02 г; в них насчитывается до 60 тыс. первичных ооцитов. К 5 мес. яичники приобретают форму и внешний вид боба, масса их увеличивается до 3 г. У половозрелых животных они похожи на ягоду малины, имеют неправильную форму и размеры 4x3x3 см. У взрослых свиноматок яичники округлой формы, диаметром около 5 см, поверхность их бугристая, что связано с наличием большого количества крупных фолликулов и желтых тел. Расположены они в брюшной полости на уровне 4-5-го поясничного позвонка; находятся в хорошо выраженных яичниковых карманах и тесно прилегают к мощно развитой бахромке яйцепровода.
ПОЛОВЫЕ ОРГАНЫ КОБЫЛЫ
Половые губы, ограничивающие половую щель, вверху образуют заостренную спайку, переходящую в шов промежности длиной 10-12 см; внизу губы замыкаются в закругленный угол половой щели, прикрывающий полушаровидную головку клитора. По бокам головки имеются хорошо выраженные складки слизистой оболочки преддверия.
Преддверие влагалища длиной 8-16 см. По бокам его под слизистой оболочкой и под суживателем половой щели имеются два пещеристых тела, окруженные фиброзной оболочкой. В верхней части преддверия на 1,5-2,5 см впереди половой щели расположены выводные протоки Бартолиниевых желез, по бокам его в толще слизистой оболочки расположены два ряда трубчатых желез, открывающихся в просвет преддверия несколькими выводными протоками.
Влагалище длиной 15-30 см. У молодых кобылок в задней части по всей окружности влагалища имеется хорошо выраженная поперечная складка (мочеполовой клапан) толщиной 2-3 мм с центральным отверстием.
Матка кобылы типично двурогая. Шейка матки длиной 4-8 см, диаметром 3-5 см, сзади выдается в полость влагалища. Эта часть ее длиной 2-2,5 см имеет форму втулки; в центре ее расположено устье шейки матки. Тело матки представляет собой большой полый орган длиной 8-15 см и шириной 7-12 см. Впереди разделяется на два рога; длина их 8-15 см, ширина 4-7 см. Рога расходятся в стороны, несколько вперед и вверх, поэтому большая кривизна их расположена снизу. В области малой кривизны серозная оболочка переходит в широкие маточные связки. Слизистая оболочка тела и рогов матки образует много продольных складок, сверху покрыта высоким цилиндрическим мерцательным эпителием; в толще ее расположены ветвящиеся трубчатые железы. В области шейки матки слизистая оболочка формирует большое количество продольных складок различной высоты.
Яйцепроводы имеют вид сильно извитых трубочек, длиной 15-30 см. Маточные отверстия их находятся вблизи верхушки рогов матки на вершине сосочков, образованных стенками яйцепроводов. Расширенный брюшной конец яйцепровода (воронка) имеет неровные края, тесно прилегает к яичнику в области овуля-ционной ямки. Широкий край ее - бахромка полностью прикрывает овуляционную ямку.
Яичники чаще бобовидной (рис. 7, а), иногда округлой или неправильной овальной формы, длиной 6 см, толщиной 4 см и шириной 3 см. Подвижные, т. е. не имеют строго определенного месторасположения, находятся в поясничной части брюшной полости вблизи верхушек рогов матки, несколько выше их. Правый яичник расположен на уровне третьего поясничного позвонка, на ладонь ниже от почек и в сторону тазовой полости, левый — на уровне четвертого поясничного позвонка на 4-6 см в сторону от медиальной линии. Снаружи яичники покрыты серозной оболочкой. Под ней расположена плотная фиброзного типа белочная оболочка, покрывающая почти весь яичник, за исключением овуляционной ямки, свободной от серозной оболочки и выстланной зародышевым эпителием. Расположена овуляционная ямка по малой кривизне над корковым слоем и имеет бархатистый вид. Мозговой слой яичника расположен по большой кривизне. В этом месте прикрепляется яичниковая связка и здесь начинается серозный покров яичника.
ПОЛОВЫЕ ОРГАНЫ СУКИ
Половые губы валиковидной формы, образуют верхний тупой и нижний заостренный углы половой щели. Заостренный угол опускается ниже седалищных бугров. Промежность большая. Клитор хорошо развит, часть его скрыта в складке слизистой оболочки. По бокам преддверия влагалища имеются пещервстые образования (луковицы преддверия – bylbus vestibuli). Во время полового акта они наполняются кровью, что приводит к сильному сужению просвета влагалища. Выводные протоки желез находятся на нижней стенке преддверия влагалища.
Влагалище длинное, слизистая оболочка его имеет многочисленные продольные и поперечные складки.
Матка двурогая двураздельная. Тело матки короткое, в передней части на протяжении 2-4 см разделено перегородкой. Рога относительно длинные и имеют прямолинейное направление, основаниями сходятся друг с другом под углом (рис. 8). Шейка матки короткая и плотная. Выдается в просвет влагалища; нижняя складка выступающей части имеет запирательный валик. Широкие маточные связки и яйцепроводная складка покрыты обильными жировыми отложениями. От верхушки рогов матки по направлению к внутреннему паховому кольцу отходят круглые маточные связки. При наличии рудимента пахового канала возможно образование паховых маточных грыж.
Я ичники
овальной формы, нередко уплощены,
скрыты в яичниковом кармане; диаметр
их зависит от породы и величины самки
и в среднем составляет 1-2 см. В половой
сезон поверхность яичников становится
бугристой вследствие появления крупных
фолликулов и желтых тел. Яйцепроводы
извилистые, расширенная часть их
заканчивается хорошо развитой бахромкой.
ичники
овальной формы, нередко уплощены,
скрыты в яичниковом кармане; диаметр
их зависит от породы и величины самки
и в среднем составляет 1-2 см. В половой
сезон поверхность яичников становится
бугристой вследствие появления крупных
фолликулов и желтых тел. Яйцепроводы
извилистые, расширенная часть их
заканчивается хорошо развитой бахромкой.
ПОЛОВАЯ ФУНКЦИЯ САМОК
Животные способны к воспроизведению в течение репродуктивного периода жизни. Этому периоду предшествует период полового созревания, во время которого половые органы растут и достигают состояния зрелости. У разных видов происходит это в различном возрасте. В последующем при сохранении здоровья и наличии хороших условий кормления и содержания половая функция у них проявляется практически всю жизнь.
ПЕРИОД ПОЛОВОГО СОЗРЕВАНИЯ
Способность к воспроизведению у самок возникает постепенно, по мере их общего роста и развития. Вначале происходит равномерный быстрый рост и созревание гипофиза. Вследствие его гормонального влияния отмечается увеличение размеров и массы яичников и диаметра фолликулов в них. Созревающие фолликулы вырабатывают половые гормоны (эстрогены), которые обеспечивают рост и развитие матки, а также других участков половых путей. Эти изменения приводят половой аппарат самки в состояние функциональной готовности. Внешним проявлением этой готовности является первая течка и половая охота. В яичниках происходит овуляция. Наступает половая зрелость.
Хотя наступление половой зрелости у самок можно установить с точностью до одного дня, но процессы, ведущие к этому, происходят постепенно. Половое созревание не заканчивается первой охотой и овуляцией, оно представляет собой прогрессирующий процесс. Устанавливается характерная для данного вида ритмичность проявления половой функции, более полноценными становятся количественные и качественные изменения в половых органах. Наблюдения за свинками показывают, что с увеличением возраста их при покрытии с 5,5 до 8 мес. прогрессивно увеличивается число овулирующих фолликулов в яичниках, снижается гибель яйцевых клеток, возрастает количество нормальных эмбрионов к 25-му дню супоросности и живых поросят к моменту рождения. После 245 дней фактор времени уже не имеет значения.
У самок сельскохозяйственных животных половая зрелость наступает в следующие сроки: у кобылок – в возрасте 15-24 мес. (в течение их второго лета), живая масса варьирует в зависимости от породы, у ярочек - в 5-8 мес., живая масса 27-34 кг, у коз – в 5-7 мес., живая масса 10-30 кг, у телок молочных пород – в 8-13 мес. (возможные колебания от 6 до 18 мес.), живая масса 160-270 кг; у мясных пород (европейские) – в 10-15 мес., у свинок – в 5-7 мес. при достижении живой массы более 68-90 кг, у сук – в 6-8 мес.
На половое созревание большое влияние оказывают уровень кормления, условия содержания, порода, сезон рождения и другие факторы. Повышение уровня кормления у крупного рогатого скота и овец сокращает возраст наступления половой зрелости при одновременном увеличении к этому моменту живой массы. Так, у телок голштинской породы при уровне кормления 146% первую половую охоту наблюдали в возрасте 9,2 мес., при стандартном (100%) – в 11,2 мес. и при низком уровне (62%) – в 20,2 мес. Живая масса их к этому времени была – соответственно 277, 265 и 303 кг. Оплодотворяемость после первого осеменения (естественного) составила 58,68 и 79%, а число осеменений на стельность – 1,48, 1,41 и 1,55. Отелились они в возрасте 27,9, 28,5 и 32,0 мес. при достижении живой массы 548, 483 и 384 кг. Живая масса телят при рождении была выше (41, 39 и 36 кг), а мертворождаемость ниже (6, 12 и 13%) в группах с высоким и стандартным уровнем кормления. Однако показатели плодовитости при последующих двух отелах предпочтительнее были в средней группе, а выбраковано по причине бесплодия меньше в группе животных, выращенных на низком уровне кормления – 6% (при высоком и стандартном уровне – соответственно 20 и 12%). С учетом этих показателей рекомендуется средний (стандартный) уровень кормления телок [Maynard and Loosli. Animal Nutrition, 1969].
Величина живой массы у телок в 6-месячном возрасте играет важную роль в наступлении половой зрелости. Однако обильное кормление и, как следствие, ожирение в раннем возрасте у молочного скота отрицательно влияет на развитие секреторной ткани вымени. Для животных крупных пород среднесуточный прирост живой массы в первые 15 мес. не должен превышать 750 г. У свиней половое созревание стимулирует кратковременное повышение уровня кормления (особенно увеличение содержания белка), оптимальный режим содержания и присутствие производителя. Групповое содержание, смешивание гнезд и смена помещения действуют также благоприятно. Продолжительный перекорм и безвыгульное содержание свинок в период выращивания, напротив, задерживают наступление охоты и приводят к ослаблению ее признаков. Задерживается наступление половой зрелости и в холодное время года.
Телки молочных пород проявляют первую охоту раньше, чем животные мясных пород. Зарегистрированы отелы у животных джерсейской породы в возрасте 13 мес., гернсейской – в 16 мес., айрширской и голштинской – в 17 мес. Следовательно, первая охота может наступить даже в 4-месячном возрасте.
Сезон рождения оказывает заметное влияние на сроки наступления половой зрелости у кобыл, сук и особенно у овец. Так, если ярочка родится зимой или в начале весны и будет нормально развиваться, охота у нее проявится в этот же год осенью, хотя ей будет только 5-8 мес. от роду. Но если рождение произойдет поздней весной, то она не достигнет такого состояния, когда у нее может проявиться охота в этот половой сезон (осенью текущего года). Первая охота в таком случае наступит лишь в начале осени следующего года, т. е. на втором году жизни.
РЕПРОДУКТИВНЫЙ ПЕРИОД
С достижением половой зрелости у самок, проявляется инстинкт спаривания, а их половые железы вырабатывают способные к оплодотворению половые клетки. При этом их половой аппарат подчиняется определенному функциональному ритму. Процессы, которые лежат в основе этого ритма и обусловливают его, происходят в яичниках. Они заключаются в периодическом созревании одного или нескольких фолликулов, овуляции и образовании желтого тела (тел). Внешним проявлением этих изменений является течка (выделение слизи из половых органов) и половая охота (готовность самки к спариванию с самцом).
Определенная периодичность изменений в половых органах самки дала основание говорить о циклической природе половой функции. Впервые в литературу термин "половой цикл" ввел Маршалл. Под половым циклом следует понимать все многообразие структурных и функциональных изменений, происходящих в половом аппарате, эндокринной и других системах организма самки от одной половой охоты до другой (или от одной овуляции до другой). Период времени между двумя половыми охотами (или овуляциями) составляет продолжительность полового цикла.
Число половых циклов и их продолжительность не одинаковы у различных животных. Многие дикие животные на протяжении года имеют только один половой цикл. Это моноциклические животные. Время года, когда у них проявляется половая активность, называют половым сезоном. Собак и кошек также относят к моноциклическим животным, потому что у них промежутки между половыми циклами длительны. В течение года у сук течка проявляется обычно два раза: поздно зимой и в конце лета. Но нередко у них наблюдается четыре цикла в году.
Сезонность в размножении диких млекопитающих в большинстве случаев можно объяснить естественными причинами и прежде всего теми требованиями, которые предъявляет потомство к окружающей среде. Наличие тепла и света, водопоя, легкодоступного и усвояемого корма благоприятствует появлению потомства и облегчает ему дальнейшую жизнь. Тем самым эти факторы определяют время наступления полового сезона.
У сельскохозяйственных животных вследствие улучшения условий кормления и содержания и ослабления сезонных различий в их содержании, а также ослабление неблагоприятных климатических воздействий на них половой сезон удлинился, а некоторые виды (коровы, свиньи) могут проявлять половую цикличность на протяжении всего года. Таких животных относят к полициклическим. У овец и коз отмечается несколько половых циклов, следующих один за другим, после чего половая функция ослабевает. Это – полициклические животные с выраженным половым сезоном.
Учение о половом цикле впервые сформулировал Хипп. Он выделил в половом цикле следующие стадии (фазы).
Проэструс – подготовительный период, непосредственно предшествующий эструсу. Характеризуется началом быстрого роста отдельных фолликулов в яичниках. Растущие фолликулы вырабатывают эстрогены. Под их влиянием усиливается кровоснабжение половых органов, вульва набухает, слизистая влагалища приобретает красный цвет вследствие увеличения притока крови. Клетки ее подвергаются изменениям, которые у большинства видов приводят к ороговению. Секреция слизи клетками влагалища и шейки матки возрастает.
Матка увеличивается, слизистая оболочка ее становится кровенаполненной и отечной, а маточные железы – активными. У сук в этой стадии выделения из половых органов имеют кровянистый оттенок.
Эструс – кульминационный момент полового цикла. В этой фазе в центральной нервной системе животного появляется очаг повышенной возбудимости, который приводит к развитию половых ощущений. Половое возбуждение принимает господствующее положение. Животное стремится к спариванию с самцом, допускает на себя садку и стоит спокойно под ним (половая охота). Половые губы опухают еще более заметно, покраснение слизистых оболочек половых путей усиливается. Маточные железы и клетки шейки матки и влагалища секретируют много слизи жидкой консистенции. Канал шейки матки расслабляется, и у многих животных отмечается вытекание слизи (течка). Матка увеличена, хорошо проявляет ригидность. Быстро созревает фолликул (фолликулы) в яичниках, и в конечном итоге происходит разрыв его и выход яйцеклетки (овуляция).
Метэструс – стадия, следующая за эструсом (послетечковая). Эпителиальные клетки вскрывшегося фолликула (гранулеза) подвергаются гипертрофии и превращаются в лютеиновые; формируется желтое тело. В слизистой оболочке матки разрастаются кровеносные сосуды, маточные железы становятся очень активными. Выделение слизи из влагалища ослабевает, канал шейки матки закрывается. Уменьшается приток крови к наружным половым органам, вульва сморщивается. Прекращается половая охота.
Диэструс – период между следующими один за другим циклами. Характеризуется доминированием циклического желтого тела. Маточные железы активны, мускулатура матки расслаблена, шейка матки закрыта. Количество цервикальной слизи небольшое, консистенция ее вязкая. Слизистая оболочка влагалища бледная.
Анэструс – длительный период полового покоя, в течение которого функция яичников ослаблена. Развитие фолликулов не происходит или слабо выражено. Матка уменьшена и анемична, шейка плотно закрыта. Вагинальная слизь скудная, вязкой консистенции. Слизистая влагалища бледная.
А.П.Студенцов создал учение о половом цикле. В половом цикле он выделяет три стадии.
Стадия возбуждения – характеризуется ярким проявлением основных феноменов (явлений) полового цикла: течки, полового возбуждения, охоты, созревания фолликулов и овуляции. Каждый из этих четырех феноменов является специфическим и отражает одну какую-либо сторону полового цикла. Протекают они во взаимной связи, но возникают и проявляются неодновременно. В стадии возбуждения в половой и других системах организма преобладают пролиферативные процессы.
Течка – процесс выделения слизи из половых органов как следствие морфологических изменений полового аппарата самки. Характеризуется ярко выраженными пролиферативными процессами: гиперемией всех компонентов полового аппарата, новообразованием и разрастанием секретирующих клеток и желез слизистых оболочек яйцепроводов, рогов, тела и шейки матки. Число слоев эпителия во влагалище и преддверии влагалища увеличивается с 3-4 до 18-20. Одновременно отторгаются поверхностные эпителиальные клетки.
Диагностируют течку осмотром наружных половых органов, влагалища, шейки матки, исследованием выделяющейся из половых органов слизи, клиническими и лабораторными методами.
Половое возбуждение (общая реакция) – изменение в поведении самки, возникающее в связи с созреванием фолликулов во время стадии возбуждения. Проявляется половое возбуждение позднее течки и характеризуется ярко выраженной общей реакцией организма в виде беспокойства, отказа от корма, другими признаками. Самка проявляет "интерес" к самцу, может прыгать на него или других самок, но садку самца на себя не допускает.
Охота (половая охота) – положительная сексуальная реакция самки на самца. Во время охоты самка стремится к самцу, принимает позу для полового акта, часто производит акт мочеиспускания, допускает садку и коитус.
Стадия торможения – стадия ослабления и полного прекращения феноменов полового цикла – течки и полового возбуждения; вместо охоты ярко выраженный отбой (самка агрессивно настроена к самцу: стремится его укусить, ударить или убежать от него); на месте овулировавшего фолликула формируется желтое тело. Главная особенность этой стадии - преобладание инволюционных процессов.
Стадия уравновешивания - стадия отсутствия признаков течки, охоты и полового возбуждения. Характеризуется уравновешенным общим состоянием животного, наличием в яичниках и желтых тел, и фолликулов.
В настоящее время в связи с глубоким изучением гормональных изменений в организме и внедрением способов искусственного контроля воспроизводительной функции используют, как правило, терминологию стадий полового цикла по Хиппу.
СОЗРЕВАНИЕ ФОЛЛИКУЛОВ И ОВУЛЯЦИЯ
В период эмбрионального развития в яичниках женских особей закладывается большое количество примордиальных фолликулов. Каждый фолликул состоит из оогония и слоя фолликулярных клеток. Располагаются фолликулы в корковом слое. В яичниках 6-месячного эмбриона человека насчитывается до 6 млн. первичных фолликулов. К моменту рождения число их уменьшается до 200-300 тыс., но к 3 годам из них остается 70%, к 6 годам – 20%. Оставшиеся фолликулы сохраняются до зрелого возраста. После рождения новые ооциты и фолликулы не образуются. У крупного рогатого скота образование первичных фолликулов возможно и в постнатальный период (Г.А.Черемисинов).
В процессе роста фолликул глубже погружается в корковый слой яичника. Рост вначале заключается в увеличении всего фолликула и самой половой клетки. После увеличения в несколько раз оогоний превращается в первичный ооцит. Вокруг него образуется прозрачная оболочка. В это же время происходит рост и увеличение количества плоских эпителиальных (фолликулярных) клеток вокруг ооцита. Клетки эти приобретают кубическую, а затем цилиндрическую форму; количество слоев увеличивается до двух, затем -до трех и более, после чего клетки опять приобретают кубическую форму.
После окончания роста яйца фолликулярные клетки в нескольких местах расходятся и в образовавшихся полостях накапливается жидкость. Постепенно полости объединяются в одну большую полость. Яйцо оттесняется к стенке фолликула. Оно располагается в скоплении эпителиальных клеток, в яйценосном (яйцевом) бугорке (рис. 9). Те клетки, которые непосредственно окружают яйцо, образуют кольцо – лучистый венец. Ободок клеток, выстилающих полость фолликула, называют гранулезой. Содержащаяся в полости фолликулярная жидкость вязкая, богата эстрогенными веществами. В ней формируются структуры, состоящие из кристаллов микроскопической величины, каналов и сферических жировых капель диаметром около 500 мкм. В эструсе эти структуры становятся большими и на типичных кристаллах появляется нитевидный материал,
содержащий ДНК. В стадии диэструс происходит дезорганизация микрокристаллов. Фолликулярная жидкость способствует увеличению и растяжению
ф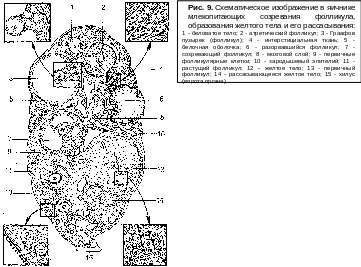 олликула
и выходу его из коркового слоя на
поверхность яичника.
олликула
и выходу его из коркового слоя на
поверхность яичника.
Гранулезные клетки окружены мембраной. Вокруг нее расположены два слоя клеток, которые образуют оболочку фолликула. Внутренний слой (theca interna) окружает фолликул с момента роста яйца до овуляции. Клетки внутреннего слоя играют большую роль в продуцировании половых гормонов (эстрогенов) и в образовании желтого тела. Их ядра округлые, в отличие от удлиненных, расположенных концентрическими слоями вокруг фолликула, ядер клеток наружной оболочки (theca externa). Состоит наружная оболочка из соединительной ткани и гладких мышечных волокон, обильно снабжена кровеносными сосудами.
Зрелый фолликул питает кровью артериола. У внутреннего полюса фолликула этот сосуд делится на 3-4 ветви, образующие в наружной оболочке широкопетлистую сеть из артериол, а во внутренней – узкопетлистую сеть из капилляров.
По мере роста фолликул становится округлым и набухшим и начинает возвышаться над поверхностью яичника. Внутренняя оболочка фолликула приближается, а затем соприкасается с оболочкой яичника. Обе они истончаются, появляется конусовидный выступ, на верхушке которого образуется разрыв. Яйценосный бугорок теряет связь со стенкой фолликула, и содержимое его, в том числе и яйцеклетка, медленно вытекает через разрыв. В некоторых случаях разрыв сопровождается выбросом части жидкости, а остальная часть ее затем продолжает медленно вытекать. Так происходит овуляция.
Перед овуляцией отмечается первое редукционное деление первичного ооцита и образование первого полярного тельца. Контролируется это, как и овуляция, гонадотропинами, которые стимулируют секрецию фолликулярными клетками особого вещества, специально предназначенного для индукции созревания.
У многих видов овуляция происходит спонтанно, т.е. независимо от наличия или отсутствия полового акта. Главную роль в механизме овуляции играют ЛГ (лютеинизирующий гормон) и окситоцин. Выделение ЛГ и увеличение синтеза простагландина Е приводит к расширению сосудов фолликула, гиперемии, повышению проницаемости капилляров и базальной мембраны. За несколько часов до овуляции преобладает действие простагландина Ф2α; сосуды суживаются, многие капилляры лопаются, соединительнотканные волокна стенки фолликула подвергаются дистрофии. Гранулеза истончается и на выступающей из яичника стенке фолликула перед овуляцией появляется конусовидный выступ.
Раздражение рецепторов полового тракта стимулирует выделение гонадотропных гормонов. На это указывает то, что у некоторых животных (крольчиха, кошка, хомяк) овуляция происходит только после полового акта. У животных со спонтанной овуляцией (у коровы) коитус ускоряет овуляцию. Удаление гипофиза делает овуляцию невозможной.
В процессе разрыва стенки фолликула определенную роль играет повышение давления жидкости в нем, сокращение гладких мышечных волокон яичника и стенки фолликула, а также разрушающее действие фермента гиалуронидазы.
На месте лопнувшего фолликула образуется желтое тело. Это происходит следующим образом. После разрыва стенка фолликула спадается, становится складчатой. В центре каждой складки, состоящей из гранулезы и внутренней оболочки, образуется петля из артериолы. От нее к внутренней оболочке отходит густая капиллярная сеть, через которую происходит васкуляризация желтого тела. Клетки желтого тела (лютеиновые) развиваются в основном из клеток гранулезы. Превращение это сопровождается значительным увеличением их в размере. Они располагаются рядами и имеют многоугольную форму. Между ними размещаются соединительнотканные клетки и кровеносные сосуды. Они образуют перегородки, направляющиеся к центру желтого тела. Благодаря перегородкам желтое тело имеет дольчатое строение. Клетки его продуцируют гормон прогестерон, имеют желтую окраску, что придает всему образованию желтый цвет.
Если в период охоты животное не было осеменено или же после осеменения не оплодотворилось, то желтое тело подвергается обратному развитию (инволюции). В этот период его называют желтым телом полового цикла. При оплодотворении и наступлении беременности у животного желтое тело называют желтым телом беременности.
Развитие фолликулов в яичниках самок происходит постоянно. Однако до наступления половой зрелости ни один из фолликулов не достигает состояния полной зрелости и подвергается обратному развитию без выхода яйцеклетки. Этот процесс называют атрезией фолликулов. Начинается атрезия с деструктивных изменений в ооците или самом фолликуле. В малых фолликулах сначала разрушается ооцит, затем дегенерирует и исчезает весь фолликул. В крупных фолликулах атрезия нередко сопровождается превращением клеток гранулезы в лютеальные и образованием структуры, сходной с желтым телом. У половозрелых животных многие фолликулы на различных стадиях развития также подвергаются атрезии.
НЕПОЛНОЦЕННЫЕ ПОЛОВЫЕ ЦИКЛЫ
Неполноценным называют такой половой цикл, в течение которого выпадает один или несколько феноменов. В связи с этим различают неполноценные циклы: анэстральный (отсутствие течки), алибидный (отсутствие охоты, "тихая овуляция"), ановуляторный (отсутствие овуляции), ареактивный (отсутствие признаков общей реакции). Могут быть смешанные неполноценные половые циклы (ареактивно-ановуляторные и др.).
Особый интерес представляют циклы ановуляторные, так как по внешним проявлениям они в большинстве случаев не отличаются от нормального, сопровождающегося овуляцией. У крупного рогатого скота и лошадей ановуляторные циклы являются одной из важных причин бесплодия. При этих циклах крупные фолликулы подвергаются атрезии, лютеинизации или дегенерации (кистозному перерождению). При атрезии вначале погибает яйцевая клетка, затем разрушается фолликулярный эпителий, сам фолликул уменьшается в размере, приобретает более упругую консистенцию и в течение 3-4 дней полностью исчезает. В процессе лютеинизации происходит постепенное заполнение полости неовулировавшего фолликула клеточными элементами его стенки. Обычно размножаются и разрастаются клетки гранулезы, которые затем превращаются в лютеальные клетки. Лютеинизация начинается от периферии фолликула и распространяется к центру. В разрастающихся клетках откладывается пигмент лютеин. К 5-6-му дню возникает атретическое желтое тело. Дегенерация – это гибель яйцеклетки и последующее увеличение фолликула в диаметре вследствие накопления жидкости и в конечном итоге превращение в кисту. Киста может образоваться уже на 5-7-й день после охоты. При ановуляторных циклах оплодотворение невозможно.
Алибидный половой цикл характеризуется отсутствием охоты у животного при наличии течки и созревания фолликулов и овуляции в яичниках. Особенно часто наблюдается у коров – в 10-30% случаев, а в первые 2 мес. после отела может достигать 65%. У телок первая овуляция, как правило, не сопровождается охотой. Оплодотворение в алибидный цикл возможно.
НЕЙРОГУМОРАЛЬНАЯ РЕГУЛЯЦИЯ ПОЛОВОЙ ФУНКЦИИ САМОК
Проблема обеспечения нормального проявления воспроизводительной функции у самок животных имеет исключительно важное значение. Решение ее в условиях современных технологий животноводства связано не только с созданием оптимальных условий кормления и содержания животных, но и с использованием биотехнологических приемов и способов управления процессами размножения. Эффективность этих способов зависит от того, насколько строго учитывается механизм естественного контроля этих процессов.
ФАКТОРЫ, ОБУСЛОВЛИВАЮЩИЕ ПОЛОВУЮ ЦИКЛИЧНОСТЬ
Регуляция половой функции осуществляется центральной нервной системой (гипоталамусом) во взаимодействии с гормонами гипофиза, половых и других эндокринных желез и рядом биологически Активных веществ.
В нейросекреторных клетках ядер гипоталамуса секретируется гонадотропин-рилизинг-гормон (ГнРГ, гонадолиберин). Это вещество (декапептид) из гипоталамуса перемещается в ножку гипофиза (medium eminence) и через соединения вилкообразных нервных окончаний с капиллярами (вазоневральные синапсы) попадает в кровь и благодаря особой системе кровообращения переносится в переднюю долю гипофиза (рис. 10). В гипофизе гормон связывается с мембраной секретирующей ткани и изменяет ее полярность, вследствие чего возрастает проницаемость для ионов кальция. По микроканалам кальций достигает секреторных гранул и вызывает освобождение их содержимого (гонадотропных гормонов).
Г нРГ
не имеет половой и видовой специфичности,
но обладает строгой специфичностью
биологического действия, т. е. стимулирует
выделение только ЛГ и ФСГ. Он синтезирован.
нРГ
не имеет половой и видовой специфичности,
но обладает строгой специфичностью
биологического действия, т. е. стимулирует
выделение только ЛГ и ФСГ. Он синтезирован.
В регулировании выделения гонадотропинов принимают участие и другие специфические neurotransmitter субстанции. Норадреналин стимулирует выделение ФСГ и ЛГ; задержка превращения дофамина (dofamine) в норадреналин блокирует эстрадиолиндуцированное выделение ЛГ, которое необходимо для вызова овуляции. Серотонин угнетает базальную секрецию ЛГ и регулирует другие нейросекреторные системы. Дофамин играет также важную роль в контроле выделения пролактина.
Определенная роль в осуществлении контроля ЦНС за деятельностью гипофиза принадлежит и ряду эндогенных опиоидных пептидов, выделяемых мозговой тканью (р-эндорфин, метэнкефалин). Эти вещества обнаружены в высоких концентрациях в гипоталамусе и крови гипофизарной портальной системы в течение полового цикла и в другие фазы репродуктивного периода. Инъекция опиоидных пептидов угнетает секрецию ФСГ и ЛГ, но стимулирует выделение пролактина. Введение же налоксона (naloxone), специфического опиоидного ингибитора, вызывает повышение концентрации и частоты эпизодического выделения го-надотропных гормонов. Очевидно, что опиоидные пептиды влияют на выделение гонадотропинов путем предотвращения выделения ГнРГ из нервных окончаний в ножку гипофиза. Поэтому ГнРГ не попадает в кровь портальной системы и в переднюю долю гипофиза для стимулирования выделения ФСГ и ЛГ.
У домашних животных секреция ФСГ и ЛГ
контролируется двумя функционально
различными, но тесно взаимодействующими
гипоталамическими системами. Соответственно
имеется два гипоталамических центра.
Тоническая эпизодическая система
обеспечивает продолжительную базальную
секрецию гонадотропинов, тем самым
стимулирует рост зародышевой и эндокринной
ткани яичника. Пулъсаторная система
контролирует непродолжительные массивные
выделения гонадотропинов, особенно ЛГ,
необходимые для овуляции.
домашних животных секреция ФСГ и ЛГ
контролируется двумя функционально
различными, но тесно взаимодействующими
гипоталамическими системами. Соответственно
имеется два гипоталамических центра.
Тоническая эпизодическая система
обеспечивает продолжительную базальную
секрецию гонадотропинов, тем самым
стимулирует рост зародышевой и эндокринной
ткани яичника. Пулъсаторная система
контролирует непродолжительные массивные
выделения гонадотропинов, особенно ЛГ,
необходимые для овуляции.
В гипоталамусе синтезируется и один из гормонов гипофиза - окситоцин. Этот гормон продвигается вдоль аксонов гипоталамо-гипофизарного тракта в заднюю долю гипофиза (рис. 11) и скапливается в ее клетках — питуицитах, где гранулы окситоцина связываются со специфическим белком. Вытяжку из задней доли гипофиза, богатую окситоцином, называют питуитрином.
Окситоцин вызывает сокращение гладких мышц матки и тем са-
мым ускоряет роды. Быстро разрушается в крови ферментом окситоциназой, но в период родов содержится в повышенных количествах. Расширение влагалища в это время рефлекторно вызывает выделение гормонов. При коитусе окситоцин также выделяется в кровь под влиянием нервных импульсов и усиливает сокращение мускулатуры матки, что способствует продвижению спермиев в половых путях самки.
Чувствительность матки к окситоцину определяется концентрацией эндометриальных рецепторов к этому гормону. Количество же рецепторов зависит от соотношения прогестерона и эстрогенов. После формирования желтого тела и повышения содержания прогестерона количество рецепторов уменьшается, а в период лютеальной регрессии, напротив, возрастает (у овец 500-кратно, Flint et. al., 1992). Введение экзогенного эстрадиола индуцирует преждевременное увеличение рецепторов.
Окситоцнн стимулирует и сокращение миоэпителиальных клеток вымени, тем самым способствуя выведению молока из альвеолярного отдела в молочную цистерну. Кроме того, этот гормон играет роль в регрессии желтого тела у жвачных. Он продуцируется в обнаруживаемых количествах гранулезой фолликулов и желтым телом. Простагландин Ф2α стимулирует секрецию окситоцина желтым телом.
Передняя доля гипофиза сама секретирует гормоны. Базофильные клетки вырабатывают ФСГ и ЛГ, ацидофильные – пролактин. Это гонадотропные гормоны. Они являются гликопротеидами. ФСГ и ЛГ состоят из двух субъединиц, одна из которых общая для обоих гонадотропинов и тиреостимулирующего гормона. Часть, представленная углеводородной цепью, специфична и обусловливает биологическую активность гонадотропина; вторая, представленная полипептидной цепью, ответственна за иммунологические свойства. Пролактин представляет единую полипептидную цепь, в которой 211 аминокислотных остатков.
У самок ФСГ вызывает пролиферацию клеток гранулезы фолликулов в яичниках и способствует развитию их от полостной до преовуляторной стадии. Он активизирует также биохимические процессы в клетках гранулезы и оболочки фолликулов, обеспечивает синтез эстрогенов.
ЛГ стимулирует синтез половых гормонов всеми клетками яичника: желтого тела, гранулезы и оболочки фолликулов и интерстициальными. Определенная базальная концентрация его является обязательным условием для осуществления синтеза гормонов в яичниках. ЛГ вызывает также увеличение кровотока в яичниках и овуляцию фолликулов, подготовленных фолликул остимулирующим гормоном.
Следует отметить, что ФСГ и ЛГ проявляют свое действие в комплексе. Так, ФСГ оказывает слабое действие на рост фолликулов в яичниках, а ЛГ не влияет на их рост, но вместе при строго определенном соотношении эти гормоны обеспечивают полное созревание фолликулов и их овуляцию, а также секрецию эстрогенов. Под влиянием ЛГ-образуется желтое тело и начинается секреция прогестерона. Соотношения этих гормонов в гипофизе определяют особенности течки и охоты у разных видов животных. У кобыл высокое содержание ФСГ и низкое содержание ЛГ, поэтому половая охота у них наиболее продолжительная, а овуляция происходит задолго до окончания охоты. Нередко овуляция задерживается или совсем не происходит. У коров, наоборот, содержание ЛГ высокое, ФСГ – низкое. У них охота менее продолжительна, а нередко овуляция происходит без признаков охоты ("тихая овуляция").
Пролактин (ЛТГ) вместе с АКТГ и соматотропином образует гипофизарный лактогенный комплекс. Как и эстрогены, пролактин стимулирует рост протоков вымени и оказывает синергичное прогестерону действие на дольчато-альвеолярную систему вымени. Вместе с кортикостероидами пролактин вызывает и поддерживает секрецию молока в развитом вымени. ЛГ и пролактин образуют лютеотропный комплекс, необходимый для поддержания структуры и функции желтого тела у лабораторных животных. У коров, овец и свиней лютео-тропное действие оказывает лютеинизирующий гормон.
Гонадотропины различных животных отличаются друг от друга по химическому составу и антигенным свойствам. Но по биологическому действию они не обладают половой или видовой специфичностью.
В плаценте человека и некоторых обезьян образуется вещество, обладающее биологическим действием, сходным с действием ЛГ; выделяется с мочой. Это хорионический (плацентарный) гонадотропин (ХГТ). У женщин его удается обнаружить в моче уже на 9-й день после зачатия. Наибольшее количество его выделяется между 50 и 70-м днями беременности, после чего экскреция резко снижается.
ХГТ стимулирует образование половых гормонов в яичниках и семенниках, а также превращение андро-генов в эстрогены в плаценте, вызывает овуляцию зрелых фолликулов, а у неполовозрелых животных - рост фолликулов и образование желтых тел. Применяется животным для ускорения овуляции при ее задержке или при наличии фолликулярных кист, для синхронизации овуляции после предварительной обработки препаратами прогестерона, при вызове суперовуляции и др. Одна единица ХГТ (1 Е) равна 0,1 мг стандартного вещества.
У беременных кобыл в возвышениях слизистой оболочки матки в месте перехода тела в беременный рог образуется вещество, обладающее гонадотропным действием (ГСЖК). Благодаря большой молекулярной массе оно не выводится почками и циркулирует в крови с 6-й по 20-ю неделю жеребости. Максимальная концентрация его на третьем месяце. Активность гонадотропина измеряется в мышиных (ме) или интернациональных (ИЕ) единицах. 1 ИЕ равна 2,5 ме, или 0,25 мг активного вещества. Мышиной единицей принято называть то наименьшее количество СЖК, которое способно вызвать признаки полового созревания у инфантильного белого мышонка весом 7-8 г.
Биологическое действие гонадотропина СЖК похоже на комбинированное действие ФСГ и ЛГ с преобладанием ФСГ. У неполовозрелых животных ГСЖК вызывает увеличение массы яичников, рост фолликулов, их лютеинизацию, иногда овуляцию и образование желтых тел. Это зависит от используемой дозы. В небольших дозах СЖК вызывает реакцию половых проводящих путей и рост фолликулов (реакция А) и только при значительном увеличении дозы стимулирует образование желтых тел, т. е. реакцию Б (М. М. Завадовский). Широко применяется для вызова суперовуляции у доноров эмбрионов, повышения многоплодия у овец, а также для стимуляции и синхронизации полового цикла у самок.
Половые гормоны – гестагены, андрогены, эстрогены – секретируются половыми железами, надпочечниками, плацентой. Они относятся к группе стероидных гормонов и имеют общую основную структуру. Деление половых гормонов на мужские и женские условно и связано с тем, что яичники вырабатывают в основном прогестерон и эстрогены, а семенники - тестостерон.
Прогестерон - наиболее распространенный гестаген – секретируется желтым телом, в небольших количествах – гранулезой и текой фолликулов перед овуляцией, надпочечниками и плацентой. Этот гормон действует на матку, подготавливает ее к беременности и обеспечивает нормальное ее протекание. Подготовка к беременности начинается- под влиянием эстрогенов. Прогестерон затем превращает эндометрий из проли-феративного в секреторный. Железы эндометрия становятся извитыми, активными и начинают выделять густой секрет, называемый "маточным молоком". Это способствует прикреплению и питанию зародыша. Шейка матки и влагалище выделяют густую слизь, которая закупоривает цервикальный канал. Прогестерон тормозит самопроизвольные ритмические сокращения миометрия, понижает чувствительность его к гормонам, химическим факторам и механическим воздействиям, способствует его гипертрофии. Таким образом обеспечивается растяжение матки растущим плодом.
Прогестерон тормозит развитие фолликулов и овуляцию, а также способствует развитию дольчато-альве-олярной системы вымени.
Прегненолон и прогестерон превращаются в андрогены: дегидроэпиандростерон, андростендион, тестостерон. Образуются они главным образом в семенниках интерстициальными клетками Лейдига и частично клетками Сертоли. Первые два гормона, биологически более слабые, секретируются в значительных количествах надпочечниками и плацентой. Оболочка фолликулов секретирует андростендион и в небольших количествах – тестостерон. Дегидроэпиандростерон и андростендион могут превращаться в коже, печени и других тканях в тестостерон. Таким образом обеспечивается высокая концентрация андрогенов в отдельных тканях без высокой концентрации тестостерона в крови. Это предотвращает его нежелательное общее воздействие на организм самки. Наиболее активный тестостерон в специфических тканях (предстательной железе и др.) превращается в еще более сильный метаболит – дегидротестостерон.
Андрогены контролируют сперматогенез, развитие и проявление вторичных половых признаков, а также оказывают анаболическое действие. Контроль за сперматогенезом заключается во влиянии тестостерона совместно с ФСГ на извитые канальцы семенника и образование жизнеспособных сперматозоидов. Развитие и функционирование вторичных половых органов (придатка семенника, спермиопровода, мочеиспускательного канала, придаточных половых желез) почти целиком зависит от тестостерона. Развитие вторичных половых признаков (рост волос, характерные телосложение и развитие голосовых связок, обусловливающее более низкий голос) является в основном следствием анаболических процессов, стимулирующихся андрогенами. Эти процессы приводят к накоплению азота и синтезу белка, формированию мышечной массы. Половое влечение (libido) у самцов и самок и агрессивность у самцов также обусловлены андрогенами.
В эндокринных органах и коже андрогены превращаются в эстрогены. Основные эстрогены: Е: - эстрон, Е2а- 17а - эстрадиол, Е2р- 17р - эстрадиол, ЕЗ - эстриол. Секретируются эстрогены яичниками, главным образом клетками внутренней оболочки и гранулезой, в которой содержатся ферментные системы, обеспечивающие превращение андрогенов в эстрогены. В небольшом количестве эстрогены продуцируются надпочечниками и семенниками (семенники жеребца секрети-руют большое количество гормона).
Главным из эстрогенов является эстрадиол 17„. Он образуется клетками внутренней оболочки фолликула. Здесь вырабатывается и небольшое количество эстрона. Эстрогены и другие стероидные гормоны накапливаются в фолликулярной жидкости. Максимальное количество их установлено между 9-м и 50-м часами перед овуляцией. Из фолликулярной жидкости эстрогены попадают к трубчатым половым органам. Это происходит за 6-30 ч до овуляции.
Эстрогены вызывают васкуляризацию и усиление кровообращения тканей половых органов, разрастание слизистых оболочек матки и влагалища, увеличение секреции слизи клетками влагалища и шейки матки, повышают активность желез матки, способствуют разрастанию миометрия, повышают тонус и усиливают спонтанные сокращения матки. Матка под влиянием эстрогенов становится более устойчивой к инфекции. Эстрогены обусловливают поведение животного, характерное для течки и охоты. Они играют важную роль в наступлении предродовых изменений в половых органах и в начале проявления сократительного потенциала миометрия, стимулируют анаболизм белка и развитие молочных протоков вымени. Чем больше половых циклов у телок к моменту оплодотворения, тем полнее развитие в их вымени молочных протоков и выше молочная продуктивность в первую лактацию.
В крови стероидные гормоны находятся в связанном с белками состоянии. Концентрируются они в специфических тканях: эстрадиол – в цитоплазме и ядрах клеток эндометрия, тестостерон — в предстательной железе. В плазме клеток органов-мишеней имеются специфические рецепторные белки, которые избирательно связываются с гормонами. Например, эстрадиол, имеющий наибольшее сродство с белком эндометрия, связывается с ним сильнее, чем эстрон и эстриол. После взаимодействия гормона с рецепторным белком происходит изменение структуры белковой части комплекса. После этого комплекс гормон - белок проникает в ядро клеток. Взаимодействуя с определенными участками хромосом, гормон активизирует синтез РНК. Одновременно повышается синтез других белков и развиваются процессы, характеризующие биологическое действие гормона на орган-мишень.
В регуляции воспроизводительной функции самок большая роль принадлежит простагландинам. Впервые на существование таких веществ указали гинекологи из Нью-Йорка Курзрок и Лиеб в 1930 г. Они заметили, что матка женщины реагирует сокращением или расслаблением в ответ на свежую сперму. В 1935 г. шведский ученый Эйлер показал, что тонизирующим действием на гладкую мускулатуру обладает липидный экстракт спермы человека, обезьян, овец и коз. Он предполагал, что это вещество вырабатывается в предстательной железе и назвал его простагландином.
Простагландины – это биологически активные липиды, которые содержатся в небольших количествах практически в каждой ткани тела. Биосинтез всех наиболее известных групп простагландинов (А, Б, Е и Ф) происходит из ненасыщенных жирных кислот – арахидоновой и линоленовой. Эти кислоты входят в состав клеточных мембран. Здесь с помощью специфических ферментов и осуществляется синтез простагландинов. Действие каждого из простагландинов порой несходно или даже противоположно. Так, простагландины Ег и Е2 повышают тонус и увеличивают амплитуду сокращений матки и расположенных ближе к ней участков яйцепроводов и одновременно расслабляют дистальный конец их. Тем самым они облегчают доступ спермиев к яйцеклетке и задерживают продвижение ее в полость матки, пока не наступит оплодотворение. ПГ-Е3 угнетает сокращения яйцепроводов и матки, а ПГ-Ф2α стимулирует их. Простагландины группы Ф наиболее широко используются в животноводстве.
У самок ПГ-Ф2α проявляет два важных действия. Во-первых, он усиливает сокращения яйцепроводов и матки, способствуя передвижению спермиев, а также участвует в механизме передвижения зигот. Наряду с окситоцином он играет существенную роль в родовом процессе. В отличие от окситоцина, к которому беременная матка нечувствительна, ПГ-Ф2α вызывает сильные сокращения ее и прерывает беременность, если в этот период накапливается в избытке. Во-вторых, ПГ-Ф2α обладает сильным лютеолитическим действием. Он выделяется эндометрием в значительных количествах в конце полового цикла и обусловливает наступление очередной охоты.
У крупного рогатого скота простагландины попадают из матки в яичник по местному маточно-яичнико-вому пути. Этот путь включает маточно-яичниковую вену и тесно прилегающую к ее стенке сильно извилистую яичниковую артерию. Простагландины из вены проникают в артерию и достигают по ней яичника, связываются с высоко сродственными к ним рецепторны-ми местами в желтом теле и тем самым нарушают стимуляцию лютеинизирующим гормоном аденилциклазы и синтез прогестерона.
У жвачных животных регрессия желтого тела происходит в результате эпизодического выделения ПГ-Ф2α из матки с интервалом около 6 ч. Выделение стимулируется окситоцином, секретируемым желтым телом. Поэтому каждое эпизодическое выделение про-стагландина связано с эпизодическим выделением окситоцина. Более того, простагландин стимулирует секрецию окситоцина яичником.
МЕХАНИЗМ ЕСТЕСТВЕННОГО РЕГУЛИРОВАНИЯ ПОЛОВОГО ЦИКЛА
Основная роль в механизме естественного регулирования воспроизводительной функции отводится центральной нервной системе (гипоталамусу), гормонам аденогипофиза и половых желез, простагландинам и ряду гонадальных факторов.
На центральную нервную систему оказывают влияние внешние (свет, температура, присутствие самца и др.) и внутренние (гормоны яичника) факторы (рис. 12). Они стимулируют или тормозят активность гипоталамуса. В цитоплазме нервных клеток ядер гипоталамуса секретируется гонадатропин-рилизинг-гормон. Под влиянием адекватных раздражителей ГнРГ из гипоталамуса через портальную систему кровообращения попадает в переднюю долю гипофиза и вызывает выделение ФСГ и ЛГ.
Эти гормоны, воздействуя на яичник, стимулируют рост и созревание фолликулов и секрецию эстрогенов. Примерно между 16-м и 19-м днями полового цикла у коров, на 12-14-й день у кобыл и на 15-й день у овец и свиней отмечается резкое уменьшение содержания прогестерона в периферической крови и увеличение эстрогенов. У коров и овец содержание эстрогенов достигает пика в начале стадии эструса и затем снижается до базального уровня к моменту овуляции; у кобыл пик гормона отмечается в течение эструса, а у свиней за 48 ч до начала эструса.
Эстрогены усиливают кровообращение и рост тканей наружных половых органов и влагалища, стимулируют секрецию слизи клетками влагалища и шейки матки (вызывают течку). Под их влиянием увеличивается активность миометрия и происходят изменения в эндометрии, которые являются началом подготовки его к приему зародыша. В промежуточном мозгу возникает очаг возбуждения. При участии гипоталамических центров и лимбической системы формируется половая мотивация, а при участии центра в коре больших полушарий — половая доминанта, т.е. состояние половой охоты.
В это время созревающие фолликулы (у
крупного рогатого скота – доминантный
фолликул) продуцируют возрастающие
количества ингибина, фолликулярного
регулирующего протеина (ФРП) и эстрадиола.
Ингибин и эстрадиол, воздействуя на
гипофиз, способствуют понижению секреции
ФСГ и таким образом блокируют появление
новых крупных фолликулов, а ФРП из
преовуляторных фолликулов угнетает
синтез эстрадиола второстепенными
фолликулами, что приводит к их атрезии.
это время созревающие фолликулы (у
крупного рогатого скота – доминантный
фолликул) продуцируют возрастающие
количества ингибина, фолликулярного
регулирующего протеина (ФРП) и эстрадиола.
Ингибин и эстрадиол, воздействуя на
гипофиз, способствуют понижению секреции
ФСГ и таким образом блокируют появление
новых крупных фолликулов, а ФРП из
преовуляторных фолликулов угнетает
синтез эстрадиола второстепенными
фолликулами, что приводит к их атрезии.
Следует отметить, что ингибин секретируется всеми полостными фолликулами и имеет более продолжительный период полураспада, чем эстрадиол, который продуцируется только преовуляторными фолликулами. Поэтому действие ингибина на выделение ФСГ более заметное, чем эстрадиола. В то же время резкое увеличение содержания эстрадиола в преовуляторных фолликулах обусловливает повышение чувствительности их к гонадотропинам крови и обеспечивает дальнейшее развитие до овуляции. На выделение ФСГ оказывают влияние и два других пептидных гормона, которые выделены из фолликулярной жидкости – активин (стимулирует выделение) и фоллистатин (угнетает секрецию гонадотропина).
У овец не ингибин, а эстрадиол является основным фактором в механизме регулирования уровня гонадотропинов в периферической крови. Резкое увеличение эстрогенов (эстрадиола) в начале эструса приводит к увеличению содержания ЛГ, пик которого наблюдается примерно за 14 ч до овуляции. В соответствии с этим отмечается и пик ФСГ, а второе его повышение наблюдается через 2 дня после овуляции.
Повышение содержания эстрадиола активизирует гипоталамус, который стимулирует выделение из передней доли гипофиза большого (овуляторного) количества ЛГ. У кобыл содержание ЛГ повышается постепенно и удерживается на относительно высоком уровне в течение 5-6 дней до и после овуляции. В ответ на резкое возрастание количества ЛГ в плазме усиливается рост клеток гранулезы фолликулов, начинается секреция прогестерона и высвобождение коллагенолитического фермента, истончающего стенку фолликула. Это приводит к разрыву ее под влиянием гидростатического давления фолликулярной жидкости. Обычно овуляция происходит спустя 12 ч после окончания охоты у коров, а у овец, свиней и кобыл до окончания охоты примерно за 6, 10-15 и 24 ч соответственно.
На месте овулировавшего фолликула образуется желтое тело, которое секретирует прогестерон. Быстрое увеличение содержания этого гормона вскоре после охоты угнетает секрецию ГнРГ и препятствует дальнейшему освобождению ЛГ. Овуляция новых фолликулов не происходит. Под влиянием прогестерона эндометрий полностью подготавливается к приему и прикреплению зиготы и обеспечивает нормальное протекание беременности. Выделение прогестерона желтым телом поддерживается у ряда лабораторных животных лютеотропным гормоном, а у коров, овец и свиней - ЛГ.
При отсутствии в матке зародыша эндометрий выделяет в конце полового цикла лютеолитический фактор (ПГ-Ф2α), который, действуя на яичник, вызывает регрессию желтого тела. Исчезновение основного источника прогестерона происходит на фоне наличия в яичниках новых растущих фолликулов. Рост их начинается сразу же после овуляции вследствие уменьшения уровня ингибина, ФРП и эстрадиола и последующего периодического увеличения ФСГ. У крупного рогатого скота первая после овуляции волна роста фолликулов заканчивается атрезией вначале мелких, а затем и самого крупного из них (доминантного) фолликула. Атрезия, в свою очередь, дает начало новой волне роста фолликулов. У этого вида животных в течение полового цикла имеют место две или чаще три таких волны роста фолликулов. В результате последней волны появляется доминантный овуляторный фолликул и происходит овуляция, завершается предыдущий и начинается новый половой цикл, а в случае успешного осеменения животного развивается беременность. У беременных животных лютеолитический эффект эндометрия устраняется присутствием эмбриона.
П ОЛОВОЙ
ЦИКЛ КОРОВЫ
ОЛОВОЙ
ЦИКЛ КОРОВЫ
Половая зрелость у телок наступает чаще в возрасте 8-10 мес. Осеменяют их в 16-18-месячном возрасте при достижении живой массы 350 кг. По экономическим соображениям целесообразно более раннее осеменение (с 13-15 мес). При этом развитие телок должно быть хорошим, а последующее кормление во время беременности обеспечить достаточный прирост и достижение безопасной для первого отела живой массы.
Коровы - полициклические животные. После наступления первой овуляции у телок половая цикличность проявляется более или менее регулярно во все сезоны года. Продолжительность полового цикла у них составляет в среднем 20 (от 18 до 22) дней. В случае недокорма половая цикличность прекращается. Чаще это можно наблюдать осенью и зимой в холодную погоду.
У коров после родов с 7-10-го дня отмечается рост фолликулов в яичниках, и в конце второй недели может произойти овуляция. В среднем интервал от отела до первой овуляции составляет 20-38 дней (от 14 до 40 дней). У многих коров первая послеродовая волна роста фолликулов не завершается овуляцией. Но она сопровождается кратковременным повышением в крови (молоке) содержания прогестерона (рис. 13). Нередко отмечается два или даже три таких подъема гормона. Погрешности в кормлении и содержании, частое доение и подсос, а также различные заболевания в послеродовой период тормозят проявление охоты и овуляции. Первый половой цикл короче, чем второй, что связано с недостаточной функциональной активностью первого желтого тела. Наличие в матке инфекции может укорачивать цикл до 10 дней. В среднем длина полового цикла у коров составляет 21 день. Нормальные колебания находятся в пределах 18-24 дней. Длительность стадий полового цикла: проэструса – 3-4 дня, эструса – 12-18 ч, метэструса – 3-4 и диэструса – 10-14 дней.
Половая охота у коров и телок продолжается в среднем 16 ч (от 10 до 23 ч), у большинства животных она длится 13-17 ч (В.С.Шипилов). Возможны колебания продолжительности охоты от 2,5 до 36 ч. У телок с возрастом половая охота удлиняется и признаки ее у них выражены более отчетливо, чем у коров. Продолжительность первой послеродовой охоты короче, чем двух последующих. В жаркие месяцы у лактирующих коров отмечается значительное укорочение охоты. У многих животных она проявляется утром или в вечернее время.
Нередко у телок первая овуляция не сопровождается внешними признаками половой охоты ("тихая овуляция"). У коров в первый послеродовой цикл частота "тихой овуляции" достигает 70% и более. После установления регулярных циклов у телок и в последующие циклы у коров частота их с хорошо выраженной охотой повышается. У животных с "тихой овуляцией" отмечаются обычные морфологические и функциональные изменения в половых органах, но характерные для половой охоты поведенческие реакции отсутствуют. Тщательным наблюдением таких животных можно распознать.
На протяжении полового цикла в яичниках коров наблюдается две или три волны роста фолликулов (рис. 14). Поэтому в них постоянно находятся фолликулы диаметром около 0,5 см. В течение охоты доминантный фолликул быстро увеличивается в диаметре до 1,5-2 см и свободно пальпируется через прямую кишку. Окончательное созревание его происходит после охоты. У телок крупные фолликулы часто образуют сферическую выпуклость над поверхностью яичника. У коров они обычно не выпячиваются и дают ощущение большого диаметра, не вызывая больших изменений в очертаниях яичника. В момент овуляции диаметр фолликула составляет 16-19 мм, но некоторые фолликулы, имеющие меньший диаметр, также могут разрываться. Овуляция может происходить в любом месте свободной поверхности яичника. В последующем в связи с развитием желтого тела форма яичника будет зависеть в основном от места овуляции.
У 4-5 % коров овулирует два фолликула. Около 60 % всех овуляций происходит в правом яичнике. Овуляция наступает через 12,5 ± 0,8 ч после окончания охоты или через 28,9 ± 0,7 ч после ее начала (B.C. Шипилов). У коров время овуляции может колебаться от 2 ч до окончания охоты до 26 ч после окончания (в среднем через 12,5 ч), у телок - от 2,2 до 22 ч после окончания, в среднем через 11 ч (Г. У. Солсбери, Н. Л. ВанДемарк). В 85% случаев овуляция происходит в вечерне-ночное время; наличие самца и акта спаривания ускоряет ее.
П
Рис. 14.
Модель развития доминантных фолликулов
в течение полового цикла у крупного
рогатого скота:
а
- стадии и основные явления в течение
полового цикла; б - секреция прогестерона
желтым телом; в - секреция эстрадиола
доминантными фолликулами; г - цикл
доминантных фолликулов осле
разрыва фолликула полость его уменьшается,
гранулеза и внутренний слой оболочки
образуют складки и почти целиком
заполняют полость. В ней остается лишь
небольшое количество фолликулярной
жидкости. Кровяных сгустков не бывает,
но на месте разрыва иногда появляется
небольшое кровоизлияние. Клетки гранулезы
превращаются в лютеальные клетки, и
через четыре дня после овуляции
формирование желтого тела в основном
завершается, секреция прогестерона
резко возрастает. К этому времени оно
имеет желтовато-коричневый цвет и его
можно пальпировать через прямую кишку.
Покровный эпителий полностью закрывает
овуляционную ямку. К 10-му дню заканчивается
васкуляризация желтого тела, оно
окончательно сформировано, становится
оранжевым или желтым. Диаметр его
достигает 2-2,5 см, масса - 4,1-7,4 г. В период
с 11-го по 17-й день цвет варьирует от
светло-оранжевого до светло-желтого. В
16-25% случаев желтое тело внутри имеет
небольшую полость, в которой содержится
желтоватая жидкость (рис. 15, 16).
осле
разрыва фолликула полость его уменьшается,
гранулеза и внутренний слой оболочки
образуют складки и почти целиком
заполняют полость. В ней остается лишь
небольшое количество фолликулярной
жидкости. Кровяных сгустков не бывает,
но на месте разрыва иногда появляется
небольшое кровоизлияние. Клетки гранулезы
превращаются в лютеальные клетки, и
через четыре дня после овуляции
формирование желтого тела в основном
завершается, секреция прогестерона
резко возрастает. К этому времени оно
имеет желтовато-коричневый цвет и его
можно пальпировать через прямую кишку.
Покровный эпителий полностью закрывает
овуляционную ямку. К 10-му дню заканчивается
васкуляризация желтого тела, оно
окончательно сформировано, становится
оранжевым или желтым. Диаметр его
достигает 2-2,5 см, масса - 4,1-7,4 г. В период
с 11-го по 17-й день цвет варьирует от
светло-оранжевого до светло-желтого. В
16-25% случаев желтое тело внутри имеет
небольшую полость, в которой содержится
желтоватая жидкость (рис. 15, 16).
По мере формирования желтое тело выпячивается из яичника. У телок оно часто сильно выступает из стромы яичника и прикреплено к ней только основанием. У коров желтое тело нередко полностью погружено в ткань яичника, и его положение можно обнаружить по выступающей овуляционной точке. У большинства желтых тел к моменту их максимального роста эта точка расположена в виде углубления в возвышении диаметром около 1 см. Некоторые желтые тела возвышаются в виде сосочка, но иногда место возвышения их диффузно разлито (рис. 16).
Наличие желтого тела в яичнике изменяет его размеры. Как правило, этот яичник значительно больше по величине, чем другой. Хорошо заметна эта разница у телок.
У ровень
секреции прогестерона желтым телом
достигает максимума через 7-8 дней после
овуляции и быстро понижается с 18-го дня
(см. рис. 13, 14). За 24-30 ч до следующей охоты
желтое тело, как правило, начинает быстро
уменьшаться в величине, цвет его
становится желтовато-оранжевым. В период
охоты диаметр желтого тела уменьшается
до 1,5 см, а к середине полового цикла
составляет около 0,5 см, и выступающая
часть его немного больше булавочной
головки.
ровень
секреции прогестерона желтым телом
достигает максимума через 7-8 дней после
овуляции и быстро понижается с 18-го дня
(см. рис. 13, 14). За 24-30 ч до следующей охоты
желтое тело, как правило, начинает быстро
уменьшаться в величине, цвет его
становится желтовато-оранжевым. В период
охоты диаметр желтого тела уменьшается
до 1,5 см, а к середине полового цикла
составляет около 0,5 см, и выступающая
часть его немного больше булавочной
головки.
В течение полового цикла заметным
изменениям подвергаются эпителиальные
клетки передней части влагалища и
секретирующие клетки шейки матки. В
середине цикла клетки влагалища варьируют
от уплощенных до низкоцилиндрических.
В стадии проэструса происходит размножение
и рост высоких цилиндрических
слизеобразующих поверхностных клеток.
В эструс этот процесс усиливается, что
приводит к сильному утолщению эпителия
передней части влагалища.
течение полового цикла заметным
изменениям подвергаются эпителиальные
клетки передней части влагалища и
секретирующие клетки шейки матки. В
середине цикла клетки влагалища варьируют
от уплощенных до низкоцилиндрических.
В стадии проэструса происходит размножение
и рост высоких цилиндрических
слизеобразующих поверхностных клеток.
В эструс этот процесс усиливается, что
приводит к сильному утолщению эпителия
передней части влагалища.
За 12-24 ч до появления первых признаков охоты начинается обильная секреция слизи из передней части влагалища и шейки матки. Количество слизи увеличивается в течение охоты и затем постепенно уменьшается к 4-му дню после охоты. В это время слизь содержит хлопья из лейкоцитов. Лейкоцитарная инвазия слизистой оболочки влагалища максимальная через 2-5 дней после охоты. Эластичность слизи варьирует, она максимальная во время охоты. В связи с эластичностью изменяется и способность слизи к кристаллизации. Наиболее выражена кристаллизация во время охоты и в течение 2 последующих дней, а в другое время отсутствует. Под микроскопом в мазках кристаллы слизи расположены в виде листка папоротника. Этот феномен (арборизация), так же как количество и свойства слизи, зависит от уровня эстрогенов. В начале охоты слизь клейкая, прозрачная, светлая, а к концу охоты становится более вязкой. Время наименьшей вязкости слизи совпадает с временем максимального ослизнения слизистой оболочки преддверия влагалища и минимального электрического сопротивления покрывающей эту оболочку слизи. Наряду с изменением внешних свойств изменяется структура и химический состав, а также рН слизи. В передней части влагалища за день до охоты показатель рН составляет 6,7-7,0, но затем уменьшается в начале охоты и к моменту овуляции опускается до минимального значения (6,5-6,6), затем быстро увеличивается до 7,0 и достигает 7,2-7,4 на 4-5-й день и 7,3-7,5 на 11-12-й день.
Циклические изменения в матке и во влагалище контролируются гормонами. Под влиянием эстрогенов в период течки и охоты прогрессирующе усиливается кровообращение половых органов, увеличивается сеть кровеносных сосудов, т. е. происходит васкуляризация. Матка кровенаполнёна, ее слизистая оболочка блестящая и покрыта отечной жидкостью. После овуляции уровень эстрогенов быстро падает и оказывается недостаточным, чтобы поддержать такое состояние сосудов. Наполненные кровью, они разрываются (главным образом в области карункулов), и небольшое количество крови выходит в матку. Кровь смешивается со слизью и выделяется из половых органов. У некоторых животных через 40-48 ч после начала охоты наблюдается выделение ярко-красной слизи. Спустя 3-5 дней после окончания охоты резко снижается интенсивность кровообращения в половых органах, слизистая оболочка влагалища и матки становится бледной, канал шейки матки закрыт, слизи немного, она вязкая, светло-желтая или коричневатая.
С началом созревания фолликула мышечные
волокна матки растут в длину. Матка
становится большей по величине. Происходят
изменения и в характере ее сокращений.
В стадии эструса сокращения с большей
амплитудой, не частые (рис. 17). В ответ
на пальпацию через прямую кишку матка
реагирует сильным сокращением. Способность
к сокращениям (ригидность) повышена за
день до охоты, в период охоты и на
следующий день после охоты; в середине
цикла сокращения более слабые и частые.
началом созревания фолликула мышечные
волокна матки растут в длину. Матка
становится большей по величине. Происходят
изменения и в характере ее сокращений.
В стадии эструса сокращения с большей
амплитудой, не частые (рис. 17). В ответ
на пальпацию через прямую кишку матка
реагирует сильным сокращением. Способность
к сокращениям (ригидность) повышена за
день до охоты, в период охоты и на
следующий день после охоты; в середине
цикла сокращения более слабые и частые.
Половая охота - строго специфическая реакция самки на самца и ее можно точно установить только рефлексологическим методом, т. е. используя самца-пробника. Однако у коров и телок при контакте с другими самками проявляются такие характерные для охоты признаки, которые позволяют практически безошибочно распознать ее. Знание этих признаков и умение их использовать позволяют животноводам успешно организовать искусственное осеменение коров и телок без использования быков-пробников. Наиболее важными являются следующие признаки.
1. Еще до начала охоты корова обнюхивает и облизывает соседних коров, особенно их половые органы, а также подошедших близко людей; при этом спина спереди от .таза выгибается, а затем сильно прогибается, хвост поднимается и опускается. Такое движение можно вызвать поглаживанием поясницы или дотра-гиванием до вульвы. Этот признак выражен у большинства животных. Иногда корова сильно изгибается и пытается облизать свои половые органы.
2. Слизистая оболочка преддверия влагалища припухшая и гиперемирована, влажная и покрыта прозрачным секретом, который часто вытекает из половой щели, поэтому пучок волос в нижнем углу вульвы всегда бывает влажным.
3. Постоянно переступает с ноги на ногу, а находясь в группе коров, пытается приблизиться к другим животным в охоте (или не в охоте) и вспрыгнуть на них, иногда даже на человека.
4. Корова в охоте проявляет признаки общего возбуждения, чаще стоит. Если кто-нибудь проходит сзади, она поднимает голову и провожает проходящего взглядом.
5. Глаза более живые, часто неподвижные и блестящие зрачки расширены.
6 у некоторых коров охота сопровождается громким
мычанием в течение нескольких часов или всего дня.
7 У многих животных наблюдается снижение удоя, но оно может пройти незамеченным; более обычным является затрудненное доение, так как корова задерживает молоко. Снижается также руминация.
Если корова находится в стаде, признаки охоты выпажены более отчетливо. Животные в охоте в 2-3 раза увеличивают двигательную активность (регистрируемую с помощью педометров), следуют за другими животными и вспрыгивают на них, независимо от того, в охоте эти животные или нет. Наибольшая активность прыгать проявляется в конце проэструса, в эструс и в начале метэструса. Если замечено, что корова повторяет прыжки, следует продолжить наблюдение за ней, так как возможно наступление охоты. Однако этот признак не является определяющим. Он может наблюдаться и у животных, не находящихся в охоте.
8 Более важным для выявления состояния охоты является поведение коровы в момент, когда на нее вспрыгивает другое животное: если она стоит спокойно или слегка уклоняется, это указывает на наличие у нее охоты. В разгар охоты у коров проявляется, как правило, только такой типичный признак (рефлекс неподвижности) и лишь изредка - активный обнимателъный рефлекс, но без совокупительных движении. У животных в это время хорошо выражены и специфические изменения в наружных половых органах. Слизь вытекает в виде длинной вязкой нити, приклеивается к ягодицам и хвосту. Если животное лежало, возле его таза можно увидеть скопление мутноватой или голу6оватого цвета слизи. У телок наблюдается и четкий подъем температуры во влагалище.
У животных с "тихой" охотой эти изменения также имеют место. Отмечается у них и возбужденное состояние, рев, попытки вспрыгивать на других коров. Но сами они не допускают садки других животных и не являются особо "привлекательными". При осеменении таких животных может произойти оплодотворение. При контакте .коровы в охоте с быком животные
облизывают друг друга, после чего корова пытается вспрыгнуть на быка прежде, чем допускает садку сама. Если корова в охоте долгое время находилась в стаде, то вследствие вспрыгивания других животных у нее стирается волос и появляются ссадины на коже крестца. Эти признаки сохраняются 4-6 дней. Но если ссадины влажные, с выделением экссудата или кожа при дотрагивании болезненна и имеет повышенную температуру, это говорит о том, что корова в охоте или охота только что прошла.
Иногда коровы в охоте следуют за другими коровами, с фырканьем обнюхивают их половые органы, опираются головой на поясницу, поднимают хвост и помахивают им. Нередко за такими коровами активно следуют другие животные.
Наблюдается повторяемость особенностей проявления и длительности половых циклов у одного и того же животного в различные годы.
Охоту у коров выявляют на основании проявления у них специфических признаков. Целесообразно трехкратное в день наблюдение по 20-30 мин до раздачи корма во время движения животных. При беспривязном содержании животные должны быть помечены бирками или ошейниками с хорошо заметными номерами. На крупных фермах наблюдение можно проводить с помощью телеустановок.
С целью более полного и своевременного выявления охоты у коров и определения оптимального времени их осеменения используют быков-пробников, а также специально подготовленных коров-выявительниц или натренированных собак, детекторы охоты различных конструкций, электрометрические приборы и т. д.
ПОЛОВОЙ ЦИКЛ ОВЦЫ
Половая зрелость у ярочек наступает в возрасте 5-8 мес. Осеменяют их на втором году жизни. При хорошем развитии самки возможно и более раннее покрытие.
Овцы — полициклические животные с выраженным половым сезоном. У диких овец половой сезон совпадает с осенними месяцами, у одомашненных пород продолжительность его колеблется в зависимости от климатических условий, кормления и содержания, породных особенностей животных. У тонкорунных овец, разводимых на юге Украины и Северном Кавказе, половая цикличность проявляется со второй половины сентября по март. Но, если в июле-августе выпадает достаточно осадков, а температура воздуха невысокая, то взрослые овцы начинают приходить в охоту со второй декады августа, а переярки и полутарогодовалые ярки – с конца июля. В Беларуси овцы могут проявлять половую цикличность в период с июля до марта.
Сезоннность в размножении овец регулируется факторами внешней среды и, главным образом, суточным количеством света. Начинается половой сезон с уменьшением светового дня и продолжается в течение всего короткого дня. Уменьшение света стимулирующим образом действует на гипофиз, повышая его гонадот-ропную активность. Это действие осуществляется через гормон эпифиза - мелатонин.
После родов охота может наступить в первый месяц, но это зависит от сезона ягнения. После весеннего ягнения охота наступает медленнее и значительно быстрее - после осеннего. Повышение содержания прогестерона в организме регистрируется после 50-го дня.
Длительность стадий полового цикла: проэструса – 2-3 дня, эструса – 24-38 ч, метэструса – 2-3 дня и диэструса – 10-12 дней.
Половая охота у овец проявляется менее ярко, чем у других видов, и определить ее у большинства животных без барана-пробника практически невозможно. Сопровождается охота выделением специфических пахучих веществ – аттрактантов (феромонов), которые легко воспринимаются самцами. Выключение зрительной рецепции не дезориентирует баранов: руководствуясь обонянием, бараны и в таких случаях безошибочно находят овец в состоянии охоты. Продолжительность охоты 24-38 ч (от 12 до 72 ч). У старых маток охота длится меньше, чем у молодых. В начале полового сезона охота короче, чем в разгар сезона. В прохладные месяцы охота продолжается дольше, чем в теплые.
Быстрое увеличение и созревание фолликулов начинается после наступления охоты. В это время один или более фолликулов достигают 0,7-1 см в диаметре.
Количество фолликулярной жидкости в них увеличивается, оболочка истончается и покрывается сеточкой капилляров. Фолликулярная жидкость кажется пурпурного цвета. Перед овуляцией на поверхности фолликула появляется небольшой сосочек, который и разрывается. Овуляция у большинства животных (80%) происходит через 31-32 ч после начала охоты, у остальных – через 22- 30 ч. При множественной овуляции второй фолликул разрывается обычно через 1-2 ч, но не позднее чем через 4 ч после разрыва первого фолликула. У многоплодных романовских овец промежуток между овуляциями может быть и больше 4 ч.
На 5-й день после овуляции образовавшееся желтое тело имеет в диаметре 0,6 см и достигает максимальной величины (0,7-0,9 см) к середине цикла. Цвет изменяется от кровянисто-красного до бледно-розового; внутри имеется центральная полость. Регрессия желтого цвета начинается на 15-й день; цвет изменяется вначале до желтоватого, затем до коричневато-желтого.
Течка у овец непродолжительна, и обнаружить ее трудно. Проявляется она гиперемией слизистых оболочек, увеличением числа слоев вагинального эпителия и ороговением поверхностных клеток, иногда точечными кровоизлияниями, повышением тонуса мускулатуры матки, усилением секреции слизи клетками шейки матки. Слизь в начале охоты прозрачная, слегка опалесцирующая, затем становится более мутной и в конце охоты приобретает салообразную консистенцию.
В отаре овцы, находящиеся в охоте, стучат копытами, помахивают хвостом, стремятся приблизиться к производителю, иногда группой ходят за бараном. Овца считается в охоте, если спокойно допускает садку барана.
У коз много общего с овцами в проявлении половой функции. Но продолжительность цикла у них несколько больше (19-21 день). Охота длится 22-33 ч. Овуляция происходит через несколько часов после охоты.
ПОЛОВОЙ ЦИКЛ СВИНЬИ
Первый половой цикл у свиньи наблюдается в возрасте 5-7 мес. К моменту первой овуляции половые органы еще недостаточно подготовлены к зачатию, число овулирующих фолликулов небольшое, отмечается задержка в передвижении спермиев к яйцепроводам. Через 2-3 половых цикла (с 8,5-месячного возраста) свинок можно осеменять.
После опороса во время лактации (подсоса) охота обычно не наступает; проявляется она только через 4-7 дней после отъема поросят. При раннем отъеме период до наступления охоты удлиняется, увеличивается частота кист яичников. При отъеме после 25-30 дней фактор времени оказывается малосущественным. У свиноматок с низкой молочностью и длительным периодом лактации охота может наступить спонтанно. Имеют место случаи наступления охоты и через 2-6 дней после опороса, но овуляция до 9-го дня не происходит. В промышленных хозяйствах целесообразно отнимать поросят на 28-32-й день. При этом достигается определенное согласование с первой естественной подпороговой эстрогенизацией организма вследствие активизации яичников на 16-й день после опороса. Отъем затем приходится на стадию диэструса, и после него у большинства свиноматок можно ожидать наступления охоты в течение недели. Повышением уровня кормления свиноматок после отъема поросят ускоряется наступление охоты, увеличивается число овулирующих фолликулов, оплодотворяемость после осеменения и численность помета.
Дикие свиньи - моноциклические животные. В условиях Беловежской пущи половая активность у них проявляется в ноябре-декабре. У всех домашних свиней после наступления половой зрелости или первой послеродовой охоты половые циклы повторяются регулярно во все сезоны года. Продолжительность их составляет в среднем 20-21 день. Возможны колебания от 11 до 42 дней. Проэструс длится 3-4 дня (иногда до 6 дней), эструс – 2-3 дня, метэструс – 2-3 дня, диэструс – 11-13 дней. У молодых свинок проэструс продолжительнее, эструс короче, чем у взрослых свиноматок. Длительность половой охоты (время допуска хряка) составляет 40-60 ч (от 12 до 120 ч). Первая охота короче, чем последующие. У старых маток она продолжительнее, чем у молодых. Существует индивидуальная повторяемость длительности охоты.
В яичниках свиней одновременно созревает и овулирует 10-20 фолликулов (иногда до 40). Описан случай массивной спонтанной овуляции (97 фолликулов) у помесной свиноматки (крупная белая х ландрас).
Рост фолликулов начинается в стадии проэструса. Во время охоты они достигают 8-10 мм в диаметре и к моменту овуляции становятся выпуклыми, мягкими на ощупь, пронизаны сетью кровеносных сосудов. В них можно заметить место будущего разрыва. Это лишенный кровеносных сосудов, просвечивающийся участок оболочки фолликула. Овуляция начинается через 30-36 ч и длится 4-7 ч. После овуляции в яичниках остаются фолликулы диаметром 0,4 см, которые к 18-му дню цикла постепенно увеличиваются до 0,9 см.
После разрыва полость фолликула спадается, но по мере образования кровяного сгустка приобретает коническую форму. Клетки гранулезы гипертрофируются, формируется желтое тело. К третьему дню оно имеет темно-красный цвет, затем изменяется до светло-красного; функционирует до 15-го дня. Кровяной сгусток к шестому дню замещается соединительнотканной пробкой или светло-желтоватой жидкостью. Пробка может сохраняться до 12-го, а жидкость – до 18-го дня. В процессе регрессии цвет желтого тела изменяется от светло- красного до желтого, затем до кремово-желтого. К середине диэструса следующего цикла желтые тела превращаются в маленькие светло-желтые тела, а еще через цикл выглядят в виде сероватых пятнышек.
В период созревания фолликулов вульва (петля) набухает и приобретает красный цвет, слизистая оболочка влагалища становится отечной, розоватого или ярко-красного цвета. Из половой щели может выделяться слизь. В начале охоты слизь светлая, к концу охоты беловатая. Выделяется слизи немного. В мазках из вагинальной слизи в период эструса отмечается феномен арборизации. Реакция слизи во влагалище слабокислая (рН 6,61), в шейке матки – слабощелочная (рН 7,49), в матке и яйцепроводах – нейтральная. Четкие изменения вульвы в период эструса наблюдают только у 75% самок. Набухание и покраснение ее редко сохраняется до конца охоты и обычно исчезает во время спаривания или до этого.
За 3—12 ч до наступления стадии эструса происходят изменения в поведении свиньи. Она становится беспокойной, часто мочится, издает характерное хрюканье, мало ест, обнюхивает половые органы других животных, вспрыгивает на них и может позволять делать садку на себя другим свиньям. Повышается "разведывательная" активность свиньи, т.е. она пытается выбраться из станка. Некоторые самки активно преследуют хряков за день до наступления эструса. В этой стадии явления течки и охоты усиливаются. Характерным признаком готовности свиньи к спариванию является рефлекс неподвижности. В присутствии самца она настораживает уши, выгибает дугой спину и застывает в такой позе; никакие подталкивания и удары не могут заставить ее сойти с места. Если в это время положить ей на крестец руку или сесть верхом, то она не уклоняется, а продолжает стоять спокойно. Но без хряка такая реакция наблюдается только у половины животных.
Главными раздражителями рефлекса неподвижности являются обонятельные и слуховые. Внешний вид самца тоже может иметь значение. Однако наблюдения показывают, что если самка не видит хряка, но слышит хрюканье и воспринимает его запах, то влечение к нему ослабляется незначительно. Записанное на магнитофон хрюканье хряка вызывает рефлекс неподвижности более чем у половины свинок, которые не проявляли рефлекса без хряка. Очень сильны и обонятельные раздражители. Свыше 60% свинок в охоте, которые не реагируют на надавливание рукой, проявляют неподвижность при переводе их в станок хряка. Запах жидкости, взятой из препуция хряка и подогретой до температуры тела, дает такой же эффект, как и запах самого хряка. Реакция зависит от стероидного вещества, вырабатываемого в препуции и придающего характерный запах свинине. Оно синтезировано и может быть использовано при выявлении охоты (препараты суидор, феромаксеин и др.). Достаточно 0,1-1 мг стероидного вещества смешать со 100 г инертного вещества и распылить или разбрызгать его в свинарнике, чтобы стимулировать половую активность у свиноматок в охоте. Спустя 1-2 мин после применения его более половины свиней, которые не проявляли рефлекса неподвижности без хряка, давали положительную реакцию в виде рефлекса неподвижности. Предлагаются механические чучела, в которых размещаются устройства для воспроизведения звуковых и обонятельных раздражителей (немецкая фирма Шипперс ГмБх).
Наиболее точный способ выявления охоты у свиней - это использование хряка-пробника в группе из 4-8 самок. Целесообразнее помещать в контрольный станок первым хряка, чтобы он привык к окружающей обстановке, а затем вводить маток. Если хряк обладает хорошей половой активностью, то бывает достаточно 5-минутного контакта с каждой самкой.
При выявлении охоты у свиней животноводы должны учитывать и такие признаки, как повышенную возбудимость, потерю аппетита, характерное хрюканье, припухание и покраснение вульвы и др. Отдельно взятый признак не является надежным показателем охоты, но по их совокупности опытный работник, знающий всех животных индивидуально, легко может определить самок в охоте. Прогон хряка по проходу свинарника, где содержатся матки, существенно облегчает эту задачу.
ПОЛОВОЙ ЦИКЛ КОБЫЛЫ
Половая зрелость у кобыл наступает обычно во второе лето их жизни; осеменяют в 3-4-летнем возрасте.
Кобылы — полициклические животные. Половая охота у них может наступить в любое время года, особенно при наличии достаточного количества корма, при содержании в теплых светлых помещениях. Однако чаще половая активность проявляется с февраля по июль-август. Затем следует длительный период анэс-труса, в течение которого функция яичников подавлена. Они небольшие, бобовидной формы и только иногда могут содержать фолликулы диаметром до 1,5 см.
В конце зимы начинается переходный период к регулярной половой цикличности. Созревание фолликулов в это время идет медленно, и они обычно не пальпируются через прямую кишку. Продолжительность охоты не характерна (до 1 мес. и более), признаки ее атипичны, неясно выражены и с трудом могут быть замечены. Как правило, такая длительная весенняя охота не заканчивается овуляцией. Часто это бывает в холодные весны, а также у очень молодых и старых слабоупитанных кобыл. После первой овуляции половая цикличность становится регулярной.
После выжеребки первая половая охота наступает на 8-10-й день (от 5 до 14 дней) и поэтому нередко покрытие приурочивают к этому времени (на 9-й день). Продолжительность этой охоты короче - 2-4 дня. Наиболее же типичная продолжительность ее 6 дней. Продолжительность полового цикла - 21-23 дня, хотя возможны колебания от 7 до 37 дней. Первые два цикла короче, а в разгар полового сезона их продолжительность довольно стабильная - около 22 дней (6 дней – охота и 16 дней – промежуток между концом одной и началом другой охоты). Проэструс длится 2-3 дня, эструс – 4-8 дней, метэструс – 2-3 дня и диэструс – 10-12 дней. Овуляция происходит в конце охоты, обычно за 1-2 дня до окончания ее. В большинстве случаев овулирует один фолликул. Частота двойных овуляций составляет 18-27%. В 52,2% случаев овуляция происходит в левом яичнике (Н. Arthur, 1975).
У кобыл двухфазное пульсаторное выделение
ФСГ с промежутком в 10-12 дней. Один пик
гормона наблюдается вскоре после
овуляции, а второй в середине или конце
диэструса, приблизительно за 10 дней до
следующей овуляции. Эта повышение
стимулирует развитие новой генерации
фолликулов. Рост и созревание их
становится заметным в фазе проэструса;
диаметр увеличивается до 1-3 см. Усиливается
кровоснабжение половых органов, слизистая
оболочки влагалища становится слегка
гиперемированной, блестящей. Слизи
мало, она непрозрачная, соломенно-желтого
цвета. В стадии эструса уже в первый
день один из фолликулов увеличивается
до 2,5-3,5 см в диаметре. В последующие дни
увеличение еще более заметно. К моменту
овуляции в яичнике кобылы может находиться
2-3 фолликула диаметром до 3-7 см, а также
другие фолликулы меньшего диаметра
(рис. 18). Эти изменения приводят к сильному
увеличению массы яичников.
кобыл двухфазное пульсаторное выделение
ФСГ с промежутком в 10-12 дней. Один пик
гормона наблюдается вскоре после
овуляции, а второй в середине или конце
диэструса, приблизительно за 10 дней до
следующей овуляции. Эта повышение
стимулирует развитие новой генерации
фолликулов. Рост и созревание их
становится заметным в фазе проэструса;
диаметр увеличивается до 1-3 см. Усиливается
кровоснабжение половых органов, слизистая
оболочки влагалища становится слегка
гиперемированной, блестящей. Слизи
мало, она непрозрачная, соломенно-желтого
цвета. В стадии эструса уже в первый
день один из фолликулов увеличивается
до 2,5-3,5 см в диаметре. В последующие дни
увеличение еще более заметно. К моменту
овуляции в яичнике кобылы может находиться
2-3 фолликула диаметром до 3-7 см, а также
другие фолликулы меньшего диаметра
(рис. 18). Эти изменения приводят к сильному
увеличению массы яичников.
Перед овуляцией напряжение в фолликулах падает, а через прямую кишку они пальпируются в виде флюктуирующих образований. Вскрываются в области овуляционной ямки.
У кобыл не отмечается предовуляционного пика ЛГ, как у других животных. Содержание гонадотропина постепенно повышается и поддерживается на высоком уровне в течение 5-6 дней до и после овуляции.
После овуляции неовулировавшие крупные фолликулы регрессируют в течение первых 4-9 дней или же подвергаются лютеинизации. На месте овули-
ровавшего фолликула отмечается кровоизлияние, и в течение 24 ч образуется сгусток. В первые три дня лютеиновая масса пальпируется в виде эластичного участка, а затем консистенция ее не отличается от других тканей яичника. Максимальной величины желтое тело достигает на 4-5-й день. Оно не выступает над поверхностью яичника и имеет вначале коричневый, затем желтый цвет. Форма коническая или треугольная, вершина направлена к овуляционной ямке. В центре его находится различное количество темно-коричневого фибрина. Функционирует желтое тело цикла до 14-го дня, после чего начинается функциональная и морфологическая регрессия. Завершается обратное развитие его через 7 недель.
Вследствие гормонального влияния яичников в стадии эструса усиливаются васкуляризация и гиперемия половых органов, канал шейки матки приоткрывается и в него можно ввести сначала один, а затем и два пальца. Шейка матки укорачивается по длине и принимает по отношению к влагалищу центральное положение. Количество слизи увеличивается, она становится более водянистой и прозрачной. К концу этой стадии слизь приобретает мутноватый цвет и вязкую консистенцию. Признаки охоты хорошо выражены, вульва набухает, отмечается мигание половой щели. При этом кобыла открывает и закрывает половую щель, поднимает хвост, выпячивает клитор и испускает небольшое количество мочи и слизи. Во время охоты кобылы сильно возбуждены, непослушны, часто ржут. Услышав или увидев жеребца, они принимают позу для спаривания. Во время садки жеребца стоят спокойно.
В стадии метэструса признаки течки и охоты угасают, животное не допускает садки. Уменьшается отечность вульвы, исчезает гиперемия слизистых половых органов, в яичнике формируется желтое тело.
В стадии диэструса у кобыл слизистая оболочка влагалища бледная, часто желтоватого цвета, покрыта небольшим количеством вязкой, мутноватой слизи. Шейка матки закрыта. Поверхность яичников гладкая, чаще бобовидной формы, в них находят фолликулы диаметром до 1,5 см.
Для выявления половой охоты без жеребца-пробника используют акустические (звуковое подражание заигрывания жеребца) и тактильные стимулы (пальпация области холки, боков и половых органов). Они позволяют определить состояние эструса (спокойное поведение, поднятие хвоста, мигание половой щели и выделение слизи, раскидывание задних конечностей) или диэструса (нервозность, лягание и ржание).
ПОЛОВОЙ ЦИКЛ СУКИ
Собаки – моноциклические животные. Половой цикл у них бывает два раза в году с интервалом около 7 мес., чаще в январе-марте и августе-сентябре. Возможно проявление половой активности в другое время года. У самок мелких пород половые циклы бывают чаще. Молодые самки, достигшие хорошего развития к осени, проявляют свой первый цикл в этот же сезон.
Проэструс у сук длится 9-10 дней (от 4 до 14 дней), клинически хорошо выражен. Характеризуется кровянистыми истечениями из половых органов и прогрессирующим припуханием половых губ. Припухание начинается через несколько дней после появления истечений, интенсивность которых достигает максимума в ранний проэструс и сохраняется на таком уровне до начала охоты. Истечения имеют специфический запах и улавливаются самцами на большом расстоянии. У молодых сук вследствие сильного отека слизистой оболочки иногда наблюдается выпадение влагалища. В течение большей части проэструса самки привлекательны для самцов, но не допускают на себя садку.
Эструс продолжается 9 дней (5-12 дней). Кровянистые истечения в начале этой стадии уменьшаются в количестве, но могут продолжаться на протяжении всей охоты. Более часто, однако, они становятся светлыми или желтоватого цвета. Отечность и припухание вульвы достигают максимума к началу охоты. Половые губы становятся мягкой консистенции. Отечность их может сохраняться даже в первое время метэструса.
С завершением проэструса или за день до этого поведение самки изменяется. Она начинает проявлять интерес к самцу, хотя еще и не готова к спариванию. С началом эструса желание к коитусу уже очевидно и вскоре сука принимает позу для спаривания. Наиболее сильно половая активность проявляется в первые два дня эструса, хотя побуждение для самца к коитусу сохраняется до конца охоты.
Общая продолжительность двух стадий достигает 21 дня. При удлинении проэструса укорачивается эструс, и наоборот.
В стадии метэструса происходят изменения эндометрия, характерные для беременности (псевдобеременность). Длится эта стадия до 8 недель и иногда завершается ложными родами с последующей лактацией. Затем наступает длительный период анэструса, который продолжается до начала нового полового цикла в следующем половом сезоне. Продолжительность анэструса около 3 мес.
В период полового покоя вагинальный эпителий низкий цилиндрический, кубоидального типа, представлен двумя или тремя слоями. В мазках из вагинальной слизи в это время содержатся неороговевшие эпителиальные клетки и небольшое количество нейтрофилов. В течение проэструса эпителиальные клетки изменяются до высоких, чешуйчатых, слоистого типа и сохраняются в таком состоянии до позднего проэструса. Слой ороговевших клеток и непосредственно лежащие под ним слои эпителиальных клеток теряются путем дес-квамации в течение проэструса. А в стадии эструса остаются низкие, плоские клетки, которые превращаются в цилиндрические эпителиальные в первые 1-3 недели после окончания охоты. В стадии метэструса (и беременности) эпителиальные клетки более высокого цилиндрического типа, чем при анэструсе. В соответствии с изменением вагинального эпителия в течение цикла изменяется и содержание мазков. В проэструсе появляется много эритроцитов, происходит постепенное увеличение больших ороговевших клеток и прогрессирующее исчезновение лейкоцитов. В начале эструса в слизи находятся, главным образом, большие ороговевшие клетки и эритроциты. В конце эструса эритроциты разрушаются и вскоре исчезают. Вновь появляются нейтрофилы и вместе с малыми эпителиальными клетками они доминируют в мазках в первые 2—3 недели метэструса.
Появление крови в вагинальной слизи в проэструсе связано с диапедезом эритроцитов через неповрежденный эпителий матки, но возможно в незначительном количестве и повреждение кровеносных сосудов.
На протяжении анэструса яичники овальной или слегка уплощенной формы. У самок средней величины длина их 1,4 см, ширина - 0,8 см. Видимых фолликулов не содержат. На разрезе заметны лишь небольшие остаточные желтые тела предыдущего цикла в виде желтоватых или коричневатых точек. В начале проэструса развивающиеся фолликулы достигают диаметра 0,5 см. К моменту овуляции они увеличиваются до 0,6-1 см. В это время яичники крупнее, причем степень увеличения их зависит от числа созревающих фолликулов и их расположения. В процессе созревания стенка фолликулов утолщается и это усложняет возможность отличить их от желтых тел. Перед овуляцией в фолликуле появляется сосочек величиной с булавочную головку. Эпителий на нем становится коричневатого цвета, что контрастирует с остальной светлой частью фолликула. Характерный признак зрелых фолликулов у собак - утолщение его стенки, связанное с гипертрофией и складчатостью гранулезы. Овуляция происходит спонтанно через сутки после начала охоты. Все фолликулы овулируют в течение короткого периода времени. Формирующиеся желтые тела содержат вначале центральную полость, но по мере лютеинизации клеток она заполняется в течение 10 дней. К этому моменту желтое тело достигает величины 0,6-1 см в диаметре; цвет его желтовато-розовый. У небеременных сук оно сохраняется до 30-го дня после овуляции, затем начинает медленно регрессировать. У беременных самок желтое тело функционирует до конца беременности и быстро регрессирует после родов.
СПОСОБЫ ИСКУССТВЕННОГО РЕГУЛИРОВАНИЯ ПОЛОВОЙ ФУНКЦИИ
По мере постижения естественного механизма нервно-гормональной регуляции воспроизводительной функции у самок разрабатывались и методы искусственного ее контроля. Были предложены эффективные гормональные средства для стимуляции функции яичников и синхронизации половой охоты, вызова суперовуляции у одноплодных животных, индуцирования родов и лечения гинекологических заболеваний. Наиболее стабильные результаты получены путем применения гормонов гипоталамуса и гипофиза, половых гормонов, простагландинов и других биологически активных веществ. Эффективными оказались и способы целенаправленного воздействия факторов внешней среды на животных с целью повышения воспроизводительной функции.
ГОРМОНАЛЬНЫЕ ПРЕПАРАТЫ
Из гормонов гипоталамуса наиболее широкое применение имеют синтетические аналоги ГнРГ (сурфагон, диригестран, фертирелин, гонадорелин, бусерилин и др.). При парентеральном введении они вызывают выделение ЛГ, уровень которого в периферической крови повышается уже через 30 мин и достигает максимума через 2-3 ч. Выделение больших количеств ЛГ стимулирует процесс овуляции.
Сурфагон (Россия, Латвия) выпускается в виде стерильного раствора в ампулах и флаконах емкостью 2,5, 10, 20, 100 мл, содержащих в 1 мл 5, 10 и 15 мкг вещества. Минимальная фармакологическая доза для коров, вызывающая выделение гипофизом ЛГ и ФСГ – 10 мкг, терапевтическая – 10-50 мкг; для свиней – 10 мкг, овец – 2,5 мкг.
Диригестран выпускается в ампулах по 5 мл; в 1 мл содержится 40 мкг гонадорелина. Доза коровам – 100-250 мкг АДВ (2,5-6,25 мл препарата), свиноматкам – 100 мкг (2,5 мл препарата).
Оба эти препарата применяются для "запуска" половой цикличности у высокопродуктивных коров после отела и при гипофункции яичников, для лечения животных с, кистами яичников; для индукции овуляции у коров и телок с естественной или вызванной простагландином половой охотой, а также у коров-доноров эмбрионов; при задержке овуляции и для повышения функциональной активности желтого тела. Животным с выраженной половой сезонностью их можно применять для стимулирования половой цикличности в период анэструса. Вводят их внутримышечно.
Фертирелин (Ovalyse) и гонадорелин (Fertagyl) – синтетические пептидные препараты ГнРГ. Фертирелин применяют коровам в дозе 100 мкг, гонадорелин – 0,5 мг (внутримышечно). Бусерилин (Receptal) – синтетический пептидный аналог ГнРГ. Применяют коровам в дозе 10-20 мкг, кобылам – 40 мкг (внутримышечно). Все эти препараты применяют: коровам при фолликулярных кистах, отсутствии и задержке овуляции; при анэструсе (отсутствии половой цикличности, эффективна одна инъекция – доза); для снижения частоты эмбриональной смертности – одна инъекция бусерилина на 12-й день после осеменения; кобылам для индукции овуляции (одна или несколько инъекций).
Гормон задней доли гипофиза окситоцин используется в ветеринарной акушерской практике в виде синтетического полипептидного препарата (Oxytocinum), препарата гипофиза крупного рогатого скота, содержащего окситоцин и вазопрессин – питуитрина (Pituit-rinum) и комбинированного препарата, содержащего питуитрин и экстракт молочной железы лактирующих коров, – маммофизина (Mammophysinum). Окситоцин выпускается в ампулах и флаконах по 1, 2, 5, 10, 20 и 50 мл; в 1 мл содержится 5 ЕД. Доза окситоцина (внутримышечно) коровам и кобылам 30-60 ЕД, свиноматкам массой до 200 кг до 30 ЕД, овцам и козам 10-15 ЕД и собакам 5-10 ЕД.
Эти препараты используются как средства, усиливающие сокращения матки: для стимуляции родового процесса (у кобыл и свиней; свиньям целесообразно вводить многократно малые дозы – 1-2 мл, дозы выше 10 ЕД вызывают спазм матки); при задержании последа, эндометрите, атонии и субинволюции матки; для ускорения инволюции матки после патологических родов, кесарева сечения и вправления выпавшей матки; при маточной травме и геморрагиях и т.д.; перед осеменением для повышения оплодотворяемости; при задержке молокоотдачи, а также при лечении животных с заболеваниями молочной железы, когда требуется полное освобождение полости от секрета. У сук при травме матки и геморрагиях экстракты гипофиза нельзя использовать.
В Англии питуитарный экстракт выпускается в виде препарата гипостона.
Гормоны передней доли гипофиза – ФСГ и ЛГ являются основой многих биотехнологических способов контроля воспроизводительной функции у животных. Наиболее известные препараты ФСГ: FSH-P (США), фолликотропин Спофа (Чехия), ФСГ-СУПЕР (Российская Федерация), фоллитропин (Литва), ФСГ (Беларусь) и др.; препараты ЛГ: овогон-ТИО (Российская Федерация) и др.
Препараты фолликулостимулирующего гормона высокой степени очистки (хроматографической) получают из передней доли гипофизов домашних животных (обычно свиньи). Представляют собой лиофилизи-рованную массу белого или серовато-желтого цвета, хорошо растворимую в физиологическом растворе или дистиллированной воде. Вводят их внутримышечно.
FSH-P выпускается во флаконах вместимостью по 10 мл. В картонную упаковку помещают два флакона. В одном флаконе содержится FSH-P 50 мг по Арморовскому стандарту, в другом растворитель (10 мл 0,9%-ного раствора натрия хлорида).
ФСГ-СУПЕР имеет оптимальное для коров соотношение гонадотропинов: 1000-1500 ед. ФСГ/1 ед. ЛГ. Выпускается во флаконах по 5 мл. Один флакон содержит 50 ед. Арморовского стандарта (1000 ИЕ).
ФСГ расфасован в пенициллиновые флаконы по 20 мл массой 75-150 мг, активностью 500-1200 ИЕ. Обладает высокой активностью при оптимальном соотношении ФСГ и ЛГ.
Фолликотропин выпускается во флаконах по 40 ME (2 мг). Упаковка содержит 5 флаконов препарата и 5 флаконов растворителя.
Фоллитропин выпускается во флаконах с активностью не менее 300 ЕД.
Эти препараты применяют для вызывания суперовуляции у коров-доноров эмбрионов и индукции половой охоты у животных с гипофункцией яичников. Доза на одну корову-донора: 42 мг по Арморовскому стандарту (FSH-P и ФСТ-СУПЕР), 24 мг (12 флаконов фолликотропина Спофа), 1100 ЕД фоллитропина или 1200 ИЕ ФСГ-Б. Молоко от коров-доноров эмбрионов используют без ограничений.
Овогон-ТИО – препарат лютеинизирующего гормона, изготовленный из передней доли гипофизов крупного рогатого скота. Представляет собой лиофилизированную аморфную массу белого или серовато-желтого цвета. Выпускается в ампулах с активностью 750, 1000 и 1500 ИЕ. Перед применением овогон-ТИО растворяют стерильным физиологическим раствором в объеме 2,0 мл. Применяют препарат для синхронизации суперовуляции у коров-доноров эмбрионов и для нормализации половой функции у животных (при отсутствии овуляции и при многократных неплодотворных осеменениях, а также при фолликулярных кистах). Коровам-донорам эмбрионов овогон-ТИО вводят дважды через 52 и 60 ч после первого введения простагландина в дозе 1500 ИЕ. Для повышения оплодотворяемости при повторных осеменениях овогон-ТИО вводят в дозе 1000-1500 ИЕ за час до первого искусственного осеменения. При фолликулярных кистах яичников препарат вводят в дозе 3000 ИЕ.
Гонадотропин хорионический (хориогонин, human chorionlc gonadotrophin, hCG) получают из мочи беременных женщин. Он стимулирует секрецию половых гормонов в яичниках и семенниках, а также превращение андрогенов в эстрогены в плаценте; вызывает овуляцию зрелых фолликулов и образование желтых тел, выпускается во флаконах в виде стерильного лиофилизированного порошка по 500, 1000 и 1500 ЕД (1 ЕД = 0,1 мг стандартного вещества). В Англии – аналогичный препарат хорулон (Chorulon) и LH 1500, в Италии – профази. Эти препараты применяются животным для синхронизации овуляции после предварительной обработки препаратами прогестерона или ФСГ, а также для ускорения овуляции при задержке или отсутствии ее; для лечения животных с фолликулярными кистами; при недостаточности желтого тела; для улучшения результатов осеменения у многократно осеменяемых коров. Сукам применяют для сокращения проэструса/эструса и лечения кист яичника; кошкам – для индукции овуляции. Самцам применяют для стимулирования libido. Доза коровам и кобылам 1500-3000 ЕД (внутривенно, внутримышечно); свиньям – 500-1000 ЕД (внутримышечно или подкожно); овцам и козам – 100-500 ЕД (внутривенно или внутримышечно); собакам – 100-500 ЕД (внутримышечно); кошкам – 100-200 ЕД (внутримышечно).
Хорионик гонадотропин (маслянистая суспензия для инъекций), в 1 мл хорионического гонадотропина содержится 200 IU и 1,0 мг эстрадиола бензоата. Применяется свиноматкам для синхронизации половой охоты и оплодотворения, при задержке охоты, для вызова овуляции, предупреждения беременности и стимулирования оплодотворения. Доза – 2-3 мл. Охота обычно наступает через 2-5 дней.
Нимфалон (Nymfalon) — хорионический гонадотропин с прогестероном. Применяется в тех же целях, что и хорионический гонадотропин.
Гонадотропин менопаузный (human menopausal, hMG) получают из мочи женщин, находящихся в пост-менопузе. Обладает фолликулостимулирующим действием. По своим свойствам аналогичен зарубежному препарату пергонал-500. Выпускается во флаконах в виде стерильного лиофилизированного порошка по 75 ЕД. Применяется для вызывания суперовуляции у коров-доронов эмбрионов и стимулирования функции яичников при анэструсе. Вводят внутримышечно в дозе: коровам 1000-1500 ЕД.
Гонадотропин СЖК (equine chorionic gonadotrophin, eCG) применяется в виде нативной сыворотки и очищенных препаратов: фоллигона, фостима(Росит, Англия), гонадотропина сывороточного, гравотаджа и ово-гена (Российская федерация), прегмогона (Германия) и др.
Фоллигон – высокой степени чистоты гонадотропин СЖК. Выпускается во флаконах по 1000 или 5000 ИЕ; в отдельных флаконах помещается растворитель. Применяется для стимулироавния половой цикличности при анэструсе, повышения оплодотворяемости и вызова суперовуляции у коров-доноров эмбрионов. Доза коровам-донорам (внутримышечно) от 1500 до 3000 ИЕ, для стимулирования половой функции – 500-1000 ИЕ, сукам - 500 ИЕ, овцам – 400-700 ИЕ.
Половые гормоны, естественные и синтетические, представляют наиболее многочисленную группу биологически активных веществ и широко применяются в медицине и ветеринарной медицине.
Прогестерон кристаллический получают синтетически из сапогенинов растений, растворим в этиловом спирту и растительных маслах. Для подкожных и внутримышечных инъекций применяется в виде 1- или 2,5%-ного масляного раствора, расфасованного в ампулы по 1 мл. Как и эндогенный гормон, синтетический прогестерон задерживает наступление течки и охоты и может быть использован для синхронизации половых циклов, предупреждения гибели зародышей и сохранения беременности, для лечения животных с заболеваниями яичников. Доза коровам (внутримышечно) 60-125 мг, свиньям – 30-40 мг.
PRID (progesterone-releasing intravaginal device) – прогестеронвыделяющее внутривлагалищное устройство в виде спирали; содержит 1,55 г прогестерона и дополнительно 10 мг эфира эстрадиола, который обладает слабым лютеолитическим действием. Используется для синхронизации охоты у коров и телок (предпочтительнее в комплексе с простагландином), при истинном анэструсе (гипофункции яичников). Вводится на 12 дней во влагалище. За 24 ч до извлечения устройства делается инъекция простагландина. Охота наблюдается через 2-5 дней после извлечения устройства.
Внутривлагалищное прогестеронвыделяющее устройство EASI-BREED "CIDR" содержит 1,9 г прогестерона; вводится во влагалище на 7-12 дней. Простагландин инъецируют в день извлечения устройства. Применяется в тех же целях, что и PRID.
Предложены и другие эффективные препараты из группы синтетических прогестинов: оллитренболон (Ollytrenbollone, Regumate Equine), альтреногест (Altrenogest, Regumate Porcine), норгестамет (Crestar), флуорогестона ацетат (интравагинальные пессарии, Chronogest) и мегестрола ацетат (таблетки Ovarid), мед-роксипрогестерона ацетат (таблетки Perlutex tablets, для инъекций Perlutex for injection) и др.
Оллитренболон- жидкая субстанция, содержит 2,2 мг вещества в 1 мл. Применяется кобылам для стимулирования половой цикличности, контроля сроков проявления или задержки охоты в случной сезон. Вводится с кормом в дозе 27,5-33 мг в течение 10 или 15 дней. Охота наступает в течение 8 дней после последней дачи, овуляция – через 7-13 дней.
Альтреногест – суспензия, содержит 4 мг вещества в 1 мл. Применяется для синхронизации половой охоты у свинок. Помещается на корм непосредственно в момент кормления животных. Скармливается в дозе 20 мг (5 мл) в течение 18 дней. Охота наступает через 2-3 дня.
Крестар, Synchro-Mate В system включает имплантант (длина 1,9 см, диаметр 3 мм; содержит 3 мг синтетического прогестерона норгестамета) и 2 мл жидкости, содержащей 3 мг норгестамета и 5 мг эстрадиола бензоата. Этот препарат применяют коровам мясных пород и телкам. Лактирующим коровам, молоко которых используется в пищу людям, применять нельзя. Имплантант вводят под кожу снаружи в области корня уха, а жидкую часть препарата – внутримышечно. Инъецированный прогестерон действует немедленно и предотвращает овуляцию зрелых фолликулов, а имплантированный гормон тормозит созревание фолликулов в течение последующих 9 дней; эстрадиол способствует регрессии желтого тела, если оно имелось в яичниках в начале обработки. На 9-й день имплантант извлекают путем надреза кожи кончиком скальпеля.
Флуорогестона ацетат (Chronogest, интравагинальные пессарии), медроксипрогестерона ацетат (Veramix, Veramix Plus, интравагинальные пессарии) используются для синхронизации полового цикла у овец и коз, чаще в комбинации с ГСЖК. Пессарии вводят в переднюю часть влагалища на 12-14 дней, гонадотропин инъецируют в момент извлечения пессариев.
Медроксипрогестерона ацетат (Perlutex, таблетки) применяют для прерывания или задержки полового цикла у сук и кошек. Сукам дают препарат в дозах 10-20 мг в течение 4 дней с момента появления кровянистых выделений, затем по 5-10 мг в последующие 12 дней; для задержки цикла - ежедневная дача в дозах 5-10 мг. Дозы для кошек – 2,5 мг.
Медроксипрогестерона ацетат (Perlutex, для инъекций) применяют для задержки эструса у сук и предупреждения гиперплазии предстательной железы у кобелей. Сукам инъецируют подкожно 50-150 мг в период анэструса; самцам – 50-100 мг каждые 3-6 мес.
Депо-Промон (Медроксипрогестерона ацетат) применяют собакам и кошкам для задержки полового цикла. Инъецируют подкожно во вторую половину анэструса по 1 мл с 6-месячными интервалами у собак и с 4-месячными – у кошек.
Пролигестон для инъекций (Covinan) применяют для прекращения или задержки эструса у сук и кошек. Сукам в начале проэструса вводят подкожно в дозе 100-600 мг, кошкам в начале проэструса или эструса – 100 мг. В таких же дозах используется этот препарат в конце периода анэструса для непродолжительной задержки полового цикла или же с интервалом в 3, 4, а затем 5 мес для задержки цикла в течение длительного периода.
Применение прогестинов у сук и кошек может предрасполагать к развитию пиометры.
Эстрон (фолликулин) – естественный эстрогенный гормон, выпускается в масляном растворе в ампулах по 1 мл (10000 ЕД). Три других естественных эстрогена - эстрадиола бензоат и эстрадиола дипропионат, а также эстрадиола валерат – обладают более выраженным, чем у эстрона, пролонгированным действием. Выпускаются в ампулах по 1 мл 0,1%-ного раствора. Все эти препараты надо вводить парентерально (внутримышечно). Активность естественных эстрогенов выражается в интернациональных единицах. Одна ИЕ равна 0,1 мкг стандартного эстрона или мышиной единице, или 1/3 крысиной единицы.
Агофоллин содержит в 1 мл эстрадиола дипропио-ната 1 мг. Выпускается во флаконах по 15 мл.
Из синтетических эстрогенов наиболее известны диэтилстильбестрол и синестрол. Диэтилстильбестрол обладает эстрогенным действием, в два раза превосходящим действие синестрола и фолликулина. Диэтил-стильбестрола пропионат или дипропионат - синтетические эстрогенные препараты пролонгированного действия.
Синтетические эстрогены обладают таким же биологическим действием, как и естественные эстрогены, но они не разрушаются в пищеварительном тракте и поэтому могут применяться внутрь. В настоящее время в ряде стран они не используются, так как наличие остаточных количеств их в продуктах питания придает им канцерогенные свойства.
При введении в организм эстрогены вызывают течку с ее обычными внешними признаками, но при отсутствии овуляции. У крупного рогатого скота особо высокая чувствительность к эстрогенам. После введения их течка проявляется через 12-48 ч. После осеменения в период стимулированной течки и охоты оплодотворение не происходит. Овуляция, а затем проявление нормальной половой цикличности возможны только, если в яичниках в момент применения гормона имелись созревающие фолликулы. В таких случаях эстрогены стимулируют выброс овуляторных количеств ЛГ и овуляцию, которая могла быть и без их применения.
Рекомендуются эстрогены для стимулирования "созревания" шейки матки у кобыл перед применением окситоцина; при консервативном лечении задержания последа и эндометрита (кроме острого токсического) у коров в комплексе с другими гормональными препаратами или лекарственными средствами; лечения misalliance в течение первых 4 и предотвращения беременности – в течение 5 дней после спаривания, а также недержания мочи у овариэктомированных сук; гиперплазии простаты у кобелей; понижения гиперсексуальности у кобелей. Дозы кобылам – 3-6 мг, коровам – 3-5 мг; сукам при misalliance – 0,1 мг/кг массы (не более 3 мг), для предотвращения беременности – 5-10 мг (вызывает отек слизистой оболочки яй-цепроводов и задержку продвижения зигот), при недержании мочи – 1 мг ежедневно в течение 3 дней, а затем каждые 3 дня; кобелям при гиперплазии простаты 1мг в день.
Андрогены естественные или синтетические аналоги их применяются редко. Наиболее известные препараты: метилтестостерон (Orandron, таблетки) тестостерона (фенил) пропионат для инъекций (Androject) и тестостерона эфир для инъекций (Durateston). Пропионат тестостерона в 2-3 раза активнее метилтестос-терона и обладает более продолжительным действием. Выпускают его в виде масляного раствора по 1 мл в ампуле в концентрации от 0,5 до 5 %. В малых или физиологических количествах тестостерон у самцов усиливает половое влечение и оказывает благоприятное влияние на сперматогенез. Но в дозах, превышающих физиологическую норму, блокирует секрецию гонадотропинов и вызывает прекращение сперматогенеза. У самок введение тестостерона угнетает лактацию. Применяют андрогены самцам при половом недоразвитии и функциональных нарушениях полового аппарата, а также при сосудистых и нервных расстройствах. Андрогены обладают анаболическим эффектом, поэтому могут быть использованы при лечении ослабленных животных. Дозы согласно наставлениям по применению.
Из группы простагландинов для ветеринарного применения рекомендованы следующие препараты: энзап-рост-Ф (Венгрия), в состав которого входят ПГ-Ф2α, ацетат натрия и фенол. Выпускается в ампулах по 1 или 5 мл, действующего вещества по 5 или 20 мг, упакован в открываемых коробках по 1 или 5 ампул. Вводится внутримышечно в дозах: крупному рогатому скоту и свиньям по 20 мг, лошадям и собакам – по 5 мг. Аналогичный препарат динолитик/лутализ содержит в 1 мл раствора 5 мг динопроста. Коровам и телкам вводится в дозе 25 мг (5 мл), свиноматкам – 10 мг (2 мл), кобылам – 5 мг (1 мл).
Синтетические лютеолитические препараты, содержащие клопростенол: эструмат, планах (Estrumate, Planate), эстрофан, ремофан (Чехия, Лечива); эстуфа-лан, анипрост, клатрапростин, магэстрофан (Россия); суперфан (Латвия); биоэстрофан (Польша) и другие выпускают в ампулах по 2 мл (упакованы в коробки по 10 ампул) или флаконах по 20 мл с содержанием действующего вещества от 60 до 250 мкг/мл. Применяют внутримышечно крупному (500 мкг) и мелкому (125-250 мкг) рогатому скоту, лошадям (125-500 мкг), свиньям (до 350 мкг).
Лупростиол (Luprostiol, Prosolvin, Англия) также применяют этим животным в дозах: крупному рогатому скоту – 15 мг, лошадям – 7,5 мг, овцам и козам, свиньям – 7,5 мг.
Тиапрост (Tiaprost, Iliren) применяют только свиньям в дозе 300-600 мг. Широко используются в медицине и ветеринарии для стимуляции и синхронизации родов у животных, при лечении больных с гинекологическими заболеваниями и для регулирования половой цикличности.
СПОСОБЫ РЕГУЛЯЦИИ ПОЛОВОГО ЦИКЛА
У крупного рогатого скота вследствие проявления половой цикличности в течение всего года имеется возможность широкого применения гормональных препаратов. Показания к применению их возникают часто. У многих высокопродуктивных коров секреция гонадотропных гормонов после отела подавлена, замедляется инволюция половых органов. Все это создает предпосылки к возникновению послеродовых заболеваний, нарушению функции яичников и задержке первой половой охоты. Нередки случаи прекращения половой цикличности после проявления одной-двух охот или в другие периоды. Умение подобрать подходящий препарат и оптимальное время для его применения во многом предопределит успех в предупреждении расстройств функции половых органов, а также в регулировании сроков проявления половой охоты и осеменения, если, конечно, будут учтены и другие обстоятельства (баланс энергии, упитанность животного, эндокринный статус и др.).
Для обеспечения своевременного проявления половых циклов после отела и повышения воспроизводительной способности высокопродуктивных коров целесообразно применить ГнРГ (сурфагон). Вводить его можно с 9-10-го дня в дозе 50 мкг однократно или в виде пяти инъекций по 10 мкг через 10-12 ч (Г. Ф. Медведев, Д. С. Долина). Эффективно введение препарата в эти же сроки и животным с замедленным процессом инволюции матки. Для повышения функциональной активности желтого тела сурфагон вводят на 8-10-й день после осеменения в дозе 10 мкг. В такой же дозе применяют его в первые 2-4 ч после начала охоты для синхронизации овуляции.
Для регулирования сроков проявления половой охоты применяют гестагены, простагландины и гонадотропные гормоны, нередко в комбинации с эстрогенами. При введении гормональных препаратов группе животных обеспечивается наступление охоты одновременно (синхронно) у большинства из них. При этом требуется меньше времени на выявление охоты. Больше возможностей появляется для улучшения организации искусственного осеменения и сокращения сроков его проведения. После осеменения можно разместить животных в одном помещении (загоне), обеспечить кормление и содержание в соответствии с физиологическим состоянием и затем наладить квалифицированный контроль за течением родов. Сезон для родов может быть выбран по желанию.
Наиболее целесообразна и эффективна синхронизация половой охоты у мясного скота. Одна обработка и затем осеменение в течение одного-двух дней дают возможность максимально использовать все преимущества искусственного осеменения. В молочном скотоводстве синхронизация половой охоты имеет как преимущества, так и недостатки. У нетелей более трудные роды, что требует тщательного контроля во время их приема. Так как продолжительность периода отелов при осеменении в синхронизированную охоту существенно сокращается, контроль за течением родов осуществить проще. С другой стороны, при раздое первотелок необходимо больше приложить усилий и напряжения и с группой животных успешно справиться труднее. Кроме того, некоторые владельцы не желают одновременно вводить в стадо большое количество первотелок.
При введении в организм экзогенного прогестерона в течение длительного периода (16-19 дней) задерживается наступление течки и охоты, но желтое тело регрессирует, половые центры сенсибилизируются к эндогенным или введенным извне гонадотропинам. После прекращения введения гестагена следует периодическое выделение ГнРГ, который вызывает выделение гонадотропинов и через 2-6 дней проявляется охота. Наступление ее можно ускорить введением ГСЖК.
Медроксипрогестерон (МАП) и хлормадинон (КАП) скармливают телкам и коровам мясных пород в течение 18дней, ацетат мегестрола (меленгестррола ацетат) в течение 14 дней в дозах соответственно 200 мг, 10-12 мг и 35-45 мг. После курса обработки на протяжении 3 дней охота наступает у 60-80% телок, однако оплодотворяемость у них после первого осеменения обычно на 15 % ниже, чем в контроле. Это связано с нарушением соотношения прогестерон-эстрогены. Чтобы избежать этого, целесообразнов начале обработки дополнительно ввести эстрогены. Эстрогены, кроме того, ускоряют регрессию желтого тела посредством воздействия на матку, поэтому дачу прогестинов (мегестрола ацетата) можно сократить до 9 дней. В таких случаях на второй день обработки инъецируют 5 мг эстрадиола бензоата (дипропионата) или эстрадиола валерата. Осеменение после комплексной обработки так же эффективно, как и в естественную охоту. Проводят осеменение обычно в выявленную охоту (в течение 5 дней), но у ряда животных овуляция может наступить без признаков охоты. Поэтому нередко сочетают осеменение в выявленную охоту и в фиксированное время (через определенное время после прекращения дачи препарата): животных, которые не проявили охоту, осеменяют в фиксированное время. Однако при этом необходимо учитывать состояние яичников, матки и влагалища. Осеменять следует только животных с хорошо выраженной ригидностью матки, имеющих в яичниках крупные фолликулы и проявляющих признаки течки.
Норгестамет вводят (имплантируют) подкожно снаружи в области корня уха в дозе 6 мг на 8-12 день с последующей инъекцией простагландина за 24 ч до извлечения имплантанта. В начале обработки дополнительно делают инъекцию эстрадиола валерата (5 мг). После удаления имплантанта охота наступает через 24-120 ч (чаще через одни-двое суток) у 80% или более животных. Осеменяют в фиксированное время через 48 и 72 ч. Нередко простагландин в такую схему обработки животных не включают. Крестар и Synchro-Mate В system имплантируют на 10 или 9 дней. В первый день инъецируют дополнительно 3 мг норгестамета и 5 мг эстрадиола бензоата. Осеменяют животных также через 48 и 72 ч после извлечения трансплантанта. По данным Wishart and Drew (1977), процент стельности составил 66,2 при осеменении через 48 и 60 ч, 62,1 -через 48 и 72 ч и 65,7 – при однократном осеменении через 54 ч.
Внутривлагалищно прогестины вводят в виде металлической спирали, покрытой импрегнированной прогестероном силиконовой трубкой (внутривлагалищ-ные прогестеронвыделяющие устройства – PRID, EASI-BREED "CIDR") или специальных пессариев, пропитанных 3 г прогестерона или 200 мг флюорогестона ацетата (кронолона) с помощью специального влагалищного зеркала (расширителя). Слизистая оболочка влагалища абсорбирует прогестерон из устройства и в организме поддерживается высокая концентрация его (как во время диэструса) в течение всего периода обработки. PRID вводится на 12 дней; иногда дополнительно ецируется эстрадиола бензоат. EASI-BREED "CIDR" вводится во влагалище на 7-12 дней. После извлечения устройства охота наступает через 2-3 дня. Осеменяют животных в фиксированное время через 57 и 74 ч.
Более высокие результаты синхронизации и оплодотворяемость обеспечиваются при введении спирали на 13-14-й день цикла, по сравнению с 2-3-м днями. Целесообразно за 24 ч до извлечения устройства сделать инъекцию простагландина. В таких случаях обеспечивается синхронизация охоты у 100% животных. Пессарии вводятся во влагалище на 9-10 дней. Одновременно с использованием пессариев инъецируют прогестерон и эстрогены.
Более распространенным средством для регулирования полового цикла у крупного рогатого скота в Республике Беларусь являются простагландины. Они быстро вызывают регрессию желтого тела, т.е. устраняют источник прогестерона также эффективно, как и механическое удаление его из яичника рукой через прямую кишку (энуклеация) или хирургическим путем. При отсутствии прогестерона начинается рост и созревание фолликулов и наступает охота.
Энуклеация желтого тела рекомендовалась в тех случаях, когда у коровы или телки после отела или очередной охоты желтое тело функционировало дольше, чем обычно, несмотря на отсутствие беременности. Операция требовала высокой квалификации специалиста. И хотя она относительно безопасна, однако при грубом проведении ее возможны спайки яичника с яичниковым карманом и другие осложнения, которые в последующем приводят к снижению воспроизводительной функции животного. Энуклеация успешно может быть заменена введением ПГ-Ф2а или его синтетического аналога клопростенола. После внутримышечной инъекции 25 мг ПГ-Ф2α или 120-500 мкг клопростенола (2 мл суперфана, клатрапростина, эстрофана, ремофана, эстуфалана, анипроста или др.) коровам и телкам на 5-16-й день полового цикла происходит регрессия желтого тела, а через 2-3 дня у животных проявляются признаки течки и охоты. Овуляция наступает через 70-96 ч. Процент плодотворных осеменений в стимулированную охоту такой же, как и в естественную.
До 5-го дня желтое тело не чувствительно к простагландину, а с 17-18-го дня начинается его естественная регрессия. Наиболее стабильные результаты достигаются при введении простагландина с 8-го по 15-й день. По данным Watts T. L. (1983), после инъекций ПГ-Ф2а на 5-7-й день (содержание прогестерона в плазме крови 2,78 нг/мл) охоту проявили 43,0% телок, из них оплодотворилось 56,8% (от всех 25,4%); на 8-11-й день (прогестерона 5,18 нг/мл) реагировало 83,7% животных, оплодотворилось 62,1% (от всех 51,9%); на 12-15-й день (уровень гормона 5,22 нг/мл) реагировали все телки, оплодотворилось 78,3% .
В группы для синхронизации необходимо подбирать хорошо развитых телок с живой массой 350 кг или более (в отдельных случаях в конце стойлового периода можно включать и животных с массой 330-340 кг). Их клинически исследует и устраняют стельных (простагландин может вызвать аборт!) животных и с инфантильными половыми органами. Синхронизацию охоты у отобранных животных проводят согласно нескольким схемам.
1. Отобранным телкам делают две инъекции простагландина с промежутком в 11 дней. К моменту второй инъекции у всех животных в яичниках будут иметься чувствительные к простагландину желтые тела, образовавшиеся после естественной или стимулированной первой инъекцией препарата овуляции. После второго введения ПГ в результате регрессии желтого тела у всех животных могут проявиться признаки очередной охоты. Так как овуляция происходит почти одновременно, можно осеменять их без выявления охоты в строго определенное (фиксированное) время двукратно через 72 и 96 ч или однократно через 76-80 ч.
Результаты синхронизации и осеменения во многом зависят от состояния половых органов телок. Наиболее высокая оплодотворяемость после двукратного осеменения была у животных, у которых к моменту первой инъекции эстрофана обнаруживались в яичниках желтое тело (61,4%) или крупный фолликул (61,1%), ниже – у животных с нормальными по величине яичниками (48,3%) и самая низкая – с уменьшенными яичниками (39,0%). Высокая оплодотворяемость (66,6%) достигалась и при осеменении телок, у которых матка в момент первой инъекции препарата проявляла ригидность, хотя яичники имели небольшую величину (Г.Ф.Медведев, 1986).
Нестабильность результатов осеменения в фиксированное время может быть связана и с другими факторами, в том числе и с недостаточной синхронностью овуляции у различных животных. Для коррекции времени овуляции можно вводить сурфагон (10 мкг) в первые 2-4 ч после проявления охоты или через 72 ч после инъекции простагландина. Этот аналог ГнРГ целесообразно инъецировать также в начале естественной охоты или за 10-15 мин до первого осеменения "проблемным" животным.
2. Всем отобранным телкам инъецируют простагландин. В течение 2-4 дней осеменяют животных, проявивших в это время охоту. Неосемененным животным делают вторую инъекцию простагландина через 11 дней после первой. Осеменяют после выявления охоты. Животных, оставшихся неосемененными, осматривают через 76-80 ч и при наличии признаков течки осеменяют. После второй инъекции в фиксированное время можно осеменять всех животных.
3. Из всех отобранных телок простагландин вводят только тем, у которых в яичниках имеется хорошо сформированное желтое тело. После инъекции простагландина наблюдают за животными и при выявлении охоты осеменяют. Телок, не проявивших охоту, осматривают через 76-80 ч и при наличии признаков течки также осеменяют. Осеменяют в течение первых 4 дней и животных в естественную охоту. Неосемененным после первой инъекции животным делают вторую инъекцию простагландина через 11 дней. В это же время исследуют повторно оставшихся интактных телок и при наличии в яичниках желтого тела вводят простагландин. Осеменяют по мере выявления охоты или же в фиксированное время.
4. В течение первых 4-5 дней осеменяют телок в естественную половую охоту. На 5-й или 6-й день клинически исследуют неосемененнных телок и синхронизируют охоту согласно второй или третьей схеме.
При осеменении телок в фиксированное время необходимо учитывать степень проявления течки. Осеменять целесообразно только тех животных, у которых изменения в половых органах хорошо выражены (отечность половых губ, сильная гиперемия и овлажнение слизистой оболочки преддверия влагалища), истечение слизи в момент осеменения. Оплодотворяемость в таких случаях близка к стандартной - 57,1-63,4%.
Для достижения высоких результатов синхронизации охоты и оплодотворяемости телок важно правильно организовать эту работу. При беспривязном содержании животных в приспособленных помещениях, разделенных на несколько секций или загонов или же без них, до начала синхронизации оборудуют станок (можно несколько станков) и с обеих сторон к нему пристраивают расколы. Вблизи станка выбирают подходящую площадку (хорошо освещенный тамбур), где можно расположить инструменты для осеменения, устройство для оттаивания спермы и сосуд Дъюара. Для клинического исследования, введения простагландина и осеменения телок перегоняют в одну часть помещения, а затем поочередно пропускают через станок в другую часть. При этом необходимо максимально осторожно обращаться с животными, так как развитие стрессового состояния значительно снижает оплодотворяемость (В. В. Жаркий, 1983). Чтобы не допустить в процессе синхронизации ошибок при осеменении телок, следует регистрировать каждое обработанное животное, его индивидуальный номер, а место инъекции простагландина хорошо выстричь. Каких-либо отрицательных последствий применения простагландинов на последующую воспроизводительную способность телок (коров) не обнаружено.
Для контроля репродуктивной функции у овец широко используются препараты прогестерона как в отдельности, так и в комплексе с другими гормонами. Применяют их для стимуляции половой функции у анэстральных животных в неслучной период или для синхронизации охоты у проявляющих половую цикличность животных. Обычно прогестины вводят овцам в виде внутривлагалищных пессариев.
Наиболее эффективными при внутривлагалищном применении являются кронолон (флуорогестона ацетат), МАП (медроксипрогестерона ацетат) и ацетат мегестрола. Пессарии (губки) пропитывают спиртовым раствором препаратов из расчета: кронолона - 25-30 мг для овцематок и 40 мг для ярочек, ацетата мегестрола – 30 мг. Высушенные пессарии вводят с помощью влагалищного зеркала во влагалище на 12-14 дней. Затем извлекают за прикрепленную к ним нить. Охота наступает через 1-3 суток (чаще через 48-72 ч). Оплодотворяемость в синхронизированную охоту низкая, поэтому рекомендуется ее пропускать и осеменение проводить в следующую, естественную охоту. Снижение оплодотворяемости может быть связано со слабой абсорбцией гормона из пессариев или же вызвано нарушением баланса стероидных гормонов, отрицательно влияющим на передвижение и выживаемость спермиев. Кроме того, у 8-10% маток после извлечения губок отмечаются воспалительные процессы в половых путях.
Для повышения эффекта синхронизации в момент извлечеия пессариев или за 24-48 ч до этого вводят гонадотропин СЖК в дозах 1000 ме осенью и 1500 ме в июле-августе. Несколько лучшие результаты получают при инъекции гонадотропина за 48 ч до извлечения, однако это преимущество не компенсирует дополнительных затрат труда в процессе обработки.
В период полового сезона для синхронизации охоты у овец можно использовать ПГ-Ф2а или его синтетический аналог. Энзапрост вводят в дозе 10-12 мг внутримышечно дважды с промежутком в 8-9 дней. Охота проявляется через 36-48 ч после инъекции. Синхронизация охоты позволяет наиболее эффективно использовать искусственное осеменение. Однако нередко оплодотворяемость в синхронизированную охоту ниже, чем в естественную, а эмбриональные потери, наоборот, выше. Имеются сообщения (Н. Joe Bearden, John W. Fuquay, 1992), что и при естественном осеменении овец в синхронизированную простагландином охоту оплодотворяемость низкая, так как спермин повреждаются в шейке матки и немногие из них достигают яйцепроводов.
Для стимуляции охоты у овец в весенне-летний период можно использовать световой метод. Он заключается в искусственном сокращении продолжительности светового дня до 10-11 ч в сутки путем содержания животных в затемненных кошарах утром или днем. Эффект тем больший, чем ближе к естественному половому сезону создается затемнение. Однако само это мероприятие не всегда выполнимо, да и оплодотворяемость у овец со стимулированной охотой относительно низкая. Более высокие результаты стимуляции половой цикличности могут быть достигнуты при содержании овец вначале в сильно освещенном помещении со световой экспозицией до 21 ч в декабре-феврале с последующим уменьшением освещения до 14 ч в марте и 5 ч - в июне.
Световой фактор регулирует проявление половой Цикличности у овец посредством гормона эпифиза – мелатонина. Введение этого гормона животным внутривлагалищно, в рубец (2 или 4 таблетки по 18 мг через 20 или 10 дней в течение 40 дней), подкожно в виде трансплантанта или внутрь (дозы от 0,33 до 10 мг в день) симулирует уменьшение светового дня и обеспечивает начало полового сезона.
Вызвать половую цикличность у овец в весенне-летнее время с помощью ГСЖК труднее. Дело в том, что яичники чувствительны к гонадотропинам в предовуляционной фазе. Введение их в это время усиливает действие эндогенных гормонов и вызывает множественную овуляцию. При введении гонадотропинов в период охоты или вскоре после нее яичники реагируют в основном на ЛГ, но не на ФСГ и отвечают развитием гипертрофированных фолликулов и образованием кист. После сформирования желтого тела яичники становятся нечувствительными к гонадотропинам, и вызвать овуляцию в этот период удается редко. Если же она и происходит, то в большинстве случаев не сопровождается половой охотой. Двукратная с промежутком 16 дней инъекция ГСЖК также не позволяет добиться высоких результатов, хотя при этом несколько увеличивается число овец, пришедших в охоту. Использование комбинированного препарата овогена (состоящего из ГСЖК и витаминов A, D и Е) в неслучной сезон дает более стабильные результаты. После инъекции 2 мл препарата в течение 6 дней проявляет охоту до 60% овцематок.
В осенне-зимний период ГСЖК применяют овцам для вызова множественной овуляции и повышения многоплодия. Вводят нативную сыворотку подкожно в дозе 1200 ме на 13-14-й день цикла, т. е. за 2-3 дня до предполагаемой охоты. Введение гонадотропина в другое время снижает эффективность обработки. Для получения хороших результатов требуется ежедневная выборка овец в охоте и формирование из них групп для последующей обработки.
В свиноводстве в основном применяется способ синхронизации полового цикла у молодых животных. До начала обработки отбирают свинок в возрасте более 245 дней, хорошо развитых и проявивших естественную половую охоту не менее двух раз. Для обработки обычно используют суисинхрон или эвертас. Скармливают препарат в течение 20 дней из расчета 5 г на животное в сутки. Охота наступает через 4—5 дней после последней дачи препарата. С целью синхронизации овуляции и осеменения свинок в фиксированное время необходимо спустя 24 ч после прекращения скармливания препарата ввести ГСЖК в дозе 2500 ме, а через 84-88 ч после этого инъецировать хорионический гонадотропин в дозе 500 ИЕ. Осеменение проводят первый раз через 23-24 ч, второй - через 8-14 ч после первого. Взрослым свиноматкам через 24 ч после отъема поросят вводят ГСЖК в дозе 3000 ме, затем через 56-60 ч ХГТ. Осеменяют через 24 ч после инъекции ХГТ, а через 8-16 ч – повторно.
Оллитренболон или альтреногест также вводят с кормом или водой ежедневно в течение 18 дней в дозе 15-20 мг. Растворяют их в чистом растительном масле из расчета одна доза в 5 мл раствора. Осеменяют свинок в фиксированное время (на 6- и 7-й день) после прекращения дачи препарата. Процент опороса 64-73, численность приплода 9,5-9,8 на опорос. Взрослым свиноматкам эти препараты можно скармливать в дозе 20 мг в течение трех дней перед отъемом поросят. Эффективен комбинированный препарат ПГ-600 (ГСЖК-40 ИБ, ХГТ-200 ИЕ). Вводят его в день отъема или на 8-й день, если отсутствует естественная охота.
У свиней простагландины не проявляют лютеолитического действия до 11-12-го дня цикла, поэтому трудно определить режим инъекций их для синхронизации охоты. Однако возможно увеличить продолжительность функционирования желтого тела путем инъекции эстрогенов на 10-14-й день и затем ввести простагландин через 5-20 дней. Охота наступает на 4-6-й день.
В коневодстве нередко возникает необходимость предотвратить половую охоту у кобыл в неподходящее время или синхронизировать наступление ее у группы животных, прекратить слишком продолжительную охоту или подавить ненормальное проявление ее или же стимулировать начало половой цикличности.
Для задержки или синхронизации охоты можно инъецировать ежедневно прогестерон в дозе 0,3 мг/кг живой массы; охота наступает через 3-7 дней после прекращения введения гормона. Оллитренболон и аль-треногест также можно использовать для этих целей. Для начала половой цикличности препарат в прозрачном растительном масле вводят с кормом в течение 10 дней в дозе 0,044 мг/кг массы. Лучшие результаты достигаются, когда в яичниках присутствуют фолликулы, а освещенность помещения повышается. Для задержки охоты с целью регулирования нагрузки на жеребца эти гестагены вводят в течение 15 дней. Охота наступает через 2-3 дня.
Для регулирования полового цикла у кобыл можно использовать и ПГ-Ф2а или его аналоги. У кобыл желтое тело чувствительно к простагландину в период с 5 по 14-й день цикла. После инъекции простагландина охота наступает через 3 дня, овуляция – через 7-12 дней. Для ускорения овуляции можно инъецировать ГнРГ или хорионический гонадотропин.
Рациональное применение гормональных препаратов и других биологически активных веществ в большинстве случаев оказывается успешным. В то же время применение их без учета состояния организма может вызвать отрицательные последствия. Так, неграмотное применение половых гормонов и ГСЖК для стимуляции половой функции нередко является причиной развития кист яичников. В ответ на применение гонадотропинов организм продуцирует антигормональные антитела, которые могут вызвать расстройства воспроизведения. Ошибочная инъекция простагландина вызовет прерывание беременности. Все это необходимо учитывать в практической деятельности.
АНАТОМИЯ И ТОПОГРАФИЯ ПОЛОВЫХ ОРГАНОВ САМЦОВ
Половые органы самцов состоят из двух половых желез - семенников с придатками и спермиопроводов; семенникового мешка (мошонки), в котором находятся семенники; полового члена с наружным половым протоком – мочеполовым каналом (уретрой); придаточных половых желез: пузырьковидных, предстательной и куперовых (луковичных).
Мошонка (scrotum) представляет собой мешок, состоящий из двух полостей; расположена в паховой области между бедрами позади рудиментарных сосков. Снаружи мошонка разделена в медиальной плоскости вертикальной бороздой на две половины. У быков, баранов и козлов она несколько сплюснута спереди назад и имеет отчетливо выраженную шейку. У жеребцов мошонка занимает почти горизонтальное положение, а шейка выражена слабо. У хряка мошонка находится позади бедер, имеет косое направление, шейка отсутствует. В каждой полости мошонки располагается семенник, к которому основанием прикреплен семенной канатик.
Стенка мошонки состоит из трех слоев: кожи, мускульно-эластической оболочки и общей влагалищной оболочки (рис. 19). Кожа покрыта нежным волосом и содержит потовые и сальные железы. Она тесно связана с мускульно-эластической оболочкой. Последняя, заворачиваясь внутрь, делит семенной мешок на две половины; в верхней части разделяется на два листка, между которыми располагается половой член. На дне мошонки мускульно-эластическая оболочка срастается с общей влагалищной оболочкой, которая является продолжением брюшины. Общая влагалищная оболочка образует узкие паховые каналы, затем расширяется и выстилает обе полости мошонки. Она выделяет смазывающую жидкость. К внешней стенке оболочки с задней и наружной стороны прикрепляется наружный подниматель семенника. Этот поперечно-полосатый мускул берет начало от внутреннего отверстия пахового канала; он подтягивает семенники.
М ошонка
с ее мышцами поддерживает семенники,
защищает их от внешних воздействий и
обеспечивает постоянную температуру,
обычно ниже температуры тела, что важно
для активного сперматогенеза. Относительно
постоянная температура семенников у
нормальных самцов поддерживается
благодаря терморегулирующему действию
наружного поднимателя семенника. В
холодную погоду этот мускул сокращается,
подтягивает мошонку ближе к брюшной
стенке. Мошонка сморщивается, уменьшается
испарение влаги с ее поверхности и
отдача тепла во внешнюю среду. Внутренняя
семенная артерия с окружающим ее
сплетением из ветвей семенной вены
укорачивается и расширяется, что
способствует притоку к семенникам более
теплой крови. В жаркую погоду наружный
подниматель семенника расслабляется
и мошонка с семенниками далеко отвисает
от тела. При этом увеличивается испарение
влаги и отдача тепла во внешнюю среду,
удлиняется и сужается семенная артерия
с венозным сплетением, что способствует
охлаждению притекаемой к семенникам
крови. У хряков отвисание мошонки слабое.
Терморегулирующее действие мошонки
контролируется андрогенами и осуще-
ошонка
с ее мышцами поддерживает семенники,
защищает их от внешних воздействий и
обеспечивает постоянную температуру,
обычно ниже температуры тела, что важно
для активного сперматогенеза. Относительно
постоянная температура семенников у
нормальных самцов поддерживается
благодаря терморегулирующему действию
наружного поднимателя семенника. В
холодную погоду этот мускул сокращается,
подтягивает мошонку ближе к брюшной
стенке. Мошонка сморщивается, уменьшается
испарение влаги с ее поверхности и
отдача тепла во внешнюю среду. Внутренняя
семенная артерия с окружающим ее
сплетением из ветвей семенной вены
укорачивается и расширяется, что
способствует притоку к семенникам более
теплой крови. В жаркую погоду наружный
подниматель семенника расслабляется
и мошонка с семенниками далеко отвисает
от тела. При этом увеличивается испарение
влаги и отдача тепла во внешнюю среду,
удлиняется и сужается семенная артерия
с венозным сплетением, что способствует
охлаждению притекаемой к семенникам
крови. У хряков отвисание мошонки слабое.
Терморегулирующее действие мошонки
контролируется андрогенами и осуще-

ствляется после полового созревания. Если же окружающая температура настолько высока, что необходимый перепад температур тела и семенников не может сохраниться, то нарушается сперматогенез вследствие дистрофии спермообразующего эпителия. И чем длительнее воздействие температуры, тем больше степень поражения. Искусственным нагреванием мошонки в воде или посредством теплоизоляции ее вызывали полное бесплодие у баранов, быков, морских свинок. У баранов временное бесплодие может быть вызвано при содержании их зимой в теплом помещении.
У быков, которые имеют слишком крупные семенники и маленькую мошонку, затрудняется терморегуляция, нарушается сперматогенез, отмечается бесплодие.
Обычно температура семенников является средней между температурой тела и поверхности мошонки. У быков в зависимости от температуры окружающей среды температура на поверхности нижней части мошонки на 3-7ºС, а семенников – на 1,5-4ºС ниже температуры тела. У козла и барана разница между температурой тела и семенников более значительная и достигает 5-7ºС. Чем выше температура окружающей среды, тем менше перепад температуры тела исеменников, и наоборот.
Семенники (testis, orchis, didymis) вырабатывают мужские половые клетки и половые гормоны, главным образом тестерон. Каждый семенник представляет собой овальное, вытянутое тело, подвешен в мошонке на семенном канатике. У жвачных продольная ось семенника расположена вертикально, хвост придатка находится вверху сзади. У хряка расположение семенников косое, вблизи анального отверстия, хвост придатка находится спереди, вверху (рис. 20). У быка длина семенника варьирует от 10 до 13 см, ширина 5-6,5 см, масса – 300-400 г; такой же величины семенники у хряка, а у барана и жеребца они меньше – соответственно 9-11х6 см, масса – 200-300 г и 10-12х5-6 см, масса – 200-250 г.
С наружи
семенник и придаток семенника покрыты
собственно влагалищной оболочкой,
которая является висцеральным листком
брюшины. Она тесно сращена с белочной
оболочкой. Белочная оболочка заходит
внутрь семенника со стороны головки
придатка и образует средостение,
разделяющее семенник. От средостения
отходят соединительнотканные перегородки,
которые делят семенник на дольки (рис.
21). Перегородки соединяются с поверхностной
частью белочной оболочки.
наружи
семенник и придаток семенника покрыты
собственно влагалищной оболочкой,
которая является висцеральным листком
брюшины. Она тесно сращена с белочной
оболочкой. Белочная оболочка заходит
внутрь семенника со стороны головки
придатка и образует средостение,
разделяющее семенник. От средостения
отходят соединительнотканные перегородки,
которые делят семенник на дольки (рис.
21). Перегородки соединяются с поверхностной
частью белочной оболочки.
Семенник хорошо снабжен кровеносными сосулами и нервами, которые подходят к нему в составе семенного канатика. Иннервация осуществляется внутренним семенным нервом, который отходит от заднего брыжеечного узла. Кровоснабжение обеспечивается семенной артерией. Этот сосуд в месте соприкосновения с верхним краем семенника немного расширяется, идет дальше по заднему краю и разветвляется под оболочками, образуя систему извилистых артерий, хорошо просматриваемых снаружи. Концевые артерии разветвляются, входят в семенник вдоль средостения и питают ткань семенника.
Паренхима семенника представлена многочисленными дольками. В каждой дольке имеется один или несколько извитых (семенных) канальцев (seminiferous tubule). Промежутки между канальцами заполнены небольшим количеством соединительной ткани, в которой имеются интерстициальные клетки Лейдига, продуцирующие андрогены, а также кровеносные сосуды и нервы.
Образование спермиев происходит в семенных канальцах. Диаметр канальцев 100-200 мкм, длина – более 1 м. Общая протяженность их у зрелого быка достигает 5 км, у хряка - 4-6 км, у кобеля – до 1,2 км. Снаружи канальцы покрыты тонкой базальной мембраной. На ней располагается зародышевый эпителий. Эпителий представлен двумя типами клеток: базальными клетками Сертоли и первичными половыми клетками – сперматогониями. Клетки Сертоли расположены радиально на равных расстояниях друг от друга. Ядра их находятся у основания клеток, вблизи мембраны, а цитоплазма тянется до просвета канальцев. Они играют важную роль в питании сформированных, но еще незрелых спермиев. Кроме того, эти клетки секретируют эстрогены, ингибин, ГнРГ, специфические протеины (андрогенсвязывающий и регулирующий секрецию ФСГ), а также жидкость канальцев. Промежутки между клетками Сертоли заполнены сперматогониями, сперматоцитами и сперматидами. Извитые канальцы направляются к средостению семенника и впадают в прямые канальцы, а последние образуют сеть семенника.
П ридаток
семенника (epididymis) представляет собой
трубку различной ширины, идущую по всей
длине семенника. Состоит из головки,
тела и хвоста (рис. 22). Головка придатка
образована семявыносящими канальцами
(uasa efferentia) и начальной частью канала
придатка. Всего семявыносящих канальцев
12-15 (возможно от 6 до 20); длина их 10-20 см.
Они отходят от сети семенника (rete testis)
в месте связи семенника с придатком и,
сильно извиваясь, образуют сосудистый
конус. Последний составляет большую
часть придатка и соединен с длинным
каналом придатка. В области головки
канал придатка зигзагообразно извивается.
Извилины окружены белочной оболочкой
и вместе с ней придают головке придатка
вид большой плоской трубки. В области
тела придатка амплитуда извилин канала
уменьшается, сужается оболочка и это
придает телу вид более узкой и прямой
трубки. По направлению к нижней части
семенника канал расширяется, размах
его извилин увеличивается и вместе с
окружающей оболочкой он образует
массивный хвост. Хвост придатка связан
с семенником и фиксирован в нижней части
мошонки направляющей связкой семенника.
ридаток
семенника (epididymis) представляет собой
трубку различной ширины, идущую по всей
длине семенника. Состоит из головки,
тела и хвоста (рис. 22). Головка придатка
образована семявыносящими канальцами
(uasa efferentia) и начальной частью канала
придатка. Всего семявыносящих канальцев
12-15 (возможно от 6 до 20); длина их 10-20 см.
Они отходят от сети семенника (rete testis)
в месте связи семенника с придатком и,
сильно извиваясь, образуют сосудистый
конус. Последний составляет большую
часть придатка и соединен с длинным
каналом придатка. В области головки
канал придатка зигзагообразно извивается.
Извилины окружены белочной оболочкой
и вместе с ней придают головке придатка
вид большой плоской трубки. В области
тела придатка амплитуда извилин канала
уменьшается, сужается оболочка и это
придает телу вид более узкой и прямой
трубки. По направлению к нижней части
семенника канал расширяется, размах
его извилин увеличивается и вместе с
окружающей оболочкой он образует
массивный хвост. Хвост придатка связан
с семенником и фиксирован в нижней части
мошонки направляющей связкой семенника.
Семявыносящие канальцы и начальная часть канала придатка выстланы изнутри цилиндрическим эпителием с ресничками, причем в канале придатка эпителий многослойный. Реснички эпителия колеблются по ходу выносящего протока. На остальной части канал придатка выстлан цилиндрическим эпителием с секреторными клетками. В основании клеток имеются внутриэпителиальные железы. Просвет канальца придатка достигает 1 мм, при этом диаметр его увеличивается от головки к хвосту. Длина канала составляет: у быка – около 35 м, у хряка – 64 м, у жеребца – 80 м.
Образовавшиеся в семеннике спермин продвигаются по каналу придатка, дозревают, покрываются липидной оболочкой и накапливаются в нем.
Спермиопровод (ductus deferens) без ясной границы отходит от придатка и тянется вдоль семенника вверх. Вначале он извилист, затем становится более прямым. В составе семенного канатика проходит через паховый канал, а при попадании в брюшную полость отделяется от артерий, вен и нервов и направляется в тазовую полость, где располагается с соответствующей стороны над шейкой мочевого пузыря. В этом месте он ампулообразно расширяется (ампула спермиопровода). Вместе с пузырьковидной железой спермиопровод открывается общим протоком на семенном холмике, расположенном вверху начальной части мочеполового канала. Ампулы спермиопроводов тесно связаны между собой складкой брюшины. У хряка и кобеля ампулы отсутствуют. Спермиопровод снаружи покрыт брюшиной, в толще своей имеет продольный и кольцевой слои мышц; изнутри он выстлан многослойным цилиндрическим мерцательным эпителием с наличием базальных клеток, обладающих выделительной функцией. В слизистой оболочке конечной части ампул спермиопроводов у жеребца, быка, барана и кобеля имеются трубчатые железы. Они выделяют фруктозу, лимонную кислоту и пигмент липохром, который придает сперме некоторых быков светло-желтый цвет. В ампулах спермиопроводов спермин скапливаются перед эякуляцией. У быков ампулы хорошо пальпируются при ректальном исследовании. Длина их достигает 10-15 см, толщина - около 10 мм.
Семенной канатик (spermatic cord) представляет собой сдавленный с боков конус, основание которого прикреплено к семеннику и придатку семенника, а вершина доходит до внутреннего пахового кольца. Снаружи покрыт серозной оболочкой, в которую заключены спермиопровод, внутренняя семенная артерия и одноименные вена и нерв, лимфатические сосуды и слабо развитый внутренний подниматель семенника.
М очеполовой
канал (urethra) является общим протоком для
секретов семенников, придаточных половых
желез и для выделения мочи. Начинается
от шейки мочевого пузыря, тянется вдоль
тазовой полости (тазовая часть) и в
области большой седалищной вырезки
поворачивается вниз вперед и по желобу
(пенисовая часть) направляется к головке
полового члена, где и заканчивается
мочеполовым отверстием. Изнутри уретра
выстлана в начальной части переходным
эпителием, на конечном участке – плоским
многослойным эпителием. В толще эпителия
имеется много мелких уретральных желез.
Средний слой уретры - сосудистый,
представлен густым сплетением сильно
извитых вен и соединительной тканью с
гладкими и эластическими волокнами,
которые формируют пещеристое тело
мочеполового канала. Располагается
оно, главным образом, снизу. Вены в нем
расширены, образуют каверны и при
наполнении их кровью обеспечивается
зияние канала. Снаружи уретра прикрыта
мочеполовым и луковично-пещеристым
мускулами, сокращения которых способствуют
выведению спермы и мочи.
очеполовой
канал (urethra) является общим протоком для
секретов семенников, придаточных половых
желез и для выделения мочи. Начинается
от шейки мочевого пузыря, тянется вдоль
тазовой полости (тазовая часть) и в
области большой седалищной вырезки
поворачивается вниз вперед и по желобу
(пенисовая часть) направляется к головке
полового члена, где и заканчивается
мочеполовым отверстием. Изнутри уретра
выстлана в начальной части переходным
эпителием, на конечном участке – плоским
многослойным эпителием. В толще эпителия
имеется много мелких уретральных желез.
Средний слой уретры - сосудистый,
представлен густым сплетением сильно
извитых вен и соединительной тканью с
гладкими и эластическими волокнами,
которые формируют пещеристое тело
мочеполового канала. Располагается
оно, главным образом, снизу. Вены в нем
расширены, образуют каверны и при
наполнении их кровью обеспечивается
зияние канала. Снаружи уретра прикрыта
мочеполовым и луковично-пещеристым
мускулами, сокращения которых способствуют
выведению спермы и мочи.
Придаточные половые железы – пузырьковидные, предстательная и куперовы – расположены по ходу тазовой части мочеполового канала (рис. 23). Секрет этих желез составляет жидкую часть спермы.
Пузырьковидные железы (glandulae vesiculares) у быка имеют грушевидную форму и бугристую поверхность, расположены по одной с обеих сторон возле ампул спермиопроводов и открываются общими со спермиопроводами отверстиями в начальную часть мочеполового канала. Длина желез составляет 10 см и более, толщина – 2,5 см. Они разделены на дольки; выделяют секрет, содержащий фруктозу и лимонную кислоту. У мелкого рогатого скота пузырьковидные железы также имеют бугристую поверхность. У барана длина их 5 см, ширина – 2,5 см и толщина – 1,3 см. Величина желез у козла несколько меньше. У жеребца пузырьковидные железы мешковидные, с ровной поверхностью и центральной полостью, в длину достигают 13-15 см. Самые крупные пузырьковидные железы у хряка: длина до 12 см, ширина – 7 см и толщина – 3 см; поверхность их гладкая. У кобеля этих желез нет.
Предстательная железа (gl.prostata) состоит из тела и рассеянной части. Расположена в месте соединения шейки мочевого пузыря с мочеполовым каналом. У быка тело железы состоит из двух слитых воедино частей, имеет вид узкой полоски длиной 4 см, лежащей поперек уретры. Рассеянная часть железы окружает мочеполовой канал сверху и снизу и открывается в него несколькими отверстиями. Эта часть железы прикрыта мочеполовым мускулом и труднее обнаруживается. У барана и козла имеется только рассеянная часть. У жеребца и кобеля тело предстательной железы наиболее хорошо развито, а рассеянная часть выражена слабо или совсем отсутствует. У хряка тело железы крупное, имеет бугристую поверхность; рассеянная часть хорошо выражена. Выделяет предстательная железа жидкий секрет, который богат минеральными веществами и антагглютинином.
Куперовы, железы (gl. bulbourethralis) расположены по одной с каждой стороны мочеполового канала. У быков они находятся на расстоянии 10-12 см сзади от предстательной железы и частично прикрыты луко-вично-пещеристым мускулом. Снаружи они окружены толстым слоем волокнистой ткани и имеют вид небольших эллипсоидных тел беловатого цвета; длина их 2-3 см, толщина – 1 см. Протоки открываются одним отверстием в мочеполовой канал. Вырабатывают вязкое слизеподобное вещество. У барана и козла куперовы железы в 2-2,5 раза меньше, у кобеля они отсутствуют. У хряка железы сильно развиты, имеют вид толстых (2-3 см) продолговатых (до 15 см) пластинок шириной 3-4 см. У жеребца они величиной с грецкий орех.
Половой член (penis) – орган совокупления. Имеет прикрепленную часть – корень, основную часть – тело и свободный конец – головку. Основу полового члена составляет пещеристое тело. Оно берет начало от седалищных бугров двумя ножками, которые вскоре сходятся и образуют тело пениса. Пещеристое тело представляет собой трубчатую систему несимметричных кровеносных сосудов, которые при половом возбуждении наполняются кровью при высоком давлении. Этому способствует задержка оттока крови по глубоко расположенным венам вследствие сдавливания их набухающей эректильной тканью. Снизу пещеристое тело имеет углубление (желобок). В нем располагается мочеполовой канал. Снаружи половой член покрыт соединительнотканной (белочной) оболочкой.
У быка пенис цилиндрической формы, имеет около 90 см в длину и 2,5-3 см в диаметре. По направлению к свободному концу он несколько сужается. Пещеристая ткань слабо развита, за исключением корня. В расслабленном состоянии пенис образует S-образный изгиб, который расположен непосредственно сзади мошонки. Выпрямляющий мускул вытягивает пенис вперед, сжимает вены полового члена, тем самым способствует эрекции. За счет выпрямления изгиба длина полового члена увеличивается. Втягивающие (отводящие) мускулы прикреплены к передней части S-образного изгиба и при сокращении придают пенису изогнутое положение. Тело пениса полностью скрыто в области промежности и двуслойной мускульно-элаетической оболочке; удерживается подвешивающей связкой пениса.
Головка пениса быка имеет длину более 7 см, приплюснута и снабжена острым концом. На головке различают: шейку, отросток мочеполового канала и чехол (колпачок). На шейке головки имеется шов, закрученный по ходу головки в левую сторону. Во время эякуляции шов натягивается и конечная часть полового члена заворачивается в сторону, описывая почти полный круг. Отросток мочеполового канала не доходит до конца полового члена. В головке пещеристой ткани немного, и во время эрекции увеличивается она не сильно, но становится более твердой. Однако и при расслаблении консистенция головки твердая. Покрыта головка пениса тонким многослойным плоским эпителием, образующим много впячиваний в соединительнотканный слой. Обильно снабжена чувствительными нервными окончаниями из симпатического и парасимпатического нервных волокон. Рецепторы, воспринимающие холод (тельца Краузе, гениталъные) и осязательные (Мейснеровы тельца), расположены поверхностно. Более глубоко в эпителии находятся чувствительные к давлению нервные окончания – Фатер-Пачиниевы тельца.
У хряка, барана и козла половой член, как и у быка, имеет S-образный изгиб. В выпрямленном состоянии пенис достигает длины у хряка 50-70 см, у барана и козла – 40-50 см. У этих животных головка пениса слабо выражена, заострена, причем у хряка она штопорообразно закручена. Мочеполовой отросток у козла и барана продолжается за пределы пениса на 3-4 см; у барана он изогнут, у козла - прямой. У кобеля в передней части пениса заложена кость длиной 8-10 см; головка утолщена, а в задней части ее имеется пещеристое образование (луковица), которое набухает во время эрекции. У жеребца пенис прямой и в состоянии напряжения достигает длины 90 см или более; головка хорошо развита и содержит много эректильной ткани, которая сильно набухает в процессе совокупительных движений и придает головке грибовидную форму. Отросток мочеполового канала находится в ямке головки.
Головка полового члена располагается в препуциальном мешке (praeputium). У быка препуций длиной около 38 см. Внутренняя оболочка препуция имеет два листка: париетальный, выстилающий полость препуция, и висцеральный, переходящий на половой член; покрыта она плоским многослойным эпителием, сильно складчатая, при выдвижении полового члена складки расправляются и прикрывают тело пениса. В толще своей слизистая оболочка имеет особые сальные железы, которые выделяют препуциальную смегму, способствующую скольжению пениса. Снаружи препуций покрыт кожей; заканчивается отверстием, расположенным позади пупка. Отверстие снабжено пучком длинных волос.
У хряка полость препуция разделена кольцевой складкой на широкую переднюю и узкую заднюю части. В верхней стенке передней части находится небольшое отверстие, которое ведет в дивертикул препуция. У жеребца препуций сложный, образует двойной кожный мешок. В нем различают наружный и внутренний препуции, состоящие, в свою очередь, из наружного и внутреннего листков.
Половой член снабжается кровью наружной семенной и внутренней срамной артерией, у жеребца – еще и кровью запирательной артерии. Наружная семенная артерия питает и мошонку. Иннервируется пенис ветвями срамного нерва и наружным семенным нервом. Концевые части их представлены в виде сложных ге-нитальных, мейсснеровых (осязательных) телец.
СПЕРМАТОГЕНЕЗ
Сперматогенез можно разделить на две четко различающиеся фазы: сперматоцитогенез и спермиогенез. Сперматоцитогенез охватывает период размножения сперматогоний, превращения их в сперматоциты и заканчивается образованием сперматид. В течение спермиогенеза сперматиды в результате ряда различных новообразований и дифференцировок изменяют свою форму и превращаются в окончательно сформированных спермиев.
СПЕРМАТОЦИТОГЕНЕЗУ неполовозрелых животных сперматогоний расположены между клетками Сертоли одним слоем вдоль базальной мембраны семенных канальцев. С наступлением полового созревании они начинают интенсивно размножаться, заполняют периферию канальцев и продвигаются в их просвет. У быка и барана различают три класса сперматогоний: А, промежуточный и В. С учетом морфологии и степени дифференциации в каждом классе выделяют один или несколько типов: А0, AI; A2, А3, промежуточный, В1 и В2. Сперматогоний класса А менее дифференцированы и формируют резерв стволовых клеток. Пополняется он благодаря ассиметричному делению наиболее ранних типов клеток А: при делении одна дочерняя клетка остается как несовершенная стволовая клетка, а другая совершенствуется путем последующего митоза и мейоза (рис. 24,а). Образующиеся клетки типа В2 увеличиваются, затем также делятся митотически и в результате этого образуются два первичных сперматоцита. Этим заканчивается фаза "размножения", на которую уходит 13-14 дней.
В се
сперматогоний контактируют с базальной
мембраной канальца, но после последнего
митотического деления цитоплазма клеток
Сертоли внедряется между ней и первичным
сперматоцитом ("приподнимает его").
При этом цитоплазма соседних сертолиевых
клеток соединяется между собой, образуя
своеобразный "семенниковый барьер".
Он препятствует непосредственному
контакту клеток, подвергающихся мейозу,
с кровью и тем самым не дает элементам
иммунной системы распознавать их как
чужеродных или влиять на них другим
веществам крови.
се
сперматогоний контактируют с базальной
мембраной канальца, но после последнего
митотического деления цитоплазма клеток
Сертоли внедряется между ней и первичным
сперматоцитом ("приподнимает его").
При этом цитоплазма соседних сертолиевых
клеток соединяется между собой, образуя
своеобразный "семенниковый барьер".
Он препятствует непосредственному
контакту клеток, подвергающихся мейозу,
с кровью и тем самым не дает элементам
иммунной системы распознавать их как
чужеродных или влиять на них другим
веществам крови.
После следующей фазы «роста», в течение котрой сперматоциты увеличиваются в размере, следуют деления мейоза: образуются вначале два вторичных сперматоцита, а затем из каждого из них – по две сперматоцита, а заием из каждого из них – по две сперматиды. В итоге из каждого первичного сперматоцита образуется 4 сперматиды (из активной сперматогонии – 64); число хромосом в сперматидах уменьшается вдвое. В течение мейоза клетки продвигаются глубже в просвет канальца и тесно соединяются с клетками Сертоли.
Сперматиды имеют округлую форму (рис. 24, б) и относительно крупное светлое ядро. В теле молодой сперматиды находится клеточный центр, состоящий из центриоли, аппарата Гольджи и митохондрий. Центриоль представляет собой цилиндрическое тельце; стенка его состоит из девяти парных микротрубочек, в середине имеется еще две трубочки. На одном изконцов центриоли находится образование, напоминающее колесо. Аппарат Гольджи представлен отдельными диктиосомами, имеющими форму прямых или изогнутых палочек и крупных зернышек. Митохондрии сначала имеют форму гранул, в последующем –форму палочек и нитей. В них хорошо окреашивается периферическаячасть и плохо – центральная. В сперматиде, кроме того, обнаруживается остаток веретена, небольшое скопление руброфильных зернышек и темно окрашивающиеся добавочные тельца.
СПЕРМИОГЕНЕЗ
Вначале в теле сперматиды происходит перемещение отдельных компонентов клеточного центра и других составных частей. Так, отмечается поворачивание дочерних хромосомных пластинок на 90° по отношению к оси веретена. Центриоль перемещается к заднему полюсу ядра; сюда же смещаются и митохондрии. Аппарат Гольджи, напротив, мигрирует к переднему полюсу ядра, но после образования акросомы отходит в заднюю часть клетки, а в последующем отделяется от нее вместе с невключенной в спермий частью цитоплазмы.
В сперматиде ядро располагается в передней части клетки, ближе к периферии канальца. Аппарат Гольд-жи после миграции в переднюю часть клетки приобретает вид округлого почковидного тела, состоящего из плоских мешочков и большого количества мелких вакуолей. Среди основной массы мелких вакуолей в центре появляется несколько более крупных. В одной из них возникает темное зернышко – проакросомальная гранула. Иногда их несколько, но они сливаются в одну крупную гранулу. Вакуоль увеличивается за счет мелких и передвигается к переднему полюсу ядра. Часть поверхности ядра, к которой прилегает пузырек, временно уплощается, а цитоплазма перемещается в заднюю часть сперматиды, после чего полость акросомального пузырька сужается, стенки его спадаются, и он принимает форму двуслойной шапочки. Края ее далеко заходят за передний полюс ядра, образуя капюшон (акросому). Ядро снова принимает округлые контуры, затем удлиняется с общим удлинением сперматиды и формирует большую часть головки спермия. Акросомальная гранула на переднем крае головки заключена между ее наружной и внутренней оболочками, соединенными друг с другом по нижнему краю акросомы. После отхода цитоплазмы назад цитоплазматическая мембрана приходит в тесное соприкосновение со стенками акросомы.
Оставшаяся масса аппарата Гольджи, не участвующая в образовании акросомы, передвигается в заднюю часть клетки. Сюда же перемещается ицентриоль. При этом она делится на две части, которые представляют собой два аргентофильных зернышка. Располагаются они одно за другим перпендикулярно к поверхности клетки. В связи с этим различают проксимальную и дисталъную центриоли. Обе они погружены в глубь цитоплазмы и прикреплены к заднему полюсу ядра. При этом две центральные фибриллы дистальной центриоли размещаются на уровне поверхности клетки у базальной пластинки (аксиального зерна), периферические трубочки несколько углублены в цитоплазму, образовав здесь базальное тельце (кинетосому). От дистальной центриоли отрастает осевая нить.
Митохондрии группируются кольцом вокруг центриолей непосредственно позади ядра сперматиды. Они удлиняются, соединяются концами друг с другом и образуют одну или несколько длинных нитей, которые спирально обвивают осевую нить, образуя среднюю часть - тело спермия. В цитоплазме удлиняющейся сперматиды временно обнаруживается тонкая широкая трубка - хвостовая манжетка. Идет она от заднего сегмента ядра и заключает в себя осевую нить и часть цитоплазмы. Цитоплазма вне хвостовой трубки вместе с аппаратом Гольджи позднее исчезает.
Постепенно сперматида вытягивается, осевая нить удлиняется и в итоге формирует тело и хвост спермия. В области тела осевую нить обвивает спиральная нить из митохондрий. Последующая часть осевой нити покрыта слоем из плотных фибрилл. Они имеют вид полукружных ребрышек, своими концами соприкасаются и сливаются с двумя узкими продольными колонками, расположенными по противоположным сторонам хвостика.
На шейке полностью сформированного спермия остаются зернистые тельца Гольджи. Они исчезают вместе с невключенной в клетку цитоплазмой. Но в этом месте появляется цитоплазматическая капелька. Она обнаруживается до тех пор, пока спермий связан с сертолиевыми клетками, и исчезает в период прохождения его по каналу придатка семенника.
Образование спермиев у самцов после достижения половой зрелости происходит непрерывно. Активность сперматогенеза волнообразно перемещается по семенным канальцам. Между двумя последовательно появляющимися группами однотипных клеток в одном участке канальца проходит: у хряка – 8,6 дня, у барана – 10,3, у жеребца – 12,2, у быка – 13,5 дня. Это семенной эпителиальный цикл. В нем иногда различают 8 стадий (Ortavant, 1977). У быка для полного формирования спермиев из сперматогоний необходимо около 60 дней. Для продвижения по каналу придатка семенника требуется еще 8-14 дней. Головку и тело придатка спермий проходят за 5 дней, а хвост придатка – за 5-9 дней, в зависимости от интенсивности полового использования производителя.
Передвижение спермиев по внутренним протокам осуществляется за счет: положительного давления в канальцах вследствие постоянного вхождения в их просвет новых клеток; отрицательного давления в семявы-носящих канальцах и канале придатка семенника, которое создается в результате эякуляций и всасывания жидкости, поступающей в большом количестве из семенника; движения ресничек, выстилающих сёмявы-носящие канальцы и начальную часть канала придатка и, наконец, давления на семенники, возникающего в процессе движения животного. Попав в канал придатка, спермин продвигаются дальше за счет перистальтических движений его стенок.
В извитых канальцах семенника впервые спермин могут быть обнаружены в возрасте около 5 мес. – у баранов и хряков, 7,5 мес. – у быков и 13 мес. – у жеребцов. Способность же к воспроизведению у них появляется несколько позднее и связана с началом образования спермы, содержащей достаточное количество нормальных спермиев, а также с проявлением полового влечения. Факторы внешней среды (температура, продолжительность светового дня, кормление и содержание, возможные болезни) оказывают большое влияние на сроки достижения половой зрелости у самцов.
Образование и продвижение спермиев по половым протокам находится под влиянием гормонов гипофиза и семенников. Определенную роль играют также гормоны надпочечников и щитовидной железы.
Еще до наступления половой зрелости семенники подвергаются воздействию ФСГ и ЛГ. Лютеинизирую-щий гормон (гормон, стимулирующий интерстициалъные клетки) воздействует на рост и функцию клеток Лейдига, повышает секрецию и выделение андрогенов. Действие осуществляется через аденилатциклазу, путем сокращения до минимума периодов стероидогенеза. У быков интенсивность секреции и выделения половых гормонов повышается с 4-6-месячного возраста. Вскоре начинается и выделение фруктозы придаточными половыми железами.
У половозрелых животных выделение ЛГ происходит нерегулярно, эпизодически, с промежутками в 2-4 ч (3-7 раз в течение дня). Характер выделения ФСГ расшифрован недостаточно полно.
Примерно через 40 мин после каждого пика ЛГ следует пик тестостерона, затем через 40-80 мин содержание андрогена снижается до исходного уровня. Андрогены повышают чувствительность сперматогенного эпителия семенных канальцев к ФСГ. Этот гормон, вызывая размножение сперматогоний, дает начало сперматогенезу.
ФСГ воздействует и на клетки Сертоли, стимулирует секрецию андрогенсвязывающего протеина (АСП) и превращение тестостерона в эстрогены. Секреция ФСГ регулируется половыми гормонами иингибином, а также регуляторным протеином, секретируемыми клетками Сертоли.
Андрогены воздействуют на весь половой аппарат самца, стимулируют рост и функционирование придаточных половых желез, тем самым влияя на состав спермы, обеспечивают оптимальные условия для сперматогенеза, продвижения и созревания спермиев в канале придатка семенника. После превращения в мозгу в эстрогены тестостерон регулирует выделение ЛГ и влияет на поведение самца.
В семенных канальцах тестостерон превращается в 5-дигидротестостерон, который более устойчив к метаболическим превращениям. Здесь оба этих метаболита связываются с андрогенсвязывающим протеином. АСП обеспечивает высокую концентрацию тестостерона в извитых канальцах и канале придатка семенника (в 100-300 раз выше, чем в периферической крови), что важно для нормального сперматогенеза. Абсорбируется протеин в канале придатка семенника. Высокая концентрация тестостерона поддерживается также в сети семенника, семявыносящих канальцах и проксимальной части придатка.
Для поддержания постоянного нормального сперматогенеза требуется совместное воздействие ФСГ и андрогенов, вырабатываемых под влиянием ЛГ. В связи с этим должно быть строго определенное соотношение между этими гормонами. Уменьшение секреции гонадотропных гормонов тормозит продукцию андрогенов и ослабляет сперматогенез. Понижение уровня андрогенов стимулирует дополнительное выделение гонадотропинов, которые снова повышают секрецию андрогенов и образование спермиев.
У самцов, используемых в течение года, постоянный уровень секреции гонадотропинов. У сезонно используемых животных секреция ФСГ и ЛГ снижается вскоре после окончания случного сезона и остается низкой до следующего сезона. С наступлением его внешние факторы (продолжительность светового дня, температура и др.) дают толчок к выделению гипоталамусом ГнРГ, который в свою очередь стимулирует секрецию и выделение ФСГ и ЛГ гипофизом. У баранов в регуляции сперматогенеза важную роль играет щитовидная железа. Введение им тироксина летом, когда тормозится образование спермиев и увеличивается число их с различными аномалиями, способствует уменьшению таких клеток и повышению образования нормальных спермиев.
Образовавшиеся в процессе сперматогенеза спермин продвигаются по внутренним половым протокам и скапливаются в канале придатка семенника, который представляет собой значительную емкость. У быка в каждом придатке содержится до 4 мл сливочно-белого цвета вязкой жидкости с концентрацией спермиев от 2,5 до 5 млрд./мл. Половые клетки занимают примерно половину объема этой жидкости. Всего оба придатка у быка могут вмещать до 20 и даже до 40 млрд. спермиев, у барана - до 60 млрд.
Секреторная функция придатка семенника и передвижение спермиев по половым протокам регулируются окситоцином.
Поступающие в канал придатка семенника спермин имеют в месте соединения головки и тела цито-плазматическую капельку. По мере прохождения их по каналу капелька продвигается по направлению к хвосту и затем сбрасывается. С цитоплазматической капелькой связывают приобретение спермиями липопротеидного покрова (оболочки).
В канале придатка спермин приобретают подвижность и оплодотворяющую способность. У большинства видов половые клетки, взятые из семенников самца, не способны к движению, хотя они живые и могут совершать слабые колебательные движения хвостиком. У быка спермин из проксимальной и средней части головки также неподвижные, но подвижность появлялась в участках придатка вблизи семенников и уже хорошо проявлялась у спермиев из хвоста придатка. У быков нормальная подвижность и биологическая полноценность спермиев в канале сохраняется в течение 30-35 дней, а отдельные спермин могут сохранять подвижность 2-3 мес. Спермин кроликов сохраняют жизнеспособность 38 дней.
СПЕРМА: СОСТАВ И СВОЙСТВА
Семенная жидкость (сперма) состоит из двух основных частей: спермиев и жидкой части - плазмы. Семенники вырабатывают спермиев, а также являются источником различных клеточных фрагментов и жидкости, не всосавшейся при прохождении по внутренним половым протокам. Одним из компонентов этой жидкости является тестостерон. Жидкая часть плазмы является продуктом деятельности придаточных половых желез, канала придатка семенника и ампул спермиопроводов.
ВНЕШНИЕ СВОЙСТВА СПЕРМЫ
Количество и внешний вид спермы, соотношение жидкой части и спермиев и многие другие свойства варьируют в зависимости от вида животных и их индивидуальных особенностей. Жвачные выделяют небольшие эякуляты с высокой концентрацией спермиев. Жеребец, хряк и кобель дают большие эякуляты, но с низким содержанием половых клеток. У птиц объем эякулятов небольшой, а концентрация спермиев очень высокая (табл. 1).