

§ 22. Механизмы кратковременной • памяти
Нейрофизиологические корреляты краткосрочной памяти. К данной категории или стадии относится память на только что минувшие события. Представим себе следующую экспериментальную ситуацию. Обезьяна обучается по условному сигналу — зажигание лампы справа или слева на пульте — выполнять движение нажатия на соответственно правую или левую педаль или рычаг. Если выбор педали или рычага выполнен правильно, такое действие вознаграждается пищей {А. С.Ба-туев и др., 1988). Но сигналы подаются в случайном порядке и после каждого сигнала наступает пауза, которая может длиться разное время — от 5 до 20 с. В течение этого периода, называемого периодом отсрочки, животное не имеет доступа к рычагам и педалям: они скрыты от него непрозрачным экраном. После отсрочки экран открывается и обезьяна может выполнить требуемое действие (рис. 59). Значит, животное, получив условный сигнал, должно удержать в памяти информацию о его местоположении в течение всего периода отсрочки, а затем использовать эту информацию для осуществления правильного движения.
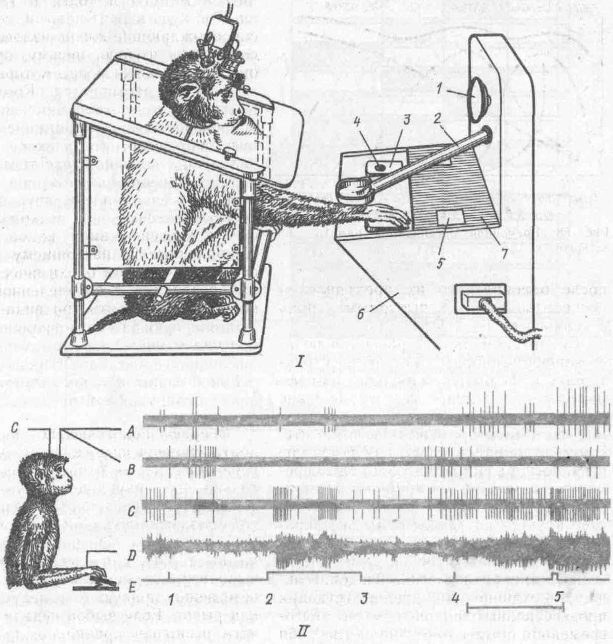
Рис. 59. Методика изучения нейронной активности мозга в тестах на краткосрочную память у обезьян;
/ — экспериментальная процедура; // -- экспериментальная программа и ее нейрональные корреляты; / — лампа-вспышка (предупреждающий сигнал); 2 — тубус для подачи пищи; 3 — педаль; 4 — лампа (условный сигнал); 5 — прозрачное окно в экране (7); 6 — общая панель; А и В активность нейронов лобной коры; С — двигательной коры; О — электромиограмма дельтовидной мышцы плеча, Е — отметки сигналов (/ — предупреждающий; 2 и Л — условный; 4 — пусковой (отодвигание экрана); 5 — нажатие на клавишу). Калибровка времени - 2 с
Такое отсроченное поведение является общепризнанным методическим подходом для изучения механизмов кратковременной памяти, с помощью которой сохраняется информация о только что прошедших событиях, но необходимых для выполнения предстоящего поведенческого акта. У таких животных из различных отделов мозга с помощью микроэлсктродоп длительно
регистрируется импульсная активность нескольких нейронов. Исследования показали, что информация о пространственном расположении условного сигнала кодируется в импульсной активности нейронов прежде всего лобной и теменной ассоциативных полей коры больших полушарий. Такое кодирование осуществляется либо рисунком разряда нейронов, либо частотой им-

Рис. 60. Сравнительное'распределение пространственно селективных нейронов лобной и теменной коры обезьян в тестах на кратковременную память:
цифры показывают процентное отношение к общему числу пространственно селективных нейронов. Этапы экспериментальной программы: / — предупреждающий сигнал; 2 — период неспецифического ожидания; 3— условный сигнал; 4 — отсрочка; 5 — пусковой период программы (открытие акра на, нажатие на рычаг, получение пищевого подкрепления)
пульсации нейронов, получивших название пространственно селективных. Оказалось, что эти нейроны не представляют собой однородную популяцию, а подразделяются на несколько групп (рис. 60). Часть нейронов обнаруживает свойство пространственной селекции только в период предъявления условного сигнала — такие нейроны названы сенсорными. Другая группа нейронов обнаружила различия в импульсной активности только п период отсрочки — их именуют нейронами памяти. Третья группа аналогичное свойство проявила только после открытия экрана, в момент, непосредственно предшествующий моторному акту, поэтому их назвали нейронами моторных программ. Описаны и смешанные типы нейронов.
Детальный анализ самого периода отсрочки показал, что все ее время заполняется кратковременными вспышками импульсной активности разных
нейронов лобной коры: в начале отсрочки, в середине и в конце, либо вспышек активности одних и тех же нейронов. И только незначительное число клеток поддерживает высокую импульсную активность на протяжении всего периода отсрочки (рис. 61). Пространственно селективные сенсорные нейроны стоят на входе сложных' нейронных кортикальных цепей и лишь кодируют сенсорные признаки сигнала, в данном случае его пространственное положение. Нейроны, активирующиеся в начальный период отсрочки, детектируют полученную информацию о свойствах условного сигнала, далее следует передача возбуждения по принципу эстафеты другим нейронным популяциям» каждая из которых отдает аксоны к нейронам-сумматорам, и от последних происходит передача задержанной таким образом информации на программирующие и пусковые структуры.
Именно для лобных отделов коры, где и обнаружены вышеописанные феномены, характерны замкнутые нейронные круги типа «ловушек», в которых импульсный поток может циркулировать (реверберировать) до тех пор, пока не будет переключен на эфферентные модули из крупных пирамид (рис. 62). Последнее может служить структурной основой удержания п ловушках импульсных потоков и эстафетного переключения активации от одного нейронного объединения к другому. Лоренте де Но был одним из первых морфологов, кто описал сложные замкнутые цепи нейронов в разных частях мозга. На основе морфологи ческих данных была создана модель памяти из замкнутых цепей, в которых импульсы циркулируют без подкрепления (рис. 63). И. С. Беритов (1948) показал, что часть звездчатых клеток проекционных зон коры имеет аксон, заканчивающийся в поле своих дендри-тов. Такая структура может работать как ловушка возбуждения.
Внутрикорковую реверберацию рассматривают как базовый механизм краткосрочной памяти для лобной коры. Важнейшим фактором формирования
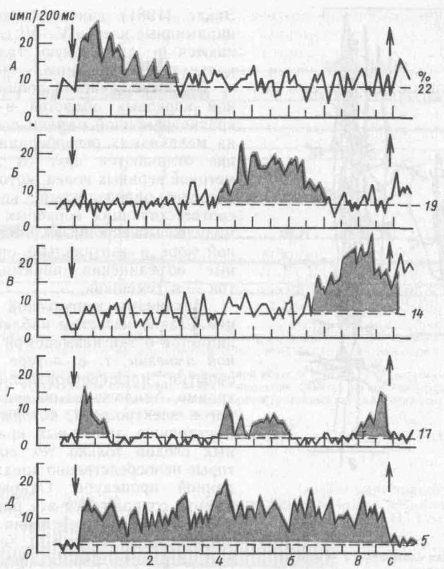
Рис. 61. Основные типы перестроек импульсной активности нейронов лобной коры в период отсрочки (по А. А. Пирогову)
стрелки указывают начало и окончание отсрочки; цифры — процентное соотношение нейронов; пунктир — уровень фоновой активности
кратковременной памяти является ориентировочный рефлекс, который фиксирует внимание животного для более полного восприятия новой информации. В опытах на обезьянах было установлено, что отвлечение внимания от экспериментальной ситуации при действии постороннего раздражения приводит к разрушению рисунка импульсной активности пространственно селективных нейронов лобной коры в период действия условного сигнала и отсрочки и к появлению ошибочных ответных реакций животного (рис. 64).
Иные явления характерны для теменной коры, где хотя и описано множество пространственно селективных нейронов, но в пределах време-
ни отсрочки не обнаружено феноменов волнообразной активности эстафетного типа, как в лобной коре. Более того, пространственная селективность сохраняется на протяжении всего периода отсрочки (рис. 65). Корковые нейронные объединения типа ловушек здесь не обнаружены, а преобладающим типом нейронов являются послойно расположенные эфферентные пирамиды. Пространственная селективность, определяющая рисунок нейронной активности теменной коры, обусловлена взаимосвязанной работой коры и таламичес-ких ядер в составе единой таламо-париетальной ассоциативной системы (см. гл. 7). Иными словами, сохранение в период отсрочки импульсной
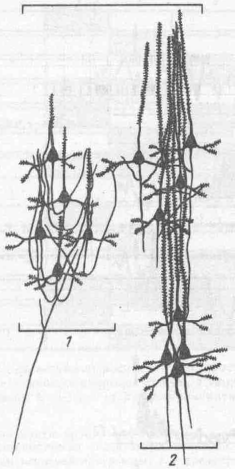
Рис. 62. Основные типы нейронных объединений в лобной коре у обезьян (по Г. П. Демьяненко,
1980):
/ — нейронная «ловушка»; 2 — вертикальный модуль
активности пространственно селективных нейронов теменной коры обусловлено импульсной реверберацией по длинным тал а мо-кортикальным кольцам прямой и обратной связи. Дж.
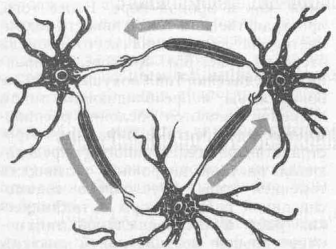
Рис. 63. Схема реверберирующей нейронной цепи; (по Г. Адаму, 1983)
Экклс (1981) считает, что корковые пирамидные клетки V—VI слоев вовлекаются в длительную таламо-кортикальную реверберацию.
Вероятно, участие лобной и теменной корковых областей в процессах кратковременной памяти основывается на механизмах реверберации. Последние отличаются друг от друга геометрией нервных колец, которые в свою очередь определяются конструкцией соответствующих корковых нейронных модулей — нейронные ловушки в лобной коре и вертикально организованные объединения пирамидных клеток — в теменной.
Механизмы импульсной реверберации. Давно известны наблюдения клиницистов о так называемой ретроградной амнезии, т. е. потере памяти на события, предшествовавшие мозговой травме. Аналогичны последствия судорог и электрошока, которые приводят к стиранию лабильных кратковременных следов только тех событий, которые непосредственно предшествовали данной процедуре. Одновременно в памяти сохраняются все ранее возникшие впечатления и сведения. Это послужило основанием для Д. Хебба (1949) рассматривать двойственную природу следов памяти. Он предположил существование клеточного ансамбля как закрытой системы, активность которой может реверберировать после прекращения сенсорного процесса. Ансамбли связаны друг с другом и могут активироваться эндогенно. Они также могут образовывать связи с моторными клеточными ансамблями. Д. Хебб считал, что фиксация следа связана со стойкими изменениями синаптичес-кой проводимости, возникающими при повторном поступлении к синапсу импульсного разряда определенной конфигурации. К аналогичным заключениям пришли и другие ученые (Д. Дже-рард, Ю. Конорски, И. С. Бериташви-ли). Дж. Экклс (1981) установил, что при частом функционировании синапса (его употреблении) возникает облегчение — длительные изменения синаптической проводимости, а при неупотреблении проводимость ухудшается.
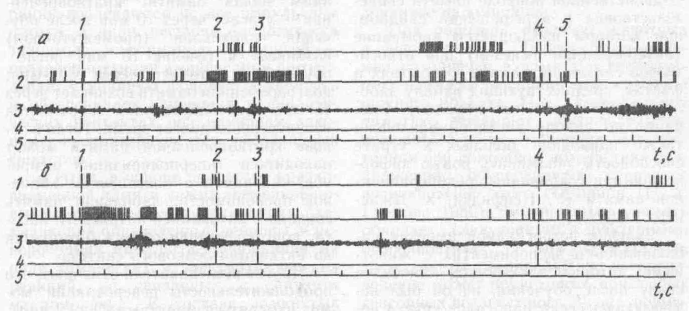
Рис. 64. Активность нейронов лобной, моторной коры и электромиограмми мри правильном решении задачи (а) и при отка.че от ее выполнения вследствие отвлечения внимания (6):
1,2 — записи активности нейронов лобной и моторной коры; 3 — ЭМГ; 4 — отметки стимулов; 5 — отметка времени (1 с); вертикальные штриховые линии — проекции отметок на осциллограммы; слева — направо: вспышка (предупреждающий сигнал}; зажигание лампы (условный сигнал); открытие экрана (пусковой сигнал); 5 — нажатие на клавишу (условная реакция)
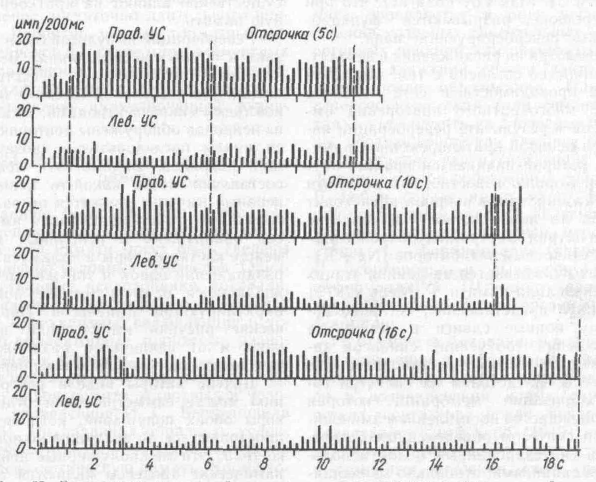
Рис. 65. Гистограммы импульсной активности пространственно селективного нейрона теменной коры при различных длительностях периода отсрочки
О двойственной природе памяти свидетельствовал и корсаковский синдром, при котором наблюдается забывание (антероградная амнезия) при относительно сохранной памяти о событиях и фактах, предшествующих началу заболевания (алкогольный паралич). Операция но поводу височной эпилепсии также приводила больных к утрате способности запоминать новую информацию при сохранении предоперационной памяти (У. Пенфильд, X. Джаспер, 1956).
Явления ретроградной амнезии наблюдались в экспериментах с животными, которым электроток наносился сразу после обучения, но он был неэффективен, если наносился спустя некоторое время после процедуры обучения. Многочисленные эксперименты привели к выводу о том, что все воздействия, так или иначе отражающиеся на проводимости в синаптических меж-нейрониых контактах, приводят к нарушениям процессов кратковременной памяти. Д. Мак Гоу полагает, что при электрошоке разрываются функциональные реверберирующие цепи.
Несмотря на расхождения в деталях, большинство согласно с тем, что изменение проводимости в сети синапсов после многократного повторения импульсов в результате реверберации является основой кратковременной памяти, с которой начинается процесс обучения. Хорошо известно, что во время функционирования нейрона происходит потеря им ионов калия и накопление ионов натрия. Внутрижелудочковое введение животным ингибиторов (Na + K) -АТФазы сказывается на ранних этапах формирования памяти. Р. Мерк (1979) выдвинул представление, согласно которому ионные сдвиги в результате упражнения (обучения) синапсов запускают длительные восстановительные процессы. Следствием их является ги-перполяризацня мембраны, которая сопровождается поступлением аминокислот и синтезом определенных белков. Вновь синтезированный белок используется синапсами, стабильно меняющими свою эффективность. При этом указывается на следующие временные гра-
ницы видов памяти: кратковременная — угасает через 10 мин после обучения, лабильная (промежуточная) возникает в течение 15 мин после обучения и угасает через 30 мин; долговременная память возникает через 45 мин после обучения и сохраняется неопределенно долгое время. Тогда в основе кратковременной памяти может находиться гиперполяризация нейронов, связанная с повышением калиевой проводимости; лабильная память связана с активацией натриевого насоса; долговременная память базируется на активации белкового синтеза.
Другие исследователи отмечают, что продолжительность реверберации может измеряться минутами (2—12 мин). Подчеркивается большое значение медиатора ацетилхолина, ибо воздействие на системы, выделяющие и связывающие ацетилхолин (путем введения атропина и скополамина, нарушающих его рецепцию постсинаптической мембраной или ядов ацетилхолинэстеразы), существенно влияют на кратковременную память.
Реверберация импульсов была показана в прямых исследованиях Всрцеа-но при одновременной регистрации активности нескольких соседних нейронов тремя микроэлектродами. В каждом из нейронов обнаружены повторяющиеся циклы последовательно возникающих разрядов. Видимо, эти нейроны составляют часть какой-то цепи, где нервный импульс движется по замкнутому пути. Такая циркуляция импульсов обнаружена в нейронных цепях между клетками коры и таламуса. При раздражении одной и той же частотой наблюдался один и тот же порядок циркулирующих импульсов. Периодические рисунки импульсации возникают и от единичного раздражения, что доказывает наличие реверберации.
Другие авторы видели реверберацию между симметричными отделами коры обоих полушарий, которая распространялась по волокнам мозолистого тела. Эти межполушарные нейроди-намические процессы являются одним из составных звеньев системной организации кратковременной памяти.
С одной стороны, в цикл реверберации вовлекаются не все импульсные потоки, а лишь несущие биологически значимую информацию, с другой — реверберирующая импульсация преодолевает случайную фоновую активность и обеспечивает изменения в синапти-ческих контактах под влиянием закономерно повторяющихся неслучайных факторов.
ф Итак, в основе процессов кратковременной памяти лежит временное повышение проводимости в синапсах, связывающих определенные нейроны, и реверберация импульсов, основанная на ряде химических и электрохимических реакций, не связанных с синтезом макромолекул. В составе замкнутых нейронных цепей в коре мозга участвуют тормозные интерпейроны. В самоорганизации следовых процессов в пределах нервного кольца могут играть значительную роль посттормозные реакции при наличии в цепи возвратного торможения. Это создает наиболее оптимальные условиях хранения следов в течение достаточно длительного времени. Вероятно, участие тормозных интернейронов в составе многоэлементных внутрикорковых цепей способствует более длительному хранению следов и поддержанию циркулирующей активности.