

§ 19. Нейронная организация условно-рефлекторного процесса
Данные о перестройке нейрональ-ной активности при выработке условных рефлексов у бодрствующих животных представляют существенное значение для понимания функциональной организации целостной системы обучения. Такие изменения реакций нейронов могут проявляться практически во всех исследованных структурах мозга. Логично допустить поэтому, что возможность образования ассоциаций является базовым свойством нервной системы.
В связи с тем, что сигнальная сущность условного рефлекса состоит
Таблица 5. Классификация разных видов функциональных перестроек активности нейронов головного мозга при формировании и реализации временной связи (Б.П.Котляр, 1980)
|
Ассоциативные перестройки
|
Нсассоциатнвные пе встройки
| ||
|
фи ического типа
|
тонического типа
|
физического типа
|
тоническото типа
|
|
Перестройки, отражающие уровень центрального тонуса, характерного для реализации временной связи
|
Перестройки, характерные для тонической составляющей ассоциативного процесса
|
Переотройки, отражающие динамику ориентировочной реакции
|
Перестройки, отражающие динамику ориентировочной реакции
|
|
Специфические перестройки, воспроизводящие безусловный ответ
|
|
Перестройки, связанные с доминирующей мотивацией
|
Перестройки, связанные с доминирую-щей мотивацией
|
|
Перестройки, отражающие реакции на время в рамках действия ассоциируемых раздражителей
|
|
|
|
|
Перестройки, связанные с условной ориентировочной реакцией
|
|
|
|
во временном сочетании, т. е. в ассоциации раздражений, представлялось необходимым, в первую очередь, выделить перестройки нейрональной активности, связанные с ассоциацией, в отличие от всех остальных сопровождающих изменений в активности нейронов.
Р, Томпсон рекомендует для этой цели использовать три критерия: 1) специфичность изменений активности на условный сигнал в сравнении е изменениями, вызываемыми индифферентными раздражителями; 2) параллельность развития изменений нейронной активности и компонентов поведенческого условного ответа и 3) градуальный рост выраженности пластических изменений на нейронном уровне в процессе выработки поведенческого навыка.
Другие авторы (Э. Кен дел, 1980) предложили для той же цели рассматривать два независимых критерия: 1 — природа ответа на условный сигнал, т. е. является ли условный ответ новым (классический условный ответ) или модификацией условной реакции на сигнальный раздражитель (альфа-условный ответ), и 2— специфичность реакции для последовательного сочетания раздражителей. Иными словами, не может ли аналогичный ответ развиваться при случайном предъявлении тех же раздражителей (при процедуре псевдообусловливания) или определяться сен-ситизацией.
Б. И. Котляром (1986) предложена классификация перестроек активности нейронов в процессе условно-рефлекторной деятельности (табл. 5). Принципиальным является введение в состав условно-рефлекторного процесса тонического типа активности нейронов. Основанием этому служили специальные исследования с переключением разнородных условных рефлексов, которые позволили рассматривать центральный мозговой тонус в качестве непременного компонента процессов обучения и памяти.
По результатам обучения на уровне отдельной клетки можно судить, что наиболее существенные изменения характерны для корковых полисенсорных нейронов (М. Я. Рабинович, 1975), функциональные свойства которых по мере сочетаний претерпевают эволюцию, отражающую становление его афферентной специализации. Так, до первых сочетаний стимула с подкрепляющим рефлексом нейроны проявляют преимущественно моносенсорныс реакции, т. е. большинство из них отвечает на стимулы адекватной сенсорной модальности стабильными коротко-латентными разрядами. По мере последующих сочетаний эти же нейроны приобретают способность отвечать на стимулы различных сенсорных модальностей и параметров; их активность становится полисенсорной. Этот период
соответствует фазе генерализации условного рефлекса, когда положительное действие обнаруживают не только сигнальный, но и другие сенсорные раздражители. Вероятно, именно в этот период данная рефлекторная система начинает приобретать доминантные свойства, выделяться среди прочих и подкрепляться разнообразными воздействиями. Наконец, при достаточно упроченном условном рефлексе ответы нейронов снова приобретают моносенсорный характер, но теперь уже. это специальные моносенсорные реакции на стимул определенной модальности и физических параметров, т. е. избирательная реакция только на тот стимул, который приобрел сигнальное значение для поведенческого акта.
Следует отметить, что характеристики процесса привыкания реакций нейронов при монотонных предъявлениях сенсорных стимулов не коррелируют с процессом ассоциативного обучения. Одни нейроны в популяции могут проявлять феномен привыкания, а другие — сенситизацию, следовательно, нейрофизиологические механизмы привыкания и ассоциативного обучения могут быть различны по своей организации. Если в основе привыкания лежит уменьшение числа квантов медиатора, ведущее к снижению амплитуды постси-наптичсских потенциалов, ассоциативное обучение может быть связано со сдвигом эндогенной активности обучаемого нейрона. Однако нельзя исключить возможность причинно-следственной связи между привыканием и сенситиза-цией. Действительно, на одном и том же мультисенсорном нейроне можно наблюдать в процессе предъявления стимулов оба явления сразу. Например, в начале процедуры привыкания ответа на световой стимул, ^лектрокожный стимул, подаваемый в интервале функциональных сдвигов после светового стимула, оказался неэффективным. Однако по мере ослабления ответной реакции нейрона на световой стимул (привыкание) начинает появляться ответ того же нейрона на электрокожный стимул (сенситизация). Последний достигает своего максимума при отсутствии
ответа нейрона на световую вспышку (А. С. Батуев, 1978).
Преобладание того или иного механизма существенно зависит от сигнальной значимости предъявляемого разд^ ражения. Так, в опытах на бодрствующих обезьянах было установлено, что сенсибилизация в реагировании нейронов лобной коры возникает лишь в том случае, если раздражители прямо или косвенно связаны с мотивационной сферой животного, сигнализируют об опасности или несут на себе элементы новизны. Привыкание ответов тех же нейронов наблюдалось в тех случаях, когда используемые раздражители были индифферентными или хорошо знакомыми для животного. Однако если ранее индифферентный агент приобретал качества сигнального фактора, будучи сочетай во времени. с биологически значимым болевым агентом, то картина резко менялась (рис. 48). Ранее угашенные нейронные ответы уже через 9—10 сочетаний возобновляются, что характеризуется всеми чертами выработки условного рефлекса и подтверждается параллельной регистрацией ЭМГ рабочей конечности. Такой пластический характер мультисенсорной конвергенции к нейронам лобной коры, диапазон и устойчивость которой определяется сигнальной значимостью раздражителя, в конечном итоге формируется доминирующей мотивацией и системой выработанных ранее временных связей.
По критерию наличия или отсутствия изменений реакций нейронов нельзя считать, что обучение характерно лишь для определенных отделов или уровней мозга. Имея в виду неравноценность отделов мозга для процессов обучения, следует признать, что сами по себе изменения реакций нейронов при выработке временных связей в отсутствие дополнительных контрольных процедур еще не является доказательством причастности тех или иных структур мозга к процессам обучения.
Обычно ищут корреляцию между изменением реакций нейронов и поведенческими сдвигами. Однако для заключения об организации системы, обеспе-

Рис. 48. Динамика импульсной активности коркового нейрона лобной области у обезьян и электромиограмма (ЭМГ) мышц плеча
при выработке условного рефлекса (по А.С. Батуеву и др., 1981)
I —- (сверху вниз): нейронная активность, ЭМГ и график изменения средней частоты (по девяти предъявлениям) при световом раздражении; И — при кожном раздражении (уколе); /// и IV — при сочетании светового раздражения с у кол ом соответственно при ! —9-м и 10— 18-м их сочетаниях; V — график изменения латентного периода ЭМГ сдвигов. На графиках / — IV: по оси абсцисс — время в с; по оси ординат—средняя частота в имп/с при Д( = 100 мс; горизонтальной штриховой линией указан уровень фоновой активности, на графике V — по оси ординат — порядковые номера сочетаний; по оси абсцисс— время в с; вертикальными штриховыми линиями А и Б и стрелками указаны соответственно момент светового раздражения и момент укола. Калибровка: 500 (верхний луч) и 250 (нижний луч) мкВ, 50 мс. Графики и осциллограммы представлены в едином временном масштабе
чивающей условно-рефлекторный процесс, надо по крайней мере знать: любые или Вполне определенные изменения реакций нейронов соответствуют новой сигнальной значимости сенсорных стимулов и в каких образованиях мозга эти изменения проявляются наиболее отчетливо. Для того чтобы установить связь нейронных перестроек именно с процессами «сцепления» условного и безусловного стимулов, необходимо исключить возможные другие причины. К таковым могут относиться:
1) возможность изменения эффективной интенсивности сенсорного стимула;
2) влияние на биоэлектрическую активность мышечных артефактов при поз-ных перестройках и выполнении движений и 3) увеличение общей неспецифической возбудимости, так называемой тонической составляющей ассоциативного процесса (по Б. И. Котляру, 1977).
При регистрации реакций нейронов разных образований мозга во время формирования условного рефлекса возникает вопрос об их первичности. Конечные изменения на уровне одного нейрона могут быть одинаковыми независимо от первичности или пторичности их характера. Поиск наиболее существенных для обучения структур мозга заключается в определении локализации тех нейронных популяций, где изменения происходят в первую очередь. Критерием вовлеченности должен являться предшествующий характер реакций нейронов относительно поведенческой реакции, из чего логически вытекает необходимость существования «критических синапсов», в которых прежде всего происходят изменения, обусловливающие распространение импульсации по новому пути. Причем наряду с сигнально-детсрминированными реакциями описан класс сдвигов импульсной активности, которые также возникают по механизму воспроизведения следов ранее выработанных связей, но без непосредственного участия внешних сигналов — центрально-детерминированные реакции (М. Я. Рабинович, 1975). Именно эти реакции могут послужить удобной моделью для иссле-
дования механизмов произвольной двигательной деятельности.
Структуры мозга, в которых обнаруживаются первичные сдвиги нейро-нальной активности, согласно Дж. Олд-су, представлены в разных отделах мозга. Их мйжно разделить на неспецифические и специфические. К первой группе отнесены те образования, в которых нейроны реагируют сходным образом на условные и любые сходные стимулы той же модальности, но отличающиеся от первых по своим физическим параметрам.
В специфических отделах нейроны не проявляют черт генерализации в реагировании и не имеют сходных реакций на новые стимулы. В качестве наиболее значимых для обучения отделов мозга отмечены фронтальная и сенсомоторная кора, а также гиппокамп, где обнаружены изменения импульсной активности пирамид слоя САз уже при первых сочетаниях условного и безусловного стимулов, и разряд нейронов предшествовал двигательной реакции.
Группа американских ученых под руководством Ч. Вуди (1982) исследовали внутриклеточно реакции нейронов сснсомоторной коры кошек при выработке мигательного условного рефлекса на звук. Безусловным стимулом служило механическое постукивание по переносице. Реакции нейронов сеисомо-торной коры предшествовали появлению первых мигательных движений. В результате обучения наблюдалось, с одной стороны, избирательное снижение порогов миографических реакций круговой мышцы глаза, осуществляющей мигание в ответ на поверхностную или внутрикорковую микростимуляцию сен-сомоторной зоны, а с другой — избирательное усиление реакции и увеличение числа реагирующих нейронов, проецирующихся к соответствующей мышце. Было установлено, что пороги генерации спайков на внутриклеточно приложенный импульс тока достоверно ниже у тех нейронов, которые проецируются к мышцам, осуществляющим реакцию мигания, по сравнению с проекционными нейронами других лицевых мышц. Условные реакции нейронов на
звуковой сигнал появляются с коротким латентным периодом, не превышающим латентный период реакций клеток первичной слуховой коры, и могут сохраняться в течение длительного времени — 14 недель. Указанные функциональные сдвиги нейронов сенсомотор-ной коры были результатом только выработки условного рефлекса. Функциональное выключение сенсомоторной области аппликацией хлорида калия вызывает утрату реакций этих нейронов и соответственно исчезновение самого акта мигания, хотя безусловная мигательная реакция не страдает. Результаты этих экспериментов свидетельствуют о первичности пластических изменений в нейронах коркового представительства двигательной условной реакции.
В пользу локализации пластических изменений в сенсомоторной коре свидетельствуют также результаты анализа нейронных механизмов условной реакции постановки конечности на опору у кошек (Б. И. Котляр и др., 1983). Под этой реакцией понимают последовательность движений, вызванных прикосновением предмета к дорзальной поверхности лапы, обеспечивающих постановку лапы на поверхность этого предмета. Сигнальная условная реакция, вызванная тактильной стимуляцией вентральной поверхности передней конечности, имела то же двигательное проявление, что и исходная реакция, развивающаяся при раздражении дорзальной поверхности лапы (безусловная) у необученных животных. В результате обучения возбудительный ответ нейронов в корковом представительстве бицепса плеча мышцы, выполняющей основную роль в сгибании локтевого сустава, на условный раздражитель был в 2—2,5 раза больше, чем ответ на тот же раздражитель у контрольных животных. Этот эффект не сопровождался изменением сенсорного притока в сенсомоторную кору.
0 Таким образом, реорганизация сенсомоторных связей, достигаемая путем обучения, состоит в изменении эффективности связей между нейронами сенсомоторной коры: 1) получающими
сенсорно обусловленное эфферентное возбуждение и 2) контролирующими сокращениями определенных групп мышц.
Внутрикорковые системные механизмы временных связей. Технические условия ограничивают возможности одновременной регистрации активности большого числа нейронов. Чаще всего используют регистрацию активности одного, реже — двух-трех нейронов при формировании временных связей. Вместе с этим важно при анализе характера имиульсации такого нейрона исходить из предпосылки его обязательного участия в составе определенной нервной сети.
У. Г. Гасанов (1981) полагает, что методическим приемом для выявления такой элементарной микросистемы является одновременная регистрация нескольких клеток с последующей выборкой отдельных импульсных потоков и статистический анализ перекрестных интервалов между импульсами двух и трех выделенных рядов.
9 Он выделяет следующие особенности взаимосвязной деятельности нейронов при обучении. 1. Деятельность микросети качественно отличается от индивидуальной активности входящих в его состав нейронов. 2. Системная деятельность корковых нейронов при разномодальных длительных раздражениях, а также на разных стадиях выработки условного рефлекса и при угашении его основывается на разных формах взаимоотношений между нейронами. 3. Микросистема основывается на дифференцированном участии в ней различного класса нейронов, по-разному меняющих свои сетевые свойства при обучении.
Предполагается, что в основе изменений реакций нейрона при обучении лежат механизмы неспецифического характера. Локальные же микросети нейронов включаются в более обширные многоуровневые системы общецеребрального характера. Для выявления собственно корковых процессов формирования временных связей необходима либо внутриклеточная регистрация, которая но своей сути не может
быть длительной, либо использование строго локальных воздействий на исследуемый нейрон, например с помощью микроэлектрофореза различных биологически активных соединений. Такая форма эксперимента предъявляет особые требования к самой процедуре условно-рефлекторного обучения, состоящей в том, что и формирование, и угашение временной связи осуществляется на протяжении одного и того же опытного сеанса при параллельной регистрации импульсной активности одного и того же нейрона.
Мигательный условный рефлекс отвечает таким требованиям. В. С. Сабанов (1985) основное внимание обратил на самые ранние этапы формирования временной связи, когда внешних проявлений условной двигательной реакции еще не наблюдалось. При сочетании ритмического звукового и механического (задув воздуха на роговицу глаза) раздражений наблюдалось два типа перестроек нейрональной активности: активация нейрона на условный сигнал, к которому данный нейрон был не реактивен, и тормозной: на условный сигнал появляется тормозная реакция, которая до сочетаний регистрировалась лишь в ответ на предъявление безусловного сигнала (рис. 49). Эта тормозная реакция вырабатывается быстро и угашается при изолированном действии условного сигнала. Характерно, что клетки с активирующим типом реакции были исходно полисенсорные, т. е. реагировали на условный и безусловный сигналы, а клетки с тормозным типом реагирования на условный сигнал были исходно моносенсорными и становились полисепсорными лишь в процессе обучения. Исходно бисенсорные клетки с активирующими условными реакциями отражают результат ассоциативного процесса, сложившегося в результате сочетаний конвергирующих стимулов на докорковых уровнях интеграции,— статическая проецируемая конвергенция, тогда как тормозные условные реакции нейронов обусловлены внутри-корковыми механизмами конвергенции и интеграции — локальная динамическая конвергенция. Вопрос о месте фор-
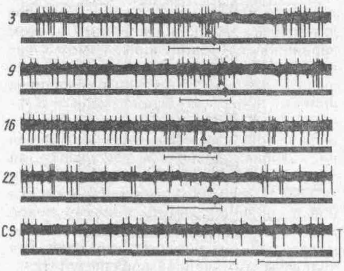
i'nc. 49, Динамика импульсной активности коркового нейрона с тормозным типом условной реак
ции (по В. С. Сабанову 1984):
тонкими горизонтальными линиями отмечено время нанесения звуковых щелчков, момент нанесения безусловного стимула (БС) — струи воздуха на роговицу глаза; цифры — порядковые номера сочетаний; верхний луч — импульсная активность нейрона; нижний луч — электромиограмма мышц глаза; CS — изолированное предъявление условного сигнала. Калибровка: 1 с и I мкВ
мирования различного рода условных клеточных реакций имеет принципиальное значение.
Рассмотрим схему на рис. 50, из которой видно, что таламический эффе-рент, передающий безусловную стимуляцию (БС), через звездчатую клетку, вступая в кору (2), приводит к активации тормозного интернейрона (3), непосредственно контактирующего с исследуемой крупной пирамидой {/). По мере выработки временной связи данный нейрон начинает реагировать аналогичной тормозной условной реакцией и на условный сигнал (УС). Следовательно, ранее подпороговый вход, являющийся проекцией талами-ческой импульсации для условного сигнала, становится пороговым для активации цепи нейронов, предпоследним звеном которой является также тормозной интернейрон (3), контактирующий с исследуемой пирамидной клеткой ( /). Одновременно с этим в систему включен еще один тормозной интернейрон (4), блокирующий проведение импульсации от безусловного стимула к исследуемому нейрону: ответ этого

Рис. 50. Схема связей нейронов для объяснения происхождения тормозных условных клеточных реакций:
/ — пирамидная клетка глубоких слоев коры; 2— звездчатая клетка; 3 и 4 — тормопный интернейрон; 5 — ветви апикальных дендритов в поверхностном слое коры; 6 — коллатерали аксонов; УС — путь проведения условного стимула от соседнего нейронного модуля; RC — путь проведения безусловного стимула из таламуса
нейрона на безусловный раздражитель исчезает. По-видимому,
определяющая роль в ассоциативном обучении принадлежит внутрикортикальным связям между возбуждающими и тормозными интернейронами, динамика которых может обусловливать множество пластических перестроек, лежащих в основе системной организации условно-рефлекторного процесса.
Наблюдая ряд электрофизиологических закономерностей при выработке условного рефлекса, Г. И. Шульгина (1978) высказала гипотезу о том, что одним из основных механизмов действия подкрепляющего стимула является ослабление тормозных — гиперполяризационных и усиление деполяризацион-ных процессов в коре головного мозга. Предвозбудительное торможение в активности корковых нейронов при ассоциативном обучении способствует последующей активации того же нейрона по механизму «отдачи», что обеспечивает синхронное срабатывание большого числа нейронов одной популяции (автостабилизирующее превентивное торможение, по II. В. Симонову).
Рассматривая нейрон с множеством его входов в составе сложных много-
нейронных конструкций, включающих .тормозные иптернейроны, наряду с механизмами локального облегчения синаптической эффективности следует признать существование одновременно работающего механизма подавления избыточных в данный момент синапти-ческих входов. Оба механизма работают одновременно и облегчение синаптической эффективности в определенных локусах постсинаптической мембраны сопровождается торможением прочих эфферентных входов, конвергирующих на один и тот же нейрон.
Постсинаптическая мембрана представляет собой сложную динамическую мозаику возбужденных и заторможенных л о кусов, которые, будучи соче-таннымн во времени с активацией эндогенных клеточных механизмов, фиксируются, обеспечивая вовлечение нейрона в состав общецеребральной нейродинамической системы.
Оригинальная идея выдвинута В. М. Сторожуком (1986), который считает, что решающая роль при обучении принадлежит неспецифическим образованиям мозга, в частности центральному серому веществу. На уровне коры основные пластические изменения происходят в синапсах, воспринимающих неспецифическую афферентацию, тогда как специфические входы не подвергаются существенным функциональным перестройкам. Проводится мысль о модуляции холинергических возбудительных постсинайтических потенциалов в корковых нейронах через систему неспецифических облегчающих и тормозящих влияний. При комбинации специфических и неспецифических входов корковый нейрон, осуществляя перестройку неспецифического входа, приобретает способность участвовать в точно локализованной импульсной реакции в ответ на специфическое, но ранее индифферентное раздражение.
Динамическая констелляция центров. Разрабатывая свое учение, А. А. Ухтомский отмечал, что за каждой естественной доминантой кроется возбуждение целого созвездия (констелляции) центров, своеобразная циклическая машина, функциональный орган, образующийся по ходу самой реакции. При этом определяющим фактором является усваивание ритма, и темпа воз-
буждения в центрах, входящих в состав констелляции.
При исследовании структур, вовлеченных в обеспечение условно-рефлекторного процесса, пришли к заключению о невозможности его привязки к узколокальным отделам мозга. Наоборот, они имеют широкое представительство в самых различных отделах мозга. Тем не менее подтверждается роль коры больших полушарий как высшего этажа головного мозга, ответственного за формирование новых межцентральных функциональных взаимоотношений в процессе обучения. М. Н. Ливанов (1972) разработал концепцию о пространственной синхронизации потенциалов различных отделов головного мозга как основе формирования констелляции центров. По мере становления временной связи были выявлены синхронные «усвоенные» колебания биопотенциалов. С помощью метода корреляционно-спектрального анализа обнаружено значимое увеличение кросскорреляци-онных коэффициентов разных структур мозга на стадии генерализации условного рефлекса. При закреплении и специализации условного рефлекса указанные отношения сохранялись только между теми мозговыми структурами, куда адресовались сочетаемые раздражители — условный и безусловный. У гашение условного рефлекса сопровождалось снижением корреляционной связи между их электрическими потенциалами.
Было установлено, что высокий уровень пространственной синхронизации сопровождается повышением возбудимости и облегчает процессы распространения возбуждения в мозговых системах. Уровень пространственной синхронизации корковых потенциалов обеспечивается регулирующими влияниями из глубоких отделов мозга.
Р. Джон (1978) обнаружил во многих структурах мозга кошки при образовании условного рефлекса на ритмические световые вспышки изменения формы поздних компонентов вызванных потенциалов, коррелирующих с двигательным условным ответом. Самопроизвольное возникновение выработанного
движения сопровождалось появлением подобных же электрографических феноменов. Это позволило думать об их связи с процессом считывания, извлечения следа памяти, широко распределенного по многим аппаратам мозга.
В последние годы в связи с развитием представлений о модульной организации мозга все большее число ученых соглашаются с тем, что отдельные модули как структурно-функциональные единицы входят в состав более крупных объединений и могут вовлекаться в различные формы мозговой деятельности.
В. Маунткасл (1981) выдвинул теорию распределенных систем мозга, которая базируется на трех основных факторах: 1. Основные структуры головного мозга построены по принципу повторения одинаковых многоклеточных единиц — модулей; по своему качественному характеру функция обработки информации в коре больших полушарий одинакова в разных ее областях. 2. Внешние связи между крупными образованиями мозга многочисленны, избирательны и специфичны. 3. Вся группа модулей разбита на подгруппы, из которых каждая соединена своей системой связей с такими же обособленными подгруппами в других отделах мозга. «Связанные между собой группы модулей нескольких структур мы называем распределенными системами... Крупные структуры мозга как части распределенных систем придают каждой из них свойство, определяемое связями, общими для всех подгрупп модулей этой структуры, и специфическим качеством их внутренней активности... Таким образом, модули распределенных систем связаны между собой в «эшелонированные» параллельные и последовательные объединения. Такая распределенная система характеризуется избыточностью потенциальных командных пунктов, и командная функция в разное время может локализоваться в разных участках системы, особенно там, куда приходит саман срочная и нужная информация. Поэтому у такой системы много входов и выходов, а функция как свойство ее
динамической активности заключена в системе как таковой»' (подчеркнуто мной.— А. Б.).
В ' вышеприведенных построениях обходится молчанием вопрос о нейрофи-зиологических механизмах «сборки», формирования и поддержания столь обширных нейронных объединений в единой рабочей системе. На этот ключевой момент обращает внимание Дж. Эк-клс (1981), обсуждая механизмы корковой межмодульной интеграции. Автор считает, что корковый модуль может участвовать в неограниченном числе сложных систем модулей, которые он именует пространственно-временными паттернами.
ф В основу своей гипотезы, в значительной мере опирающейся на морфологические схемы Я. Сснтаготаи, Дж. Экклс положил три постулата. 1. Имеет место точная проекция из одного коркового модуля к нескольким обособленным группам модулей в том же полушарии и к зеркальному фокусу в противоположном полушарии. Последние по обратным комиссуральным и ассоциативным связям проецируются на тот же начальный модуль, обеспечивая предпосылки для колец реверберации как внутри-, так и межполушарной. 2. Такие же кольца реципрокных связей существуют между корой больших полушарий и таламусом, которые обеспечиваются крупными пирамидами V слоя. Эти кольца могут служить основой для таламо-кортикальной реверберации, в результате чего активируются (с помощью таламо-кортикаль-ных входов) касательные синапсы на шипиках апикальных дендритов пирамидных клеток слоя V. 3. Апикальные дендриты крупных корковых пирамидных клеток образуют в I и II слоях касательные синапсы с проходящими горизонтально аксонами, которые принимают участие в механизмах формирования и считывания следов памяти (рис. 51, а).
Эти положения позволили высказать гипотезу, согласно которой при временном совпадении активации апикальных
Эйдсльман Дж., Маунткасл В. Разумный мозг. М., 1981. С. 57—58.
дендритов пирамид через касательные синапсы и проведении импульсации по горизонтальным аксонам I слоя происходит гипертрофия ограниченной популяции синапсов этих горизонтальных волокон с апикальными дендритами тех же самых пирамид этого модуля. На рис. 51, б толстыми линиями показано состояние, когда два соседних модуля активированы корти ко-кортикальным и и/или таламо-кортикальными входами, многие из радиально расходящихся от них волокон потенциируют синаптичес-кис контакты с дендритами пирамидных клеток в слоях I и II в других модулях. Сама по себе активация синапсов в слоях I и II весьма незначительна в связи с удаленностью от триггерной зоны, но она может быть увеличена благодаря включению пирамиды V слоя в реверберационное кольцо. Такая продолжительная активация значительно усиливает возбуждение касательных синапсов, которые выполняют ключевую роль в гипотезе Экклса о гипертрофии синапсов горизонтальных волокон с апикальными дендритами пирамид (возможно, это шипико-шипиковые синапсы). В одно и то же время шипиковые звездчатые клетки переключают импульсацию на дендриты пирамидного слоя V и вызывают растормаживающее действие на нейронный модуль. Так, может усиливаться слабый сигнал, поступающий к апикальным дендритам, и тем самым осуществлять избирательную активацию нейронного модуля.
Учитывая возможность связи модулей по верхнему плексиморфному слою коры с помощью горизонтального ветвления апикальных дендритов и по глубоким слоям коры с помощью горизонтальных коллатералей аксонов пирамидных нейронов, следует принять во внимание аксо-дендритические и депд-ро-дендритические связи с апикальными и базальными дендритами пирамид, которые в основном устанавливаются с их дендритными выростами-шипика-ми. Это наиболее пластичное соединение между нейронами может быстро менять эффективность синаптической передачи. Количество самих шипиков и

Рис. 51. Упрощенная схема связей в неокортексе в соответствии с теорией церебрального обучения Экклса {1981) (a) и схема больших объединений модулей в горизонтальном плане (б)
а — горизонтальные волокна, возникающие- как продукт бифуркации аксонов ком несу раль-пых (Ком) и ассоциативных (ас) волокон, а также аксонов клеток Мартинотти (Ма), образуют синапсы с апикальными дендритами звездчатых клеток в модуле В и пирамидных клеток в модулях А и Б. Внизу модуля А показана шипикпвая звездчатая клетка (ЗК) с аксоном, образующим касательные синапсы (КС) со стволом апикального дендрита пирамидной клетки (ПН). Обусловленные гипертрофией (в результате совпадения импульсных влияний) ассоциативные волокна из модуля В увеличивают синапсы на апикальных деидритах пирамидной клетки к модулю А. Каждый модуль показан кружком. Из двух модулей образуется множество горизонтальных радиальных волокон, которые уходят за пределы показанного участка. Каждый модуль претерпсиа-ет непременную бифуркацию в слое I и II гак, что количество радиальных вюлокон удваивается в сравнении с числом вертикальных входов — ассоциативных, каллозальных и клеток Мартинотти. В некоторых модулях происходи! гипертрофия синапсом благодаря активации проходящих горизонтальных волокон (утолщенные линии). Такие горизонтальные волокна распределяются неравномерно, а преимущественно в каком-то одном направлении.
синапсов на них увеличивается в онтогенезе и прямо зависит от накопления опыта, т. е. от образования энграмм памяти. Синаптическое соединение с шипиком является структурно весьма стабильным, а функционально — подвижным. Последнее достигается с помощью изменения геометрических параметров самого шииика, что
меняет сопротивление для распространяющихся по нему синаптических местных потенциалов в направлении к стволу дендрита. Таковыми могут быть некоторые гипотетические механизмы формирования корковой пространственно-временной структуры --- матрицы памяти.
Ч. Вуди показал, что при типичной
ситуации обучения у ненг."котизирован-ных кошек в постсинаптической клетке изменяется сопротивление мембраны, которое сохраняется в течение недель и сопровождается устойчивым увеличением возбудимости клеток. В. М. Смирнов, Ю. С. Бородкин (1979) также обнаружили на целом мозге во время формирования искусственных связей при сочетанной электростимуляции мозговых структур устойчивое падение импеданса между ними.
Такие наблюдения позволяют допустить, что одним из механизмов создания пространственно организованной динамической констелляции мозговых аппаратов является регулирование сопротивления постсинаптичсских мембран нейронов, которые вовлекаются в такую констелляцию. Длительное сохранение этих изменений свойств мембран, а следовательно, связей между отдельными нейронными популяциями обеспечивается регулирующей ролью циклических нуклеотидов (вторичных посредников) самих нейронов.
Исследованиями на бодрствующих обезьянах (А. С. Батусз и др., 1988) было установлено, что к категории наиболее подвижных и пластичных мозговых систем следует отнести ассоциативные системы мозга, динамическая корковая констелляция которых складывается по ходу программирования целенаправленного действия животного и достигает наиболее высокого уровня интеграции к моменту его выполнения. Такая динамическая констелляция и лежит в основе доминанты вместе с ее условно-рефлекторным обеспечением.