

4.У вартових, що стоять струнко на варті, швидко виникає втома. Який вид фізичної роботи має місце? Які можливі механізми розвитку втоми у зазначених умовах?
В цьому випадку, по-перші, переважає ізометричне скорочення м’язів і, відповідно, статичне фізичне навантаження. Таке навантаження потребує значного і тривалого напруження м’язів і, відповідно, значних енерговитрат. За 4-м правилом Фольборта, чим інтенсивніше робота і швидше виникає виснаження, то й швидше відновлення після роботи. Статична «робота» вартового не досягає такої високої інтенсивності, яка б спонукала відповідні процеси відновлення. При такій роботі не відбувається швидкої і високої мобілізації серцево-судинної системи і системи дихання – отже, найбільш економний і інерційний шлях ресинтезу АТФ – аеробне окислення глікогену та ліпідів – не стимулюються з такою ж високою інтенсивністю, як при динамічному фізичному навантаженні. Залишаються креатин-фосфатний, занадто короткочасний шлях відновлення АТФ і шлях анаеробного гліколізу, який супроводжується утворенням недоокислених продуктів (піровиноградної та молочної кислот). Таким чином, відбувається «засмічення» і пригнічення роботи м’язів. До того ж, специфіка цієї роботи полягає у значному нервовому напруженні. Нервові центри у даних умовах досить тривало забезпечують підтримання рівноваги в умовах нетипового розподілу тонусу, довільної уваги в умовах монотонних сталих подразників, тривалих вольових зусиль. Це зумовлює мобілізацію ЦНС на всіх рівнях, у тому числі і кори великих півкуль. Отже, досить швидко виникає втома на всіх рівнях – на рівні нервових центрів, на рівні нейро-органних медіаторів і на рівні м’язів, в яких провідну роль відіграють метаболічні процеси.
5.Відновлення пози тіла і голови забезпечує рефлекс випрямлення. Це різновид статичних тонічних рефлексів, які спрямовані на підтримання рівноваги у відсутності руху с прискоренням і полягають у відповідному розподілі тонусу для досягнення цієї мети. Інформація про зміну положення голови передається від рецепторів отолітового апарату, а також від пропріорецепторів м’язів шиї. Допоміжну роль можуть відігравати тактильні рецептори шкіри тулубу (у місці перетину шкіри при повороті голови, з якого починається відновлення шкіри) і зорові рецептори. До локального центру цього рефлексу надходять мотонейрони спинного мозку, структури заднього і середнього мозку. Без середнього мозку у бульварної тварини цей рефлекс не відбувається.
6.Якщо людина «завалюється» після обертання ліворуч на випрямлених ногах, то це свідчить що саме в цьому напрямку здійснювалось швидке обертання і тому виникла активація переважно лівого ядра Дейтерса та інших вестибулярних ядер. Отже, з лівого боку домінує тонус розгиначів. В певних випадках після обертання ліворуч людина може відхилятися на контр-латеральний бік – у напрямку реціпрокно домінуючих згиначів (з протилежного обертанню боку). Але при цьому вона рухається більш плавно, а не завалюється, як в розглянутому варіанті.
При невисокій швидкості руху може перемагати збудження ядра Дейтерса і розгиначів на боці, протилежному напрямку обертання і, відповідно, реціпрокно активуються згиначі на боці обертання. У цьому випадку людина після обертання зазвичай рухається плавно, відхиляючись на бік обертання, тобто в напрямку домінуючих згиначів.
Підстави для ствердження, що перерозподіл тонусу під час обертання зв’язаний, перш за все, зі збудженням ядра Дейтерса, полягають у тому, що обертання є єдиним адекватним подразником для рецепторів пів-колових канальців (або, ампулярних рецепторів). Останні представляють собою ту частину вестибулярного апарату, в якій генерується досить сильне збудження, здатне до іррадіації в різні нервові центри. Проте, безпосередні зв’язки цих рецепторів проектуються УІІІ парою на вестибулярні ядра. А ядро Дейтерса – одне з цих ядер, до того ж найбільш впливове. Рефлекс, що виникає з ампулярних рецепторів і забезпечує перерозподіл тонусу для збереження рівноваги, називається тонічним стато-кінетичним рефлексом (на кутове прискорення). Такі рефлекси спрямовані на підтримання рівноваги в умовах руху с прискоренням. До структур, що надходять до локального центру цього рефлексу, належать мотонейрони спинного, мозку, структури заднього мосту (зокрема, ядро Дейтерса довгастого мозку і ретикулярна формація мосту), структури середнього мозку (зокрема, червоне ядро). Для таких тонічних рефлексів характерне супроводження їх типовими фазичними рефлексами – ністагмом голови, очей (призначення – швидка оцінка змін у просторі).
7. Якщо у сечі збільшена концентрація фосфатів і зменшена концентрація Са²+, то реабсорбція Са²+ у нирках була збільшена, а реабсорбція фосфатів –
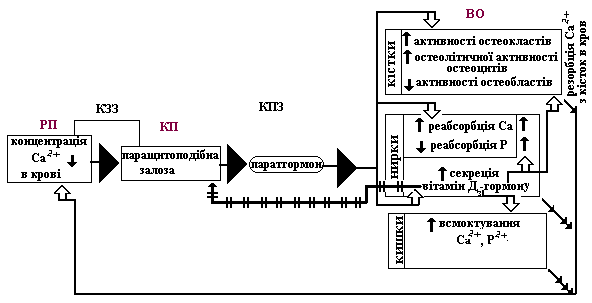 зменшена.
Саме так діє в нирках парат-гормон
(паратиреїн) – гормон пара-щитоподібної
залози. Причиною виділення цього гормону
є зменшення концентрації Са²+ у крові
(збільшення у крові концентрації Р²+).
Саме рівень цих іонів у крові (особливо,
Са²+) – є регульованим параметром.
зменшена.
Саме так діє в нирках парат-гормон
(паратиреїн) – гормон пара-щитоподібної
залози. Причиною виділення цього гормону
є зменшення концентрації Са²+ у крові
(збільшення у крові концентрації Р²+).
Саме рівень цих іонів у крові (особливо,
Са²+) – є регульованим параметром.
Але таке «недобре» ставлення паратгормону до фосфору компенсується впливами вітамін-Д3-гормону, продукція якого в нирках стимулюється тим же парат-гормоном. Вітамін-Д3-гормон збільшує реабсорбцію з ниркових канальців в кров і іонів кальцію, і фосфору. Отже, такий характер сечі може свідчити про недостатність продукції вітамін-Д3-гормону. Одной з причин цього може бути недостатнє засоєння попередників цього гормону в шлунково-кишковому тракті.
8.Екзогенні андрогени (тестостерон), які всмоктується у кров при вживанні, сприймаються за каналом негативного зворотного зв’язку як складова крові, що є РП. Отже, зменшується секреція гонадолібреринів гіпоталамусом і ЛГ гіпофізом і, як наслідок зменшується власна секреція тестостерону. Зменшення секреції гонадоліберинів, зі свого боку призводить до пригнічення секреції і ФСГ, і ЛГ аденогіпофізом. Але „втрати” по лінії ЛГ штучно компенсуються вживанням екзогенного гормону, а нестача ФСГ – ні. Між тим, об’єктом дії ФСГ є клітини Сертолі і сперматогенез, на який ФСГ впливає разом з тестостероном (без ФСГ підтримувати сперматогенез здатні занадто високі дози тестостерону). Можливо, через нестачу ФСГ клітини Сертолі припиняють виділяти особливий фактор росту сім’яних канальців – і саме тому клітини Лейдига при екзогенному прийомі тестостерону атрофуються. Нестача ФСГ означає також недостатню реактивність клітин Сертолі до тестостерону. Таким чином, сперматогенез «втрачає» обидва головних гормональних фактора, що його підтримують. Внаслідок цього сперматогенез припиняється, статева система приходить в занепад і це відбивається на стані яєчок. Отже, екзогенний тестостерон здатний підтримувати лише вторинні статеві ознаки і лібідо.
9.Вираз „діти ростуть, коли сплять” має фізіологічне пояснення – гормон аденогіпофізу СТГ за циркадіанним ритмом секретується більш вночі. Але СТГ лише стимулює печінку до секреції соматомединів. А саме соматомедини забезпечують лінійний ріст і фізичний розвиток. Їх синергісти – йодтиреоїдні гормони (осіфікація кісток). Органи-мішені – кістки (перш за все, епіфізарні частини), скелетні м’язи (де соматомединам синергує також і СТГ), печінка. Певною мірою, синергістами соматомединів є статеві гормони. Нестача естрогенів супроводжується остеопорозом та іншими дефектами фізичного розвитку. Тестостерон, з одного боку, стимулює секрецію СТГ, збільшує масу м’язів, але з іншого сприяє заростанню епіфізів кісток (отже, висока концентрація тестостерону може супроводжуватися обмеженням росту скелету).
За контуром регуляції порушення секреції СТГ (збільшення, зменшення) можуть бути зв’язані з порушенням виділення соматоліберину (при зменшенні – зменшення секреції СТГ) і соматостатину (при збільшенні – зменшення секреції СТГ) гіпоталамусом; дефектом рецепторів гіпофізу до соматостатину і соматоліберину; або (і частіше) патологією самого гіпофізу, що супроводжується гіпер- або гіпопродукцією СТГ – звідси гіпофізарний нанізм або гігантизм; або з нестачею (надлишком) соматомединів; й нарешті, з дефектом рецепторів до соматомединів у гіпофізі або гіпоталамусі.
10.АКТГ є одним з так званих тропних гормонів аденогіпофізу. Основним напрямком дії цього гормону є перш за все, пучкова (глюкокортикоїди), сітчаста (статеві гормони) і (меншою мірою) клубочкова (мінералокортикоїди) зони кори наднирникових залоз. Крім цього є побічні ефекти АКТГ – зменшення лімфатичних вузлів, селезінки, щитоподібної залози, пригнічення синтезу і збільшення катаболізму білку в організмі, покращення настрою. Найбільш впливовим цей гормон є для пучкової зони (секреція альдостерону в першу чергу регулюється концентрацією іонів натрію/калію, а ні АКТГ). Отже, секреція АКТГ за каналом негативного зворотного зв’язку регулюється в залежності від рівню кортизолу у крові. У клітинах кіркової речовини наднирникових залоз АКТГ посилює окислювальне фосфорилювання і збільшує синтез білків, активує глюкогенез, збільшує вміст аскорбінової кислоти і холестерину, що використовуються для синтезу кортикостероїдів, і, як результат, збільшую синтез і секрецію кортикостероїдів. Це водорозчинний гормон, який використовує як посередника своєї дії на клітини цАМФ.
11.Концентрація кортизолу збільшується під час 2-ї фази стресу, коли адреналін, що виділився до крові у масивних кількостях (під час 1-ї фази), запобігає реалізації негативного зворотного зв’язку кортизол-гіпоталамус-аденогіпофіз, бо під впливом адреналіну збільшується секреція АКТГ аденогіпофізом. Кортизол утворюється у пучковій зоні кори наднирникових залоз. Це жиророзчинний гормон, що проникає через мембрану у клітину і прямує до ядра клітини-мішені. Тобто механізм дії на клітину опосередкується через ДНК, т-РНК. Гіперглікемія, під впливом кортизолу, переважно обумовлена за рахунок глюконеогенезу, хоча цей гормон здатний і до глікогенолізу.
Відомо, що в умовах стресу, перш за все, дає результат мобілізація симпато-адреналової системи: зростає активність симпатичної нервової системи і з мозкової речовини мобілізується адреналін у кров (оскільки ця ланка стресу реалізується переважно швидкісним нервовим шляхом, а гіпоталамо-гіпофізарно-наднирникова – гуморальним). Симпатичні нервові впливи і гуморальні впливи адреналіну на β-клітини підшлункової залози носять гальмівний характер. Отже, секреція інсуліну в таких умовах гальмується.
12.Альдостерон утворюється у клубочковій зоні кори наднирникових залоз. Це жиророзчинний гормон, який проникає у клітину і прямує до її ядра. Зокрема, унаслідок дії гормону на білковий синтез, у епітелії дистальних канальців нефрону збільшується активність натрій-калієвої АТФ-ази і натрій-калієвого насосу – іони натрію повертаються з канальців у кров, а іони калію, навпаки, секретються з крові у просвіт канальця – отже, концентрація іонів натрію в крові збільшується, а калію зменшується (з цим ефектом зв’язані зміни водно-сольового обміну і про-запальні /сприяючі запальним процесам/ ефекти альдостерону).
13.Тривалий біль у спортсмена у м’язах передають ноцицептори повільного болю, які є вільними закінченнями С-волокон. У зазначених умовах їх подразниками являються хімічні речовини – кислоти (молочна, піровиноградна та інші, іони К, простагландини, тощо). Провідним шляхом такого болю є палеоспіно-ретикуло-таламо-кортикальний шлях (див. схему на початку файлу), а провідним медіатором субстанція Р.
14. За теорією Мелзака, Уолта (1965), яка названа теорією воротного контролю спінальної переробки ноцицептивної інформації гальмування сенсорних ноцицептивних нейронів задніх рогів обумовлено збудженням товстих неноцицептивних аферентів (ворота зачинені), а активація їх – збудженням тонких ноцицептивних аферентів (ворота відкриті); вказане гальмування, згідно з цією теорією, генерується у гель-подібній речовині заднього рогу спинного мозку й забезпечується виключено пресинаптичним гальмівним механізмом, який діє на тонкі ноцицептивні аференти. Крім такого висхідного контролю відбувається низхідний (центробіжний) антиноцицептивний контроль. Відомі два шляхи такого контролю: 1)серотонінергічні нейрони шву і холінергічні нейрони гігантоклітинного ядра ретикулярної формації є джерелами шляхів, що прямують до енкефалічних гальмівних нейронів (розташованих у гель-подібній речовині спинного мозку), які вже безпосередньо гальмують спінальні ноцицептивні нейрони – отже, опосередковують свої ефекти через опіатну систему; 2)норадренергічні нейрони блакитної плями є джерелом шляхів, що через спинний мозок прямують до спинномозкових гангліїв і здійснюють гальмівний вплив на ноцицептивні шляхи в задніх корінцях спинного мозку. Оскільки в ноцицептиних нейронах спинного мозку наявні опіатні циторецептори, низхідний контроль також здійснюється через опіатну систему.
15.Людина відчуває раніше дотик, ніж тепло, оскільки від низько-порогових рецепторів дотику інформація до відповідної зони кори йде найбільш швидкісним лемнісковим шляхом, а від теплових рецепторів –спіно-таламо-кортикальним, да ще по низько швидкісним-волокнам типу С – див.схеми.
16.Якщо у людини нормальна гострота зору (1), то це свідчить про те, що максимальна здатність ока сприймати/розрізняти близько розташовані об’єкти як окремі, у нормі. Гостроту зору можна відносити до категорії просторового розрізнення, а її показник – до своєрідного порогу просторового розрізнення на сітківці. Але вплив зорових подразників на фоторецептори сітківки здійснюється після переломлення у оптичній системі ока. Отже, нормальна гострота зору свідчить про відсутність аномалій рефракції і нормальне переломлення променів у цій оптичній системі. До того ж, це – один з численних критеріїв нормального стану всієї зорової сенсорної системи – починаючі від нейтронного апарату сітківки ока і до ЛКТ і шпорної /або острогової/ борозни у потиличній корі.
17.Нормальне поле зору (простір, який охоплюється одним оком при фіксованому стані очного яблука) є одним з численних критеріїв нормального функціонального стану сітківок. Зокрема, щодо стану сітківки, то вивчення поля зору дозволяє оцінити перш за все стан паличкового апарату (периферійний зір), оскільки центр ока фіксований. Центральний зір, що забезпечується колбичковим апаратом і досліджується при використанні кольорових стимулів, має значне менше поле. Нормальні показники поля зору є одним з критеріїв відсутності порушень у зоровому нерві, хіазмі, тракті, ЛКТ (провідному відділі зорової системи) і шпорній /остроговій/ борозні потиличної кори (кірковий відділ), тобто у зоровій сенсорній системі у цілому. Це витікає з того, що при переломленні променів з назальної частини поля зору попадають на зовнішню частину сітківки, а промені зі скроневої частини поля зору – на внутрішню. З зовнішньою частиною сітківки зв’язане ЛКТ і шпорна борозна того ж боку, що і сітківка. А з внутрішньою сітківкою – завдяки перехресту –ЛКТ і потилична кора протилежного боку. Отже, пошкодження зорової системи на цих рівнях відіб’ється на певних показниках поля зору. Таким чином, завдяки аналізу цих показників можна досить точно встановити рівень порушень у зоровій сенсорній системі.
18. Оскільки очі містяться у різних частинах голови, зображення на їх сітківках декілька різні, що слідує з законів геометричної оптики. Чим крупніше предмети і чим ближче вони до спостерігачу, тим більше таке горизонтальне розходження – диспаратність. Горизонтальна диспарантність виступає як кількісна міра різниці між місцями на сітківках, на які проектується зображення об’єкту, розташованого на кінцевій відстані від очей при звичайному положенні голови. Коли диспарантність перевищує певну величину бінокулярне спостереження 3-хмірного предмету призводить до сприймання його як подвійного (просторовий розподіл 2-х зображень при горизонтальній диспарантності можна вимірити при використанні моделі „циклопічного ока”). Таку диспарантність можна створити штучно – дивлячись на предмет і натиснувши на одне око пальцем – зображення стане двоїтися. Діапазон відстаней до об’єкту – від нульової диспарантності (об’єкт у безмежності)і кінцевої межі бінокулярного глибинного зору, при якій сприймання стає подвійним, досить великий, оскільки бінокулярно активований кірковий простір значно більш просторового еквіваленту відповідної ділянки сітківки. Коли обидва ока фіксують точку на певній відстані, всі об’єкти на колі, що йде через вузлові точки оптичних систем двох очей і точку фіксації, проектуються на ідентичні кореспондуючі ділянки сітківок. Всі об’єкти у середині або за межами цього кола (гороптерного кола) на кореспондуючі ділянки сітківок вже не проектуються. Ті з з бінокулярно активованих нейронів кори, в яких збуджені рецептивні поля /РП/ на обох сітківках точно співпадають, максимально збуджуються об’єктами у межах гороптерного кола. Ті з бінокулярно активованих нейронів кори, в яких РП не сітківках точно не співпадають, максимально збуджуються об’єктом, який міститься на певній відстані від гороптерного кола, що призводить до певної горизонтальної диспарантності.
Стереоскопічний /бінокулярний/ зір перевіряється за допомогою бінокулярного погляду на 2 рисунки, що розташовані поруч і виглядають для кожного ока відокремлено як випадкові набори точок (або за допомогою кольорового тесту). Ті, хто здатний бінокулярно корелювати окремі точки з відповідною горизонтальною диспарантністю, здатні сприймати у даному випадку й форму і глибину зображень. Зокрема, при наближенні предмету до спостерігачу фіксація очей на ньому (і, відповідно, проекція його на кореспондуючі ділянки сітківок) досягається конвергенцієй очних яблук. При цьому відбувається акомодаційний рефлекс і звуження зіниці.
Отже, механізми, що забезпечують бінокулярний зір – це спряжені рухові програми (у супроводженні відповідних вегетативних рефлексів), які виконуються, перш за все під контролем очно-рухових центрів стволу мозку. Нейрони, яки здійснюють керування горизонтальних рухів очей, розташовані, головним чином, у пара-медіальній ретикулярній формації (РФ) варолієва мосту (ПМРФ), а керування вертикальними рухами забезпечується у РФ середнього мозку (РФСМ). Аксони нейронів ПМРФ і РФСМ йдуть до рухових ядер черепно-мозкових нерві (ІІІ, ІV, VІ, VІІ пар). Рівень збудження очно-рухових центрів стволу регулюється різними структурами мозку, зокрема верхніми горбиками 4-горбія, вторинною зоровою корою, тім’яною інтегративною корою (7 поле), фронтальним лобним очним полем, префронтальною руховою корою. Зв’язки з руховими центрами ПМРФ і РФСМ також утворюють нейрони вестибулярних ядер, клочку, параклочку мозочку.
19.Слухові рецептори розташовані на базальній мембрані равлику (завитку) під нерухомо закріпленою над ними текторіальною мембраною. Базальна мембрана відокремлює середню драбину від нижньої. А тонка мембрана Рейснера відокремлює середню драбину равлику (завитку) від верхньої. При повітряній провідності звукові хвилі концентруються в вушній раковині і передаються на барабанну перетинку, де перетворюються на механічні. Ці механічні коливання проводяться далі у середньому вусі слуховими кісточками (молоточком, ковадлом і стремінцем) до овального вікна і перилімфи та ендолімфи равлику, що зумовлює коливання основної мембрани (коливання рідини равлику стають можливими завдяки наявності ще одного вікна – круглого). Основна мембрана підкидує волоскові слухові клітини (рецептори), вони упираються волосками у текторіальну мембрану і народжується РП. При кістковій провідності звукові коливання передаються до кісток черепу. Коливання кісток черепу заохочують до коливань рідину і основну мембрану завитку. Але ефективність таких коливань і, відповідно, кісткової провідності значно менш, ніж при повітряній провідності. Якщо людина при закритті вушних проходів ватою (тобто пригніченні повітряної провідності) краще чує більш „тугим” вухом, то це свідчить про те, що на боці „тугого” вуха порушено не сприймання звукових подразнень, а їх повітряне проведення. Це може бути серна пробка у зовнішньому проході або пошкодження барабанної перетинки або слухових кісточок.
20.При виробленні інструментального УР уникнення у щурів виникала внутрішня потреба захисту від болю, або уникнення болю, що викликав електричний струм. Таку потребу слід вважати складовою потреби збереження цілісності організму. А поведінку, що виникає на підставі такої потреби – проявом вітального інстинкту самозбереження. Якщо за допомогою внутрішнього резерву неможливо досягти цієї мети, то виникає біологічна потреба і мотивація, що призводить до поведінкового акту. В даному випадку простий безумовний рефлекс – рефлекторне захисне згинання кінцівок не вирішує проблеми збереження цілісності. Отже має відбутися більш складна форма безумовно-рефлекторної діяльності – драйв-рефлекс, який полягає у пошуку способу уникнення больового подразнення. Для реалізації такої поведінки необхідне критичне збудження відповідної частини гіпоталамуса, і, як наслідок, іррадіація цього збудження на моторні центри кори великих півкуль, а також на підкоркові центри, що беруть участь у руховому і вегетативному забезпеченні поведінки уникнення. Отже, виникає мотивація, яка на структурно-функціональному рівні виявляється у запуску відповідної видової генетично закріпленої поведінкової програми – програми втечі від шкідливого подразнику і пошуку вкриття. Якщо використовується індиферентний сигнал, який попереджає про наступне больове подразнення, то між сенсорною корою, що сприймає дію такого сигналу і кірковими центрами у даному випадку драйв-рефлексу встановлюється тимчасовий зв’язок. Після цього відповідну поведінку вже запускає сигнал попередження – отже виробився умовний рефлекс. При цьому суб’єкт (щур) має здійснити певні власні дії для того, щоб задовольнити свою потребу – саме такі умовні рефлекси називають інструментальними, і за цією рисою вони мають спільність с природженими драйв-рефлексами.
21.Плач і рухова активність у немовляти є різновидом інстинктивної поведінки – а саме – проявом інстинкту самозбереження. Безпорадна дитина лише у такий спосіб може задовольнити свої біологічні потреби, що пов’язані з зовнішніми (і внутрішніми) подразниками. Оскільки її фізичний розвиток не дозволяє здійснити інший поведінковий акт, спрямований на задоволення своїх потреб. Пристосувальна реакція, яку забезпечує така поведінка, сигнального значення – це притягнення уваги дорослих, без яких дитина не здатна задовольнити більшу частину своїх потреб. Відповідну схему можна конкретизувати виникненням відчуття голоду, спраги, холоду, болю – схеми.
22.Тахікардія унаслідок розуміння, що для запланованих дій не вистачає часу, свідчить про виникнення емоційного стану напруги. У даному випадку емоції виконують, перш за все, мобілізуючу і компенсаторну функцію. Це особливо наглядно, якщо розглянути ці події з позиції формули Симонова. СН= -f П (ЕнІнЧн – ЕіІіЧі). Отже, виникнення стенічної емоції, а ще раніш того, як вона буде усвідомлена, - мобілізація симпато-адреналової системи здійснюється, коли існуючих енергії, інформації і часу не вистачає. Під впливом адреналіну звільняється в кров глюкоза і зростає енергетичний рівень функціонування організму(Еі↑). Вплив адреналіну на нервові центри – оптималізуючий: актуалізуються нервові зв’язки, прискорюється обробка інформації, покращується її пригадування і запам’ятовування (Іі↑). Отже, зростання енергетичного потенціалу і прискорення діяльності ЦНС можуть скоротити час, необхідний для виконання дій (Чн↓). Але такі результати відбуваються при умові, що складність завдання співпадає з мобілізованими резервними можливостями суб’єкту – СН-1 (стан напруги, або стенічна емоція 1 ст. – уваги, мобілізації, активності). Якщо мобілізовані резервні можливості менш, ніж пред’явлені суб’єкту вимоги, виникає СН-2, який зазвичай характеризується емоціями люті, гніву і афективною поведінкою (при цьому напруга і мобілізація ресурсів досягає максимального рівня). СН-3 – астенічна емоція відбувається на фоні вичерпаного ресурсу і неможливості досягти поставленої мети. Характеризується реакціями протилежного СН-1, 2 напрямку – погіршення інтелектуальної діяльності, компенсаторних процесів, зменшення енергетичних ресурсів і пригніченим емоційним станом (страх, нудьга). Але СН-3 може мати певну користь як своєрідний захист від не результативного зростання напруги, якщо суб’єкт здатний зменшити суб’єктивно значущість цілі. Якщо ні – організм попадає в безвихідне становище і виникає СН-4 – невроз як захворювання і поломка регуляторних механізмів.
23.Механізм, що дозволяє організму швидко оцінити, чи має місце відновлення гомеостазу за певними конкретними його параметрами – це канал зворотного зв’язку, який починається від регульованого параметру, за станом якого стежать рецептори відхилення (а саме відхилення параметру гомеостазу від заданого рівня формує біологічну потребу). Рецептори відхилення, у свою чергу, інформують про стан параметру контролюючий пристрій (КП). Такий вигляд має канал зворотного зв’язку у контурі нервової регуляції, де КП – це нервовий центр. У контурі гуморальної регуляції рецептори, що сприймають рівень певного параметру гомеостазу, є невід’ємна частина КП, а КП зазвичай представляє собою певну залозу внутрішньої секреції. Отже, за каналом зворотного зв’язку організм інформується про стан регульованого параметру. Однак, якщо внутрішнього резерву організму недостатньо для відновлення того чи іншого параметру гомеостазу й виникає необхідність активного контакту з навколишнім середовищем, виникає так звана біологічна потреба, що проявляється в мотиваційному збудженні певних зон гіпоталамусу, іррадіації його до відповідних зон кори і супроводжується поведінковим актом, спрямованим на задоволення внутрішньої біологічної потреби. Згідно з концепцією Анохіна, програма поведінки включає в себе не тільки «план» дій, але й «план» результату (закодований в акцепторі результату), з яким звіряється результат, отриманий внаслідок поведінки. Отже, на більш високому рівні знов реалізується канал зворотного зв’язку і співставляються параметри реально отриманого результату з параметрами ідеального результату. При цьому цілком зрозуміло, що задоволена внутрішня біологічна потреба перш за все передбачає нормалізацію параметру (параметрів) гомеостазу, що відхилилися і зумовили поведінку. Таким чином, канал зворотного зв’язку контуру нервової та гуморальної регуляції «вписаний» в акцептор результату як його необхідна складова.
24.Швидке утворення УР свідчить про достатню силу збудження. А проблеми з внутрішнім умовним гальмуванням – про недостатню силу гальмування. Таку характеристику за Павловим має сильний нестриманий тип (холерик по Гіппократу). Недостатнє гальмування у такого типу негативно впливає на здатність до переключення з одного виду діяльності на інший. Тобто, рухомість у такого типу страждає як похідний з асиметрично розвинутих властивостей ЦНС. Отже, Павлов у такого типу навіть не характеризує рухомість, оскільки високою вона бути не може по визначенню. Цей тип – велике розчарування вченого. Бо на початку досліджень І.П.Павлов захоплювався цім типом ВНД, називав собак з таким типом „нервовими багатирями” і збирався саме їх призначити на роль сангвініків.
25.Якщо у людини сильні і збудження, і гальмування, але не піддається переробці диференціювання, то це свідчить про низьку рухомість нервових процесів. Отже, це за Павловим – спокійний тип /сильний, врівноважений, інертний/, що корелює з флегматиком за Гіппократом.
26.Якщо вироблений напередодні рефлекс не виявляється, це вказує на те, що тимчасовий зв’язок не був закріплений у ЦНС на достатньо глибокому рівні. В такому випадку тимчасовий зв’язок носив лише динамічний характер – існував у вигляді циркуляції збудження по ланцюгу нейронів, що ситуативно поєдналися. У основі такого поєднання лежить полегшення проведення збудження в структурах мозку, синхронність і син-фазність активності втягнутих в процес його утворення нейронів. Як наслідок, збільшується ефективність синаптичної передачі і виникає просторово-часова організація між-нейронних відносин. Але такий тимчасовий зв’язок схожий на оперативну пам’ять і довго у ЦНС не підтримується. Певну роль у цьому відіграє вид умовного рефлексу (УР) за співвідношенням подразників. Особливо швидко зникає тимчасовий зв’язок при занадто короткочасному виробленні штучних УР. Ці дві ознаки – 1)відсутність близькості між умовним і безумовним подразником (штучний рефлекс) і 2)лише динамічний характер тимчасового зв’язку, оскільки закріплення його на більш глибокому рівні не відбувається за відсутністю поєднань умовного і безумовного подразників подалі (тобто, випадковість такого поєднання) – дозволяють відносити такий рефлекс до особливої категорії – сумаційного рефлексу.