

Бінокулярна координація очних рухів
Око приводять до руху 6 зовнішніх м’язів. Виділяють такі рухові програми:
1)Співдружні рухи очей – вверх, донизу, праворуч, ліворуч - очи рухаються однаково.
2)Вергентні рухи – рухи одного ока віддзеркалюють рухи іншого: при наближенні зорового подразника – конвергентні; при віддаленні – дивергентні, при розгляді об’єкту на великій відстані – дивергенція виражена до такого ступеню, що вісі очей стають паралельними.
3)Циклоторсионні рухи – доступні спостереженню при нахилі голови у бік – обидва ока рухаються в одному напрямі в фронто-паралельній площині. Значні конвергентні рухи супроводжуються симетричними циклоторсионними (до 10E).
4)Саккади – швидкі скачки очних яблук між точками їх послідовної фіксації (тривалість 10-80 мсек). При довільних переходах зору амплітуда саккад може досягати більш 90E, і великі кути зміщення (наприклад, від 60E ліворуч до 40E праворуч) під час саккад завжди супроводжуються поворотами голови (якщо вона штучно не фіксована). Оскільки голова більше важить, то при одночасній імпульсації у нервах, що запускають рухи голови і очей, голова відстає від очей. Коли ж вона починає свій рух, очі, відносно голови рухаються назад, що підтримує сталим напрямок зору. Кутова швидкість рухів очей під час саккад пропорційна їх амплітуді, й досягає при великих саккадах (більш 60E) приблизно 500E/с.
Мікросаккади – амплітуда саккад складає лише декілька кутових хвилин.
Саккади можуть розділятися періодами фіксації тривалістю 0,2-0,6 с, якщо людина оглядає навколишню обстановку з чіткою структурою. Вони можуть на декілька секунд подолатися довільно (особливо після тренування). Але все одно точка фіксації зору у періоді між саккадами буде декілька зміщатися за рахунок мікросаккад та тремору ока.
85)Дрейф – тремор ока – повільне тремтіння ока, таке ж за амплітудою, як і мікросаккади. Частота дрібних рухів (мікросаккад і тремору) – 20-150 Гц
*При стеженні очні рухи можуть бути плавні, якщо об’єкт фіксований, а голова і тіло того, хто спостерігає – рухаються, а також при слідкуючих рухах очей, що супроводжують рухові об’єкти. Якщо швидкість рухового об’єкту, за яким стежить людина більш 80E/с, око відстає і слідкуючи рухи супроводжуються корекційними саккадами та поворотами голови.
*Точки фіксації зору при розгляді добре структурованого візуально об’єкту розташовані переважно поздовж контурів, їх розривів та пересічень. На частоту фіксації зору впливає інтерес людини до конкретної візуальної структури (наприклад, зір частіше фіксує очі і рот /при розгляді обличчя/ і вдвічі частіше прямує на праву половину обличчя). Тобто очно-моторне керування зазначається не тільки формальними особливостями візуального об’єкту, але й особистісним значенням зорових сигналів для людини.
*Слідкуючи рухи можуть викликати звукові і тактильні подразнення у темряві, але вони не мають зворотного зв’язку, тому менш точні і частіше перериваються саккадами.
Онтокінетичний ністагм (ОКН). Періодичне чергування повільних слідкуючих рухів і саккад називають ністагмом. Коли очі стежать за рухомим об’єктом (або псевдо-рухомим, наприклад, у потязі), обидва ока плавно співдружно зміщаються у напрямі рухомого подразника (повільна фаза ОКН, яка має більшу кутову швидкість рухів очей при активному стеженні, ніж при пасивному). Коли об’єкт зникає з поля зору, саккада у зворотному напрямку переводить зір на нову точку фіксації.
Зорове сприймання щільно зв’язане з рухами очей і голови, що змінюють напрямок погляду. Для забезпечення цих рухів існують численні поєднання між корковими зоровими ділянками і очно-руховими центрами у стовбурі мозку. Так сигнали з V2 йдуть у верхні горбики 4-горбія і керують нейронами ПМРФ і РФСМ, які контролюють напрямок погляду. Екстрастріарні зорові зони зв’язані з префронтальними корковими зонами, які відповідають за довільні рухи очей і координацію рухів кисті і роту. Ці зони, подібно відповідним зонам тім’яної кори (поле 7), поєднані з очно-руховими центрами стовбуру, а, за рахунок зворотних зв’язків, - з потиличною зоровою корою. Шлях із сітківки через нейрони верхніх горбиків важливий для передачі зорової інформації про рух у тім’яну зону (нейрони верхніх горбиків надсилають цю інформацію у подушку, звідки вона прямує до ектрастріарних коркових зорових зон і тім’яної кори /поле 7/). Вказані зв’язки сприяють інтеграції рухів погляду і зорового сприймання.
Фотореакції
-
З
ОВНІШНІЙ СЕГМЕНТЗорові пігменти
→подвійний ліпідний шар мембранних дисків/палички/) або
складок /колбочки/ ↓
(близько 1000)
↓
РЕСНІЧКА
↓
ОСНОВНА ЧАСТИНА КЛІТИНИ
|
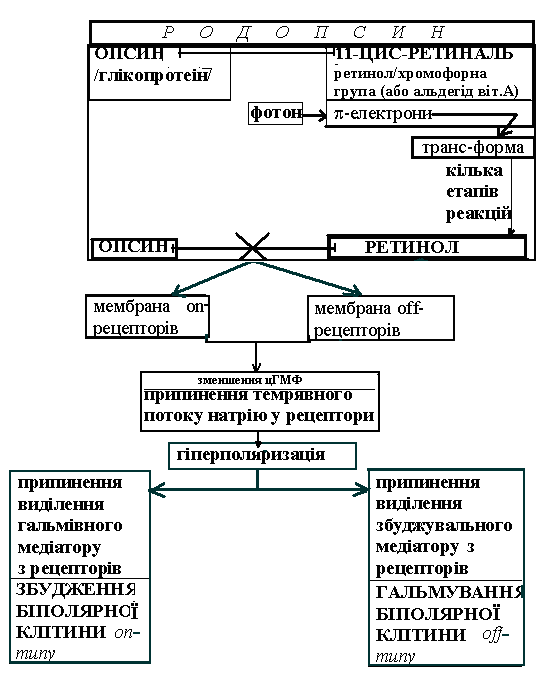
Палички і колбочки мають схожість у будові: обидва типи рецепторів включають у будову клітини зовнішній сегмент, що з’єднується з останньою частиною клітини вузькою реснічкою й складається приблизно з 1000 мембранних дисків (у паличок) і складок (у колбочок). Мембранні утворення (диски, складки) мають подвійний ліпідний шар, в який упорядковано включені молекули зорових пігментів.
Зоровий пігмент паличок – родопсин – скдадається з опсину (глікопротеїну) й альдегіду вітаміну А (11-цис-ретиналь – або ретинол / хромофорна група). Зоровий пігмент колбочок – йодопсин – має інший, ніж у паличок спектр поглинання світла й за типами колбочок поділяється на три типи (зокрема, хлоролаб, еритролаб).
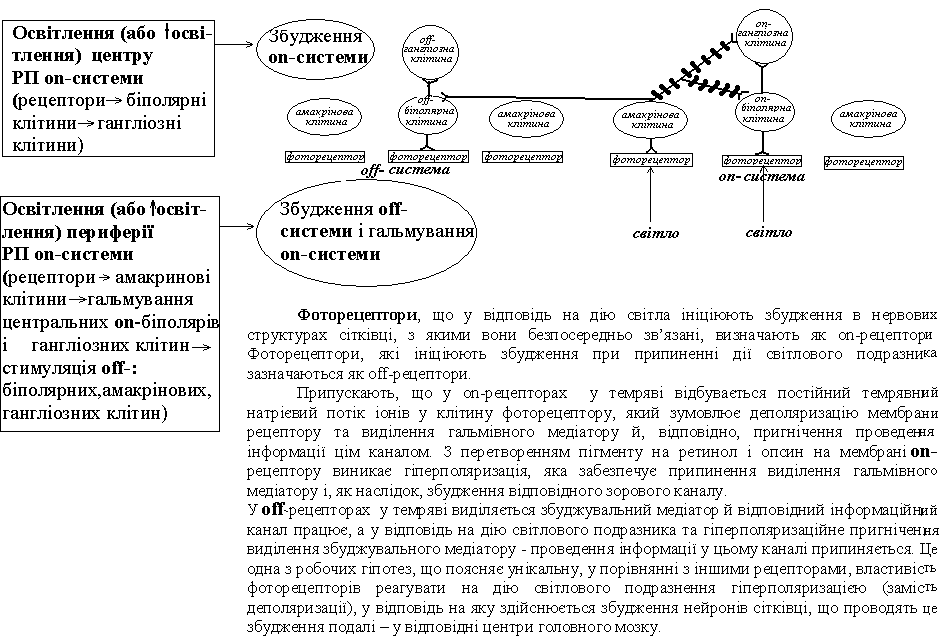
Процес трансдукції у фоторецепторі починається з поглинання фотону світла хромофорною групою пігменту (-електронами його спряжених подвійних зв’язків). Це зумовлює стереоефект – ізомеризацію ретиналю – його перехід з форм 11-цис у транс-форму. Після цього в кілька етапів вся молекула пігменту розривається на 2 частини – ретинол і опсин (мета-родопсин ІІ). Це обумовлює активацію G-протеїну(трансдуцину) → фосфоліпази → ↓концентрації цГМФ(за рахунок перетворення її в 5-ГМФ) → закриття натрієвих каналів і гіперполяризації фоторецепторів → блокування виходу гальмівного (on-система) або збуджувального (off-система) медіаторів
1.Трьохкомпонентна теорія (Юнг, Максвелл, Гельмгольц) постулює, що у сітківці існують три незалежні рецепторні системи, які представлені 3-ма типами колбочок, кожен з яких здатний реагувати або на червоний, або на зелений, або на синій колір. Прямі виміри спектрів поглинання фотопігментів одиноких колбочок, реєстрація їх рецепторних потенціалів та колір-сприймання на нижній межі фотопічної чутливості, коли розрізняються лише червоні, зелені і сині тони, об’єктивно підтвердили цю теорію. Комбінації сигналів, які отримаються від 3-х типів колбочок, аналізуються у 2-х нейронних системах – системах сприймання яскравості і кольору.
2.Теорія опонентних кольорів (Мах, Геринг). Термін „опонентні кольори” оснований на полярній природі сприймання кольору (наявності додаткових тонів). Згідно з цією теорією всі хроматичні валентності обумовлені 4-ма первинними кольорами – червоним, жовтим, зеленим, синім. Вони зв’язані попарно 2-ма фізіологічними процесами, що здійснюють на сприйняття кожного з кольорів пари антагоністичний вплив. Таким чином, виділені 2 кольорових опонентних процеси – для сприймання зеленого-красного, жовтого-синього; та 3-й опонентний процес для ахроматичних валентностей – білого та чорного. Це означає, що не може бути зеленувато-червоного або синювато-жовтого кольорів. За первинними поглядами у рамках цієї концепції, під впливом променів з певною довжиною хвилі виникає розпад пігментів у певних колбочках, що призводить до відчуття білого, червоного та жовтого; а під впливом інших світлових хвиль пігменти, навпаки, синтезуються, що супроводжується виникненням відчуттів чорного, зеленого і синього. Пізніше було доведено, що подібний механізм діє не на рівні фоторецепторів, а на рівні нейронів сітківці та вище. Ця теорія підтверджується ефектами одночасного і послідовного хроматичного контрасту: якщо сіре коло оточується ярко-зеленим кільцем, то, за рахунок одночасного хроматичного (кольорового) контрасту, воно набуває червонуватий відтінок; якщо кільце прибрати, то на його місці залишається уявне червоне кільце, яке оточує зелене коло (послідовний хроматичний контраст). Завдяки теорії опонентних кольорів виявлені антагоністичні колір-специфічні нейронні механізми, що діють у різних хребтових на різних рівнях. Такі механізми можуть працювати вже на рівні сітківки. Так, спектральні відповіді горизонтальних і біполярних клітин обумовлені просторовою взаємодією сигналів від колбочок різного типу. У риби, наприклад, визначається 3 класи горизонтальних клітин: 1)клітини, що гіперполяризуються незалежно від довжини світлової хвилі 2,3)клітини, які гіперполяризуються при стимуляції монохроматичним світлом однієї частини спектру і деполяризуються під впливом світла з іншої його частини і демонструють при цьому червоно-зелений і синьо-жовтий антагонізм. У ссавців з кольоровим зором аналогічна організація перенесена на деякі: біполярні клітини (з on- і off-центрами), гангліозні клітини і нейрони ЛКТ. У сітківці, зокрема, виявлені нейрони, що, наприклад, активуються червоним і гальмуються зеленим світлом в їх РП (прості опонентні клітини). Крім того принцип опонентності проявляється у деяких нейронів у організації їх РП – центр і периферія РП дають протилежні відповіді на різні кольори: наприклад, якщо червоний у центрі їх збуджує, а зелений гальмує, то на периферії РП відповіді протилежні (двічі опонентні клітини). Така подвійна опонентна організація поясняє феномен одночасного кольорового контрасту. Нейрони такого типу зазначені у ЛКТ і первинній зоровій корі приматів з кольоровим зором.
Оброблення колір-специфічної інформації у зоровій корковій первинній зоні V1 здійснюється, головним чином, там, де нема нейронів з орієнтаційно-специфічними РП. Серед порушень кольорового зору найбільш типовими являються двобічні (на обох очах) периферійні порушення, а ті, що зв’язані з враженням великих півкуль, - виникають дуже рідко.
Розрізняють 2 головних типи периферійних порушень колір-сприймання – трихроматичний і дихроматичний.
Аномальні трихромати розрізняють менше кольорових валентностей, ніж нормальні люди. Виділяють 3 категорії аномалій колір сприйняття: протаномали (червоно-сліпі) і дейтераномали (зелено-сліпі) не здатні розрізняти ненасичені червоні і зелені кольори – протаномали при дослідженні на аномалоскопі додають у суміш більш, ніж у нормі, червоного, а дейтераномали – зеленого; тританомали (рідка патологія) мають порушення жовто-синьої системи.
Дихроматопсія. Дихромати можуть підбирати усі тона, які бачуть, за допомогою лише 2-х спектральних кольорів. Тому число кольорів, що вони розрізняють значно менше, ніж у трихроматів (протанопи плутають деякі червоні тони з чорними, темно-сірими, коричневими, а інколи, подібно дейтеранопам, з зеленими; для них частина сонячного спектру у межах 480-490 нм виглядає ахроматичною, а для дейтеранопів ахроматичний діапазон 495-500 нм; максимум кривої фототопічної спектральної яскравості у протанопів – 520, у дейтеранопів – 580 нм; у тританопів /що також рідко зустрічаються/ сине-фіолетова частина спектру здається переходом від сірого до чорного, ахроматичною для них являється ділянка 565-575, вони плутають жовті та сині кольори).
Звичайно у дихроматів і трихроматів аномалія спадкується як рецесивна ознака, що зв’язана з Х-хромосомою (тому частіше страждають на таку патологію чоловіки); скотопічний зір (паличковий, нічний зір, у протилежність фотопічному зору – колбочковому, денному) в них, як правило, не порушений.
Повна кольорова сліпота (дальтонізм) зустрічається у менш 0,01% всіх людей. Монохромати бачать світ чорно-білим, погано розрізняють форму при денному освітленні, легко осліпляються й страждають на фотофобію. В них знижена гострота зору у центральній ямці. Вважають, що в їх колбочках у якості зорового пігменту виступає родопсин.
Порушення паличкової системи супроводжується нормальним сприйманням кольорів і обмеженою здатністю до темрявної адаптації – „куряча сліпота” (нікталопія) /причиною може бути дефіцит у їжі вітаміну А1 – попереднику ретиналю/.
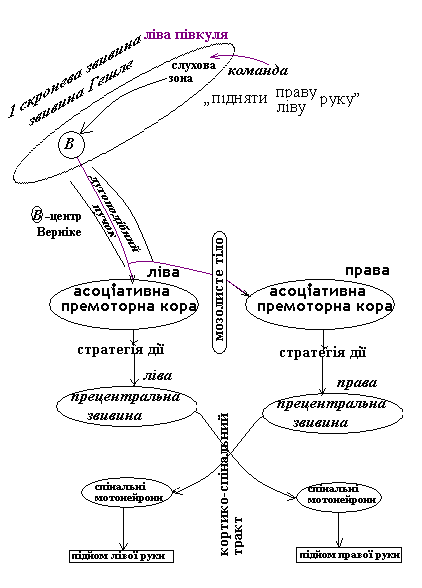
Схема передачі інформації при виконанні команди
« підняти праву чи ліву руку»


Фізичне навантаження
Ресинтез АТФ:
1)креатин-фосфатний шлях (КФ – креатин-фосфат, якого у м’язах значно більш, ніж АТФ) – ферментативний перенос фосфатної групи від КФ до АДФ (для цього потрібні мс і відновлюється АТФ близько 3,6 ммоль/хв, але запасів КФ досить лише на 1-2 с роботи; 8-10 с при „марафонських” навантаженнях)
2)гліколітичний шлях/анаеробний гліколіз (триває близько хвилини з утворенням 1,2 ммоль/хв АТФ; 90-100 с при „марафонських” навантаженнях) – з утворенням недоокислених продуктів (піровиноградної та молочної кислот)
3)аеробне окислення глікогену та ліпідів (у формі жирних кислот та ацетооцтової кислоти; чим довше фізична робота, тим більша частина енергії звільняється за рахунок окислення ліпідів) – найбільш економний і інерційний шлях (активність необхідних ферментів підвищується через 2-4 хв після початку скорочення м’язу; для цього потрібен кисень, доставка якого залежить від систем дихання і кровообігу, що впрацьовуються порівняно повільно)
Рівні розвитку втоми
1)Метаболічний (кінцевий за виникненням): а)накопичення фосфорної, молочної кислот, СО2, і др. пригнічують працездатність м’язу; частина цих продуктів разом з К+ дифундують з м’язів назовні і пригнічують здатність м’язової мембрани до генерації ПД (теорія засмічення) б)поступово витрачаються запаси глікогену (необхідного для ресинтезу АТФ і КФ), ліпідів, й, відповідно, самих АТФ і КФ (теорія виснаження) і падає напруга кисню у органі (гіпоксія), що також гальмує ресинтез АТФ і КФ (теорія задушення). Зменшення запасів глікогену в м’язах і печінці супроводжується зменшенням рівня глюкози у крові, яка є основним продуктом окислення у нервових клітинах. Це призводе до зменшення збудливості у нервових центрах і зростання активності гальмівних медіаторів (зокрема, ГАМК). На такому фоні розвивається порушення регуляції рухових функцій, розлад вегетативної регуляції і дисфункція вісцеральних систем.
2)Синаптичний на нейро-органному рівні (проміжний рівень): в основі – виснаження нейро-органних медіаторів, зв’язане з частою стимуляцією синапсів, і зменшення збудливості постсинаптичної мембрани за тою ж причиною.
3)Центральний (первинний рівень): процес гальмування у ЦНС, який: припиняє діяльність раніше, ніж розвивається глибоке виснаження на рівні працюючих клітин, і стимулює розвиток відновних процесів
На підставі цього механізму І.М.Сеченовим визначений феномен активного відпочинку – збільшення ефективності відновлення працездатності при включенні в роботу під час відпочинку інших нервових центрів (тобто, замість пасивного відпочинку – інша діяльність). Це викликає такий перебіг процесів збудження і гальмування у ЦНС, який розвантажує втомлені нервові центри і оптимізує всю регуляцію у цілому. До того ж, інша робота невеликої інтенсивності сприятиме посиленню дихання і поліпшенню кровообігу і виведення продуктів обміну з м’язів, які працюють. Відбуватиметься посилення процесів окислення. Зокрема активно окислятиметься молочна кислота (один з органів, де вона поглинається, є серце). Крім того, окислення молочної і піровиноградної кислот, що утоворюються у м’язі під час скорочення, супроводжується фосфориліруванням АДФ і КФ, тобто ресинтезом КФ і АТФ.
Правила Фольборта
1.Рівень працездатності залежить від співвідношення процесів виснаження (комплексу біохімічних змін, які супроводжуються зменшенням енергетичного потенціалу і відбуваються під час втоми) і відновлення (процесів, спрямованих на збільшення енергетичного потенціалу). Ці процеси розвиваються одночасно при перевазі одного з них. Перевага виснаження → ↓працездатності; перевага відновлення → ↑працездатності.
2.Співвідношення між виснаженням і відновленням під час праці і після неї змінюється: під час праці переважають процеси виснаження, хоча відбуваються і процеси відновлення; після праці домінують процеси відновлення.
3.Відновлення стимулюється змінами, що відбуваються в процесі виснаження. Таким чином, без попереднього розвитку втоми немає відновлення і підвищення працездатності.
4.Швидкість розвитку виснаження визначає інтенсивність відновних процесів: що інтенсивніше робота і швидше виникає виснаження, то й швидше відновлення після роботи.
5.Відновлення має хвилеподібний характер розвитку при загальній тенденції до збільшення працездатності. Розрізняють 2 фази процесу відновлення: 1)досягнення початкової працездатності 2)стійкої працездатності
6.Повернення до праці до того, як досягається стан стійкої працездатності призводить до поглиблення процесів виснаження і розвитку хронічного виснаження (відовлення розвивається повільно, протягом тижнів і навіть місяців). Повернення до праці під час розвинутого стану стійкої працездатності призводить підвищення працездатності і розвитку тренування (робочої гіпертрофії м’язів, збільшення енергетичного їх резерву, активації аеробного і анаеробного метаболізму; вдосконалення нейро-гуморальної регуляції вісцеральних систем, збільшення їх резервних можливостей). Таким чином, зміни працездатності залежать від співвідношення тривалості праці і відпочинку.
7. На розвиток втоми і послідовного відновлення впливає гальмування в нервових центрах. Процес гальмування у ЦНС припиняє діяльність раніше, ніж розвивається глибоке виснаження на рівні працюючих клітин, і стимулює розвиток відновних процесів. Відновні процеси прискорюються під час активного (замість пасивного) відпочинку (І.М.Сеченов)
Модуль 1
1.Блокада кальцієвих каналів буде запобігати виходу іонів кальцію з СПР у цитоплазму м’язового волокну. Отже, іони кальцію не зможуть здійснити конформаційну активацію тропоніну, тропонін не набуде здатності впливати на тропоміозин, а тропоміозин не звільнить активний центр на актині. Таким чином, головка міозину не зможе вставитися в активний центр на актині, бо він буде блокованим і актино-міозинові містки не створяться (до того ж, без контакту з актином не будуть активуватися ферментативні АТФ-азні властивості міозину).
2.Спринтерам бажано мати перевагу білих м’язових волокон. Їх характеризує порівняно великий розмір, висока збудливість, високий вміст глікогену і низький – мітохондрій, міоглобіну. Кровопостачання у таких волокнах незначне і, відповідно, переважає анаеробний тип метаболізму. Такі волокна швидко скорочуються і швидко втомлюються.
3.Стадії, що спостерігаються під час фізичної роботи, такі: 1)передстартового стану 2)впрацьовування 3)стійкої працездатності 4)стомлення (втоми) 5)відновлення. Але ергографія відбиває лише 2-4 стадії.
У випадку, коли на ергографі виявилося зменшення амплітуди піднімання вантажу на 50%, має місце 4 стадія – стомлення. При міографії його головні ознаки – це зменшення амплітуди поодиноких скорочень, збільшення тривалості скорочень за рахунок подовження латентного періоду і періоду розслаблення.
Механізми, що лежать у основі стомлення, різні. Розрізняють центральний первинний рівень стомлення: це процес гальмування у ЦНС, який: припиняє діяльність раніше, ніж розвивається глибоке виснаження на рівні працюючих клітин, і стимулює розвиток відновних процесів. На другому місці за швидкістю виникнення стомлення – нейро-органний синаптичний рівень, в основі котрого – виснаження нейро-органних медіаторів, зв’язане з частою стимуляцією синапсів, і зменшення збудливості постсинаптичної мембрани за тою ж причиною. Кінцевий за швидкістю виникнення метаболічний рівень: а)накопичення фосфорної, молочної кислот, СО2, і др. пригнічують працездатність м’язу; частина цих продуктів разом з К+ дифундують з м’язів назовні і пригнічують здатність м’язової мембрани до генерації ПД (теорія засмічення) б)поступово витрачаються запаси глікогену (необхідного для ресинтезу АТФ і КФ), ліпідів, й, відповідно, самих АТФ і КФ (теорія виснаження) і падає напруга кисню у органі (гіпоксія), що також гальмує ресинтез АТФ і КФ (теорія задушення). Зменшення запасів глікогену в м’язах і печінці супроводжується зменшенням рівня глюкози у крові, яка є основним продуктом окислення у нервових клітинах. Це призводе до зменшення збудливості у нервових центрах і зростання активності гальмівних медіаторів (зокрема, ГАМК). На такому фоні розвивається порушення регуляції рухових функцій, розлад вегетативної регуляції і дисфункція вісцеральних систем.
В даному випадку при порівняно нетривалій, за часом, ергографії втома імовірно зв’язана з центральним гальмуванням працюючих м’язів.