

Зоровий тракт веде до:
1)Латеральних колінчатих тіл (лкт), які у функціональному відношенні належать до таламусу
ЛКТ – остання інстанція, яка переробляє зорову інформацію перед тім, як відправити її до кори. Іпсилатеральні і контр-латеральні волокна зорового шляху закінчуються у різних ділянках ЛКТ. Аксони переважної кількості нейронів ЛКТ проходять через зорову променистість до нейронів первинної зорової кори (V1 поле /17/) у потиличній зоні. Первинна зорова кора поєднується з вторинною (поле V2), третинною (поля V3, V3а, V4) зоровою корою та інтегративними зонами потилично-тім’яної і потилично-скроневої кори головного мозку.
Проекцією зорового шляху у ЛКТ забезпечуються процеси: а)впізнання об’єктів б)кольорового зору в)сприймання руху г)стереоскопічне сприймання глибини.
Більша кількість нейронів ЛКТ має концентричні рецептивні поля (РП), які організовані подібно РП гангліозних клітин і подібно цим клітинам класифікуються за участю у реакції на включення та вимкнення зорового стимулу.
Частина нейронів ЛКТ забезпечує механізм посилення одночасного контрасту („контрастові нейрони”), інші нейрони чутливі до руху зорових стимулів. Зазначають також нейрони ЛКТ, які сприймають локальні значення світла і тіні зорового стимулу („нейрони темного-світлого”), нейрони, що відрізняються колір-специфічними РП.
Нейрони ЛКТ отримують інформацію не тільки від зорового шляху, але й від стовбуру мозку. Вважають, що ці незорові зв’язки здійснюють вплив на обробку зорової інформації у залежності від ступеню бадьорості, просторової спрямованості уваги і зв’язаних з нею рухів очей.
Коллатеральні волокна, що відходять від основного шляху, закінчуються у:
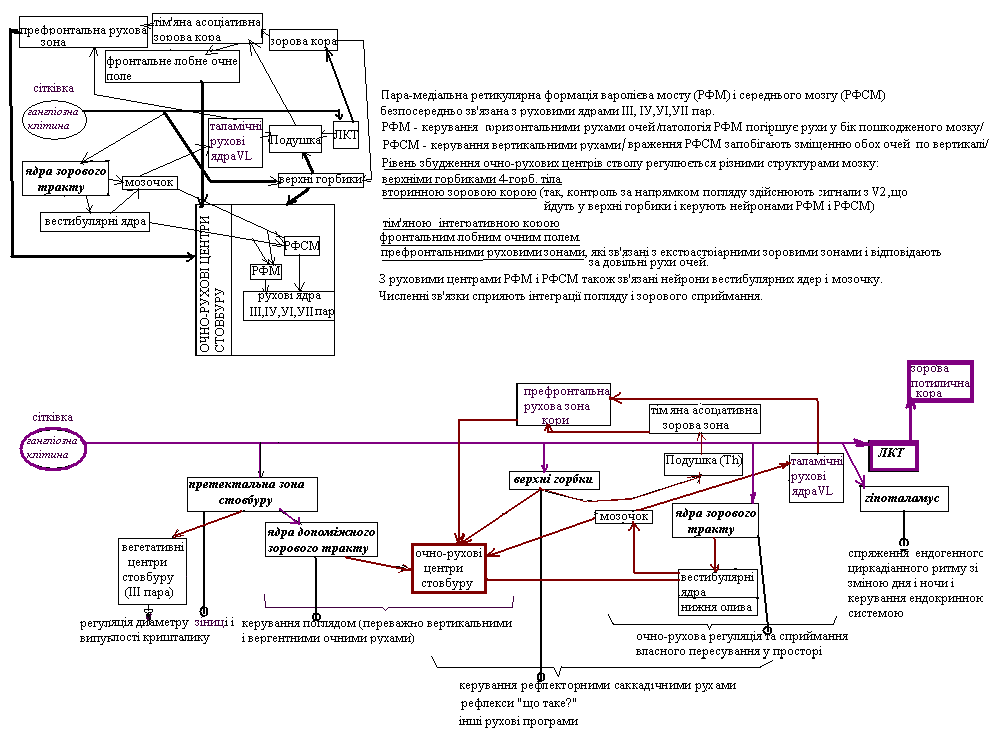
2)верхніх горбиків 4-горбія (ця проекція служить для керування рефлекторними саккадічними рухами очей: аксони нейронів верхніх горбиків йдуть у очно-рухові центри стовбуру мозку та зорову частину подушки /асоціативне ядро таламусу/, через яку зорові сигнали досягають тім’яних зорових асоціативних зон)
3)ядрах зорового тракту (нейрони цього тракту отримують інформацію від специфічно реагуючих на рух on-off-нейронів /гангліозних клітин сітківки/ і поєднують їх з вестибулярними ядрами стволу мозку та нижньою оливою, звідки інформація прямує до мозочку; ці проекції використовуються для очно-рухової регуляції та сприймання власного пересування у просторі)
4)ядрах допоміжного зорового тракту у претектальній зоні (що поєднуються з центрами стовбуру мозку, які керують поглядом – головним чином, вертикальними і вергентними рухами очей)
5)ядрах претектальної зони стовбуру мозку (частина таких зв’язків служить для регуляції діаметру зіниці)
6)ядрах гіпоталамусу (ці зв’язки служать для спряження ендогенного циркадіанного ритму зі зміною дня та ночі, а також для керування ендокринною системою, у тому числі, для здійснення, через епіфіз, впливів сітківки на пігментацію шкіри)
Проекція основного Шляху |
Функція |
Завершення проекції |
|
до верхніх горбиків 4-горбія |
Керування рефлекторними саккадічними рухами очей орієнтувальні зорові рефлекси інші рухові прогами |
у очно-рухові центри стовбуру мозку та зорову частину подушки→ тім’яні зорові асоціативні зони |
|
до ядер зорового тракту від специфічно реагуючих на рух on-off-гангліозних клітин сітківки |
очно-рухова регуляція і сприймання власного пересування у просторі |
до вестибулярних ядер стволу мозку та нижньої оливи→мозочок |
|
до ядер допоміжного зорового тракту у претектальній зоні |
керування поглядом (головним чином, вертикальними і вергентними рухами очей) |
поєднуються з центрами стовбуру мозку |
|
до претектальної зони стовбуру мозку |
зокрема, регуляція діаметру зіниці |
|
|
до гіпоталамусу |
спряження ендогенного циркадіанного ритму зі зміною дня та ночі, керування ендокринною системою(зокрема,здійснення, /через епіфіз/ впливів сітківки на пігментацію шкіри |
|
|
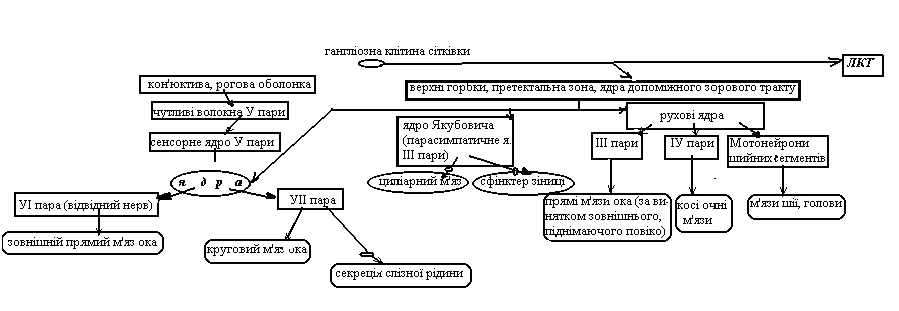
М’язи ока
круговий → рухові волокна УІІ пари парасимпатичні волокна ← секреція слізної рідини
зовнішній прямий → -„-„-„-„-„-„-„- УІ пари
косі → -„-„-„-„-„-„-„- ІУ пари
прямі → -„-„-„-„-„-„-„- ІІІ пари парасимпатичні волокна ← циліарний; сфінктер зіниці
(за винятком зовнішнього, піднімаючого повіка)
3-хмірне сприймання об’єктів за допомогою бінокулярної системи особливо важливе, коли вони розташовані у зоні безпосереднього досягнення або поруч з нею. Оскільки очі містяться у різних частинах голови, зображення на їх сітківках декілька різні, що слідує з законів геометричної оптики. Чим крупніше предмети і чим ближче вони до спостерігачу, тим більше таке горизонтальне розходження – диспаратність. Це можна проілюструвати таким експериментом: якщо витягнути руку перед собою і розглядати великий палець то правим, то лівим оком, то складається враження, що палець стрибком зміщується відносно фону – коли на палець дивиться праве око, він затуляє одні предмети, а коли ліве – інші. Чим більше відстань об’єктів заднього плану від руки, тим більше зсування, відносно них, пальця. Коли обидва ока фіксують точку на певній відстані, всі об’єкти на колі, що йде через вузлові точки оптичних систем двох очей і точку фіксації, проектуються на кореспондуючі ділянки сітківок. Всі об’єкти у середині або за межами цього кола (гороптерного кола) проектуються на не кореспондуючі ділянки сітківок. Якщо горизонтальна диспарантність (як сума певних кутів) достатньо велика, зображення подвоюється. Просторовий розподіл 2-х зображень при горизонтальній диспарантності можна вимірити при використанні моделі „циклопічного ока”.
Таким чином, горизонтальна диспарантність виступає як кількісна міра різниці між місцями на сітківках, на які проектується зображення об’єкту, який розташований на кінцевій відстані від очей при звичайному положенні голови.
Горизонтальна диспарантність і бінокулярне злиття. Коли диспарантність перевищує певну величину бінокулярне спостереження 3-хмірного предмету призводить до сприймання його як подвійного. Діапазон відстаней до об’єкту – від нульової диспарантності (об’єкт у безмежності)і кінцевої межі бінокулярного глибинного зору, при якій сприймання стає подвійним, досить великий. Це поясняється на підставі величини РП бінокулярно активованих коркових нейронів: розмір ЗРП таких нейронів у ділянках V1і V2 значно більше, ніж просторовий еквівалент відповідної ділянки сітківки. Ті з коркових нейронів, в яких РП на обох сітківках точно співпадають, максимально збуджуються об’єктами у межах гороптерного кола. Ті з бінокулярно активованих нейронів кори, в яких РП не сітківках точно не співпадають, максимально збуджуються об’єктом, який міститься на певній відстані від гороптерного кола, що призводить до певної горизонтальної диспарантності.
Стереоскопічний зір перевіряється за допомогою бінокулярного погляду на 2 рисунки, що розташовані поруч і виглядають для кожного ока відокремлено як випадкові набори точок. Ті, хто здатний бінокулярно корелювати окремі точки з відповідною горизонтальною диспарантностю, здатні сприймати у даному випадку й форму і глибину зображень.