

У эукариот свой набор днк полимераз
Эукариоты отличаются от прокариот и по набору ДНК полимераз. Клетки млекопитающих содержат четыре различных ДНКполимеразы, а дрожжевые клетки, по крайней мере, пять –и . Краткая характеристика свойств каждого фермента выглядит следующим образом:
- Характеризуется праймазной активностью, высокочувствительна к афидиколину –ингибитору полимеразной активности. Функционирует на отстающей цепи..
- имеет низкую процессивность. Участвует в процессе репарации ДНК. В отличие от обладает низкой чувствительностью к афидиколину.
- митохондриальная ДНКполимераза. Характеризуется низкой чувствительностью к афидиколину.
- основная полимераза лидирующей цепи. Обладает высокой процессивностью, которая поддерживается специальным белком, называемым антигеном ядер пролиферирующих клеток (PCNA) подобна холоферменту ДНКполимеразы III E. coli.
- функция этой полимеразы еще не полностью изучена.
Теломеры – «молекулярные часы клетки»
Особенностью репликации линейных молекул ДНК эукариот является сложность репликации концевых отделов материнских цепей. Отстающая цепь репликативной вилки в синтезе ДНК не может синтезироваться до 5'-конца в отсутствие праймера, который, в свою очередь, не образуется непосредственно на концевом фрагменте. Потери концевой ДНК делают невозможной бесконечную пролиферацию. Принято считать, что эта особенность лежит в основе «молекулярных часов», определяющих продолжительность жизни соматических клеток Хромосомы позвоночных оканчиваются последовательностью ТТАGGG, которая повторяется сотни и тысячи раз. Эти повторяющиеся последовательности получили название теломеры. Существование специальных структур на концах хромосом было постулировано в 1938 году классиками генетики, лауреатами Нобелевской премии Барбарой Мак-Клинток и Германом Мёллером . Независимо друг от друга они обнаружили, что фрагментация хромосом (под действием рентгеновского облучения) и появление у них дополнительных концов ведут к хромосомным перестройкам и деградации хромосом . В сохранности оставались лишь области хромосом, прилегающие к их естественным концам . Лишенные концевых теломер, хромосомы начинают сливаться с большой частотой, что ведет к тяжелым генетическим аномалиям . Следовательно, заключили они, естественные концы линейных хромосом защищены специальными структурами . Г . Мёллер предложил называть их теломерами (от греч. телос - конец и мерос – часть). В последующие годы выяснилось, что теломеры не только предотвращают деградацию и слияние хромосом (и тем самым поддерживают целостность генома хозяйской клетки), но и, по-видимому, ответственны за прикрепление хромосом к специальной внутриядерной структуре [своеобразному скелету клеточного ядра], называемой ядерным матриксом. Таким образом, теломеры играют важную роль в создании специфической архитектуры и внутренней упорядоченности клеточного ядра. Наличие на концах хромосом специальной теломерной ДНК позволяет решить и так называемую проблему концевой недорепликации ДНК .
Соматические клетки теряют от 50 до 200 нуклеотидов при каждом клеточном делении, что и является основой молекулярного механизма, определяющего продолжительность жизни этих клеток. В противоположность соматическим смертным клеткам, то есть клеткам, обладающим пределом размножения in vitro, большинство «бессмертных» клеток, обладающих способностью к бесконечной пролиферации, содержит специальный фермент теломеразу. Теломераза - это фермент, достраивающий свободные 3'- концы хромосом короткими повторяющимися последовательностями (в случае позвоночных - TTGGGG) . Теломераза содержит собственную РНК-матрицу (рис.8ю27 ), поэтому может быть отнесена к обратным транскриптазам.. Используя рибонуклеотид как матрицу, теломераза удлиняет молекулу ДНК, которая затем реплицируется с участием ДНК-полимеразы. Можно предположить, что ингибиторы обратных транскриптаз, такие как азидотимидин и карбовир, могут подавлять ее работу в клетке, вызывая процесс, похожий на старение клеток in vitro и включающий в себя постепенное прекращение пролиферации и специфическое изменение морфологии клеток
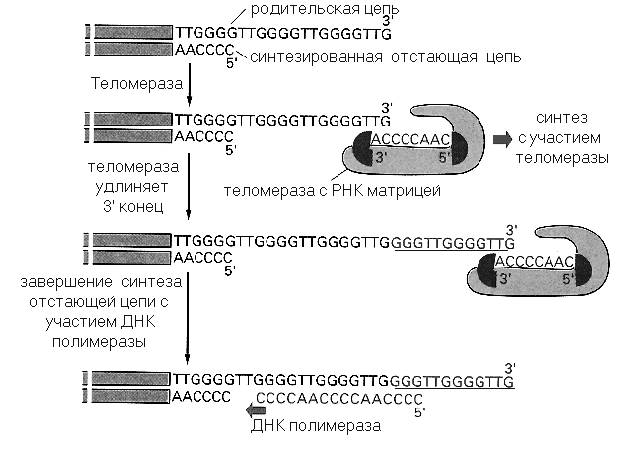
Рис.8.27.Синтез теломеров при участии теломеразы.