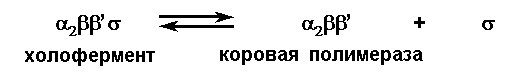Механизм синтеза рнк во многом напоминает синтез днк
Об участии в синтезе белков РНК известно с 40-х годов ХХ столетия, но роль и-РНК как молекулы посредника стала известна в 1961 году благодаря работам французских биохимиков Ф.Жакоба и Ж. Моно. В начале 60-х открывается и фермент, катализирующий синтез РНК – РНК-полимераза. В целом, механизм синтеза РНК по многим свойствам напоминает синтез ДНК. Для работы РНК-полимеразы необходимы:
Матрица. Роль матрицы, как правило, выполняет двуцепочечная ДНК.
Субстраты. Необходимы все четыре рибонуклеозид трифосфата АТФ, ГТФ, ЦТФ и УТФ.
Ионы двухвалентных металлов. Обычно это ионы Mg2+.
Схема синтеза РНК может выглядеть следующим образом:
аАТФ+bГТФ+cЦТФ+dУТФ РНК + (a+b+c+d) пирофосфат.
Синтез молекулы РНК идет в направлении 5’ 3’ с перемещением фермента по матрице в направлении 3’ 5’. Механизм соединения нуклеотидов под влиянием РНК-полимеразы подобен синтезу ДНК и заключается в нуклеофильной атаке внутреннего фосфата очередного рНТФ 3’-ОН группой на конце растущей цепи. Необратимость реакции связана с гидролизом пирофосфата, высвобождаемого в реакции синтеза РНК.
Однако существует и ряд особенностей в синтезе РНК. РНК-полимераза не нуждается в затравочном олигонуклеотиде (праймере), не обладает нуклеазной активностью, перемещается значительно медленнее по матрице (приблизительно 50-100 оснований/сек для РНК против около 1000 осн/сек для ДНК). Точек инициации транскрипция намного больше чем у репликации. Число молекул РНКполимераз в клетке намного больше, чем ДНК-полимераз. Наконец, точность полимеризации РНК - намного ниже, чем ДНК. Это допустимо, так как дефектные молекулы РНК могут быть просто удалены и взамен синтезированы новые «правильные» молекулы.
Транскриптон (оперон) - единица транскрипции.
Синтез молекул РНК начинается в определенных местах ДНК, называемых промоторами и завершается в терминаторах. Участок ДНК, ограниченный промотором и терминатором, представляет собой единицу транскрипции - транскриптон (оперон у прокариот). В пределах каждого транскриптона копируется только одна из двух нитей ДНК, которая называется значащей или матричной. Во всех транскриптонах, считываемых в одном направлении, значащей является одна нить ДНК, в транскриптонах, считываемых в противоположном направлении, значащей является другая нить ДНК.
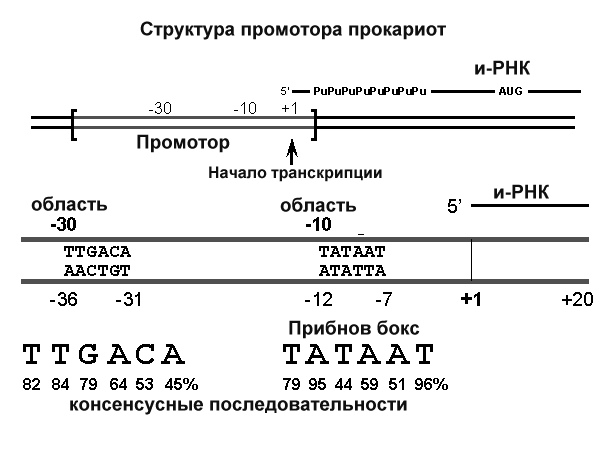
Рис.8.31. Структура промотора прокариот
Соседние транскриптоны могут быть отделены друг от друга нетранскрибируемыми участками ДНК, а могут и перекрываться, в частности так, что в пределах участка перекрывания матричными оказываются обе нити. Разбиение ДНК на множество транскриптонов обеспечивает возможность независимого считывания разных генов, их индивидуального включения и выключения. У эукариот в состав транскриптона, как правило, входит только один ген, у прокариот транскриптон может содержать несколько структурных генов.
Промоторы имеют сходное строение
Исследование структурных особенностей транскриптонов (оперонов) у прокариот позволило выявить ряд общих последовательностей у всех прокариот в областях начала транскрипции (промоторы) и терминации транскрипции. Согласование частот встречаемости оснований выявило 2 коротких консервативных участка последовательностей, расположенных на расстоянии 35 нуклеотидных пар в сторону 5’ конца от начала транскрипции (область -30 на рис.8.31) и на расстоянии 10 нуклеотидов в ту же сторону (область -10). АТ богатый участок области -10 получил название ТАТА бокса или Прибнов бокса. Учитывая более низкую температуру плавления такого участка, считается, что эта последовательность может быть легко денатурирована РНК-полимеразой и является удобным местом начала транскрипции. Обнаружены и консенсусные последовательности в областях окончания транскрипции. Копирование таких последовательностей позволяет формировать в этой области первичного транскрипта шпилечные структуры РНК, которые способствуют диссоциации РНК-полимеразы и окончанию транскрипции.
Транскриптоны эукариот устроены значительно сложнее. В промоторной зоне транскриптона выявляются зоны, которые указывают не только место начала транскрипции (как у прокариот), но частоту этого процесса.

Значительные различия коснулись регуляторной части транскриптона.. На расстоянии 27-30 пар нуклеотидов от сайта инициации расположен TATA-бокс, консенсусная последовательность которого выглядит как TATA(A/T)A(A/T) . Положение TATA-элемента строго определяет сайт инициации транскрипции , т.е. 5'-конец транскрипта . При повреждении или удалении TATA-элемента образуется набор молекул РНК с разными 5'-концами. Отдельные нуклеотидные замены в TATA-элементе могут приводить к резкому снижению эффективности транскрипции. Промоторная зона некоторых генов (например, гена гидроксиметилглутарилKoA-редуктазы - ключевого фермента биосинтеза холестерола у человека) не содержит TATA-элемента, и транскрипция начинается с нескольких разных сайтов. Образующиеся РНК различаются по 5'-концам в районе нетранслируемой лидерной последовательности. Возможно, различные лидерные зоны определяют характер регуляции экспрессии генов на уровне трансляции .
Расположенный дальше к 5’ концу участок регулирует базовую частоту транскрипции (ЦААТ бокс). Еще далее от начала сайта инициации располагаются последовательности, участвующие в изменении базового уровня транскрипции (энхансеры ускоряют, а сайленсеры замедляют этот уровень) и, наконец, имеется группа последовательностей, участвующих в регуляции экспрессии генов под действием различных внутри и внеклеточных регуляторов (гормонов, белков теплового шока, ионов металлов и т.д.). Указанные регуляторные последовательности представляют собой места связывания специфических бековых факторов.
Терминация транскрипции у эукариот - это пока довольно плохо изученный процесс. Известно лишь, что зона терминации расположена далеко за пределами 3’- конца кодирующей области. Эксперименты с разными искусственными конструкциями показывают, что для терминации транскрипции нужно существование в геноме зоны терминации и обязательно расположенного перед нею сайта полиаденилирования . Если последний убрать, то транскрипция идет за зону терминации. В то же время зону терминации можно удалять и приближать к сайту полиаденилирования, и терминация все равно будет происходить в ее пределах.
Нуклеотидные последовательности зон терминации, прилежащих к разным генам, содержат ряд гомологичных областей, однако в настоящий момент вопрос о природе сигналов терминации остается открытым.
РНК-полимераза у прокариот –основной фермент синтеза всех РНК
РНК-полимераза, выделеная у прокариот катализирует синтез всех типов РНК клетки. Это сложно устроенный фермент, состоящий из нескольких субъединиц, функции которых приведены в следующей таблице.
Таблица 8.10. Свойства и функции субъединиц РНК-полимеразы. |
||||
Субъединица |
Ген |
Размеры (кД) |
Число на холофермент |
Функция |
|
rpoB |
151 |
1 |
Полимеразная активность |
' |
rpoC |
155 |
1 |
Связывание с матрицей |
|
rpoA |
37 |
2 |
Структурный организатор полимеразы |
|
rpoD |
70 |
1 |
Узнавание промотора |
Коровый (основной) фермент состоит из 4-х субъединиц 2’, после соединения с - субъединицей превращается в холофермент, который и обеспечивает процесс инициации синтеза РНК.