

У эукариот молекула рнк модифицируется после транскрипции.
По окончании транскрипции бактериальных р-РНК и т-РНК, они могут быть сразу использованы в трансляции. Никаких дополнительных изменений в структуре этих молекул не происходит. Трансляция бактериальных иРНК может начинаться даже до окончания транскрипции. Это связано с отсутствием границ между ядром и цитоплазмой. Способность инициировать трансляцию прокариотических РНК, до завершения транскрипции, предоставляет уникальную возможность регуляции транскрипции некоторых генов. Еще одна особенность бактериальных иРНК – они полицистронны
. Это означает, что один транскрипт
является копией нескольких структурных
генов У эукариот иРНК –копия одного
гена.
Это означает, что один транскрипт
является копией нескольких структурных
генов У эукариот иРНК –копия одного
гена.
В отличие от прокариот все типы эукариотических РНК подвергаются значительной
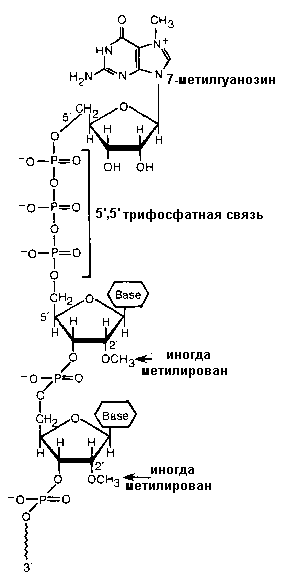
Рис 8.34. Строение «кэпа» и-РНК эукариот
посттранскрипционной модификации (процессингу). Вся совокупность ядерных транскриптов РНК-полимеразы II известна как гетерогенная ядерная РНК ( гяРНК ), поскольку одна из основных характеристик, отличающих эту фракцию ядерных РНК - это чрезвычайно высокая вариабельность размеров входящих в нее транскриптов.
По мере синтеза эти транскрипты ковалентно модифицируются по 5'-концам и 3'-концам таким образом, что они становятся отличными от транскриптов, синтезированных другими РНК-полимеразами. Эти модификации будут позже использованы в цитоплазме как сигналы того, что данные информационные РНК должны быть транслированы в белки.
Все 3 класса РНК транскрибируются с генов, которые содержат интроны. Последовательности, кодируемые интронами ДНК, должны быть удалены из первичного транскрипта до того, как РНК станет биологически активной. Процесс удаления копий интронных последовательностей получил название сплайсинга РНК.
Кэпирование и полиаденилирование иРнКопределяют дальнейшие особенности функций иРнк
В дополнение сплайсингу, у и-РНК происходит модификация 3’и 5’ концов. К 5 ' концу всех эукариотических иРНК (который синтезируется первым в процессе транскрипции) присоединяется во время процессинга остаток 7-метилгуанозина с образованием уникальной 5 ' 5 ' фосфодиэфирной связи. Этот дополнительный нуклеотид получил название кэп или колпачек. Кэпирование происходит еще до завершения синтеза всей молекулы. Образующаяся структура на 5’ конце иРНК защищает РНК от экзонуклеаз и, что не менее важно, ответственна за последующее связывание молекулы мРНК с рибосомой.
Р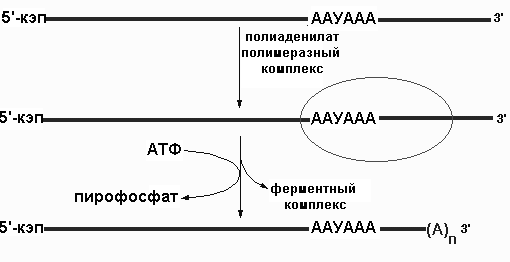 ис.8.35.Схема
полиаденилирования иРНК.
ис.8.35.Схема
полиаденилирования иРНК.
Сразу после завершения транскрипции или после специфического расщепления в определенном месте растущей цепи РНК происходит полиаденилирование. Оно заключается в том, что специальный фермент - полиаденилатполимераза присоединяет к 3'-концу каждого РНК-транскрипта, которому суждено стать молекулой мРНК, от 20 до 250 остатков адениловой кислоты (поли(А)), что и завершает процесс образования первичного РНК-транскрипта. Полиаденилатполимераза узнает специфическую последовательность AAУAAA. Этот фермент, обладающий несколькими активностями отщепляет от первичного транскрипта небольшрй фрагмент в 11-30 нуклеотидов и затем присоединяет поли(А) последовательность Функции этой последовательности неизвестны. Принять считать, что такой "хвост" способствует последующему процессингу РНК и экспорту зрелых молекул мРНК из ядра.
Процессы 5'-кэпирования и 3'- полиаденилирования характерны только для транскриптов, синтезируемых РНК-полимеразой II. Это можно объяснить специфическим взаимодействием ферментов, катализирующие эти процессы с РНК-полимеразой II, но не взаимодействующих с РНК-полимеразами I и III . Необходимость маркировать подобным образом концы молекул-предшественников и-РНК может служить объяснением, почему эти молекулы синтезируются специальной РНК-полимеразой.