

Д ля перемещения днк-полимеразы молекулу матрицы следует «раскрутить»
Рис.8.25.Роль топоизомеразы в механизме репликации
Быстрое перемещение репликационной вилки обеспечивается непрерывным раскручиванием двойной спирали ДНК. Это способствует возникновению перед вилкой сильного торзионного напряжение двойной спирали ДНК. Оно снимается при помощи специальных ферментов, названных ДНК топоизомеразами. Топоизомеразы уменьшают торзионные напряжения двойных спиралей ДНК, катализируя расщепление двойной - (топоизомеразы II) или одной цепи ДНК (топоизомеразы I). Разрывы цепей затем вновь восстанавливаются топоизомеразами.
Ретровирусы внесли изменения в центральную догму молекулярной билогии.
Особой формой репликации у прокариот является репликация с использованием РНК в качестве матрицы для синтеза ДНК. Такая форма репликация обнаружена у ретровирусов (например, у HIV ретровируса, вызывающего СПИД) и катализируется ферментом РНК зависимой ДНК полимеразой или обратной транскриптазой.
Подобно другим РНК зависимым полимеразам, обратная транскриптаза - обладает высокой склонностью к ошибкам, посколько не обладает коррегирующей активностью. Схема репликации РНК ретровирусов следующая:
1. Вирусная РНК в клетке хозяина образует пары оснований со специфической молекулой тРНК, образуя праймер для репликации ДНК.
2. Обратная транскриптаза синтезирует ДНК с образованием смешаной ДНК-РНК молекулы.
3. Рибонуклеаза H частично разрушает цепь РНК двойной РНК-ДНК спирали, удаляя 5 ' концевой отдел.
4. 3 ' конец вирусной РНК образует пары с освободившейся цепью ДНК, формируя кольцевую структуру.
5. Обратная транскриптаза создает копию цепи ДНК генома, используя 3 ' конец цепи ДНК в качестве праймера.
6. Оставшаяся цепь РНК удаляется , и вновь синтезированная цепь ДНК образует комплементраную двойную спираль с первой цепью ДНК.
7. Образовавшаяся полная молекула ДНК внедряется в хромосому клетки- хозяина
Препарат 3 '-азидо-2 ' 3 '-дидезокситимидин (АZT), после превращения в соответствующий 5 ' трифосфат в клетках, является ингибитором фермента обратной транскриптазы вируса HIV. Другие аналоги нуклеозидов , типа 2 ' 3 '-дидезоксицитидин (ддЦ) , 2 ' 3 '-дидезоксиинозин и 2 ' 3 '-дидегидро-3 '-дезокситимидин (d4T) после фосфорилирования в соответствующие трифосфаты блокируют удлинение цепи ДНК после включения в ДНК.
Биосинтез днк у эукариот связан с циклом деления клетки.
В отличие от прокариот, которые делятся постоянно, синтез ДНК у эукариот тесно связан с клеточным циклом. Клеточный цикл включает строго детерминированный ряд последовательных процессов, которые можно разделить на два периода:
1) период клеточного роста , называемый " интерфаза ", и
2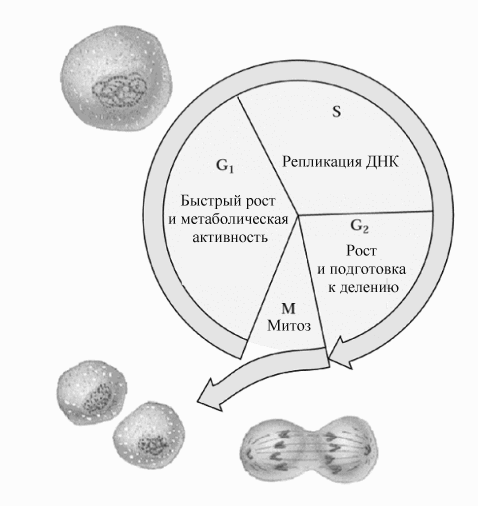 )
период клеточного деления , называемый
" фаза М " (от слова mitosis). В свою оче
)
период клеточного деления , называемый
" фаза М " (от слова mitosis). В свою оче
Рис.8.26.Фазы клеточного цикла.
редь, в каждом периоде выделяют несколько фаз (см рис.8.26). Основное время всего клеточного цикла занимает интерфаза (не менее 90% ) времени. Так, например, у быстро делящихся клеток высших эукариот последовательные деления происходят один раз в 16-24 часа, и фаза М длится 1-2 часа. Большая часть компонентов клетки синтезируется на протяжении всей интерфазы, и это затрудняет выделение в ней отдельных стадий. В интерфазе выделяют фазу G1, фазу S и фазу G2. Репликация ДНК клеточного ядра происходит в " фазу S " (от слова synthesis). Период между фазой М и началом фазы S обозначен как фаза G1 (от слова gap - промежуток), а период между концом фазы S и последующей фазой М - как фаза G2. Период клеточного деления (фаза М ) включает две стадии: митоз (деление клеточного ядра) и цитокинез (деление цитоплазмы).
Клетки могут выходить из митотического цикла на неопределенное время, сохраняя жизнеспособность и пролиферативный потенциал. Такие клетки называют покоящимися клетками, а сам переход называется переходом в состояние пролиферативного покоя или в G0-фазу.