

3.2. Маленькие популяции особенно уязвимы
Один из лучших примеров определения МЖП относится к популяциям 120 снежных баранов (Ovis canadensis), обитающих в пустынях юго-запада США [Berger, 1990; 1999]. Некоторые их этих популяций наблюдали в течение 70 лет. Изучение привело к поразительным результатам. Оказалось, что 100% популяций, насчитывающих менее 50 животных, вымерло в течение 50 лет, в то время как практически все популяции, включающие более 100 особей, сохранились на весь этот период (рис. 3.1). Несмотря на факторы, препятствующие выживанию мелких популяций, государственный контроль за местообитаниями и подселение в них дополнительного количества животных позволило спасти некоторые малые популяции, находившиеся под угрозой вымирания.
Данные, полученные в результате долговременного изучения птиц на California Channel Islands, подтвердили необходимость сохранения больших популяций. Только те популяции, которые насчитывали более 100 пар, имели
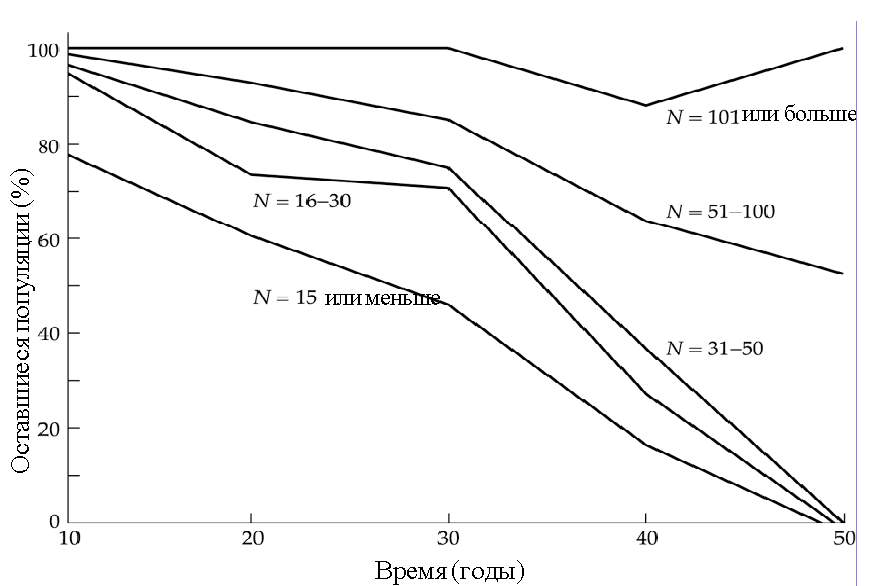
Рис. 3.1. Взаимосвязь между начальной популяцией величиной (N) канадского снежного барана и процентом его оставшейся популяции через некоторое время. Почти все популяции с числом более 100 особей прожили свыше 50 лет, в то время как популяции менее 50 вымерли в течение 50 лет. Не включены небольшие популяции, с которыми идет активная работа и куда добавляют животных [по Berger, 1990].
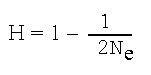
более 90% шансов на выживание в течение 80 лет [Jones, Diamond, 1976]. С другой стороны и малые популяции не всегда сдаются. Многие популяции птиц, состоящие из 10 размножающихся пар, достоверно выжили в течение 80 лет.
3.3. Проблемы малых популяций
За редким исключением, для защиты большинства видов нужны большие популяции, а виды с малыми популяциями стоят перед реальной опасностью вымирания. Вот три основные причины, которые делают малые популяции подверженными быстрому падению численности и локальному исчезновению:
1) генетические проблемы, возникающие из-за потери генетического разнообразия, родственного скрещивания и дрейфа генов;
2) демографические колебания из-за случайного изменения соотношения в уровнях рождаемости и смертности;
3) флуктуации численности, связанные с хищниками, конкуренцией, заболеваниями, равно как с природными катастрофами в виде пожаров, наводнений, засух.
3.4. Потеря генетического разнообразия
Генетическое разнообразие очень важно для обеспечения способности популяций адаптироваться к изменениям окружающей среды (см. главу 1). Только особи с определенными аллелями или комбинацией аллелей могут обладать качествами, необходимыми для выживания и воспроизводства в новых условиях. Внутри популяции частота отдельных аллелей может варьировать от часто встречающихся до очень редких. В малых популяциях частоты аллеля могут изменяться от одного поколения к следующему просто из-за случайностей, которые имеют место при скрещивании и выживании потомства; этот процесс известен как генетический дрейф. Когда аллель в малой популяции имеет низкую встречаемость, существует высокая вероятность того, что он будет случайно потерян в одном из поколений. Рассматривая теоретический пример изолированной популяции, в которой в ген представлен двумя аллелями, Райт (Wright, 1931) предложил формулу для выражения доли гетерозиготности особей (особь, у которой имеются два разных аллеля гена), которые остаются в очередном поколении (Н) в популяции размножающихся взрослых (Ne):
Согласно этому уравнению, популяция из 50 особей в следующей генерации из-за утраты редких аллелей сохранит 99% исходной гетерозиготности, а через 10 поколений – 90%. Однако популяция из 10 особей через поколение сохранит только 95% исходной гетерозиготности, а через 10 поколений – лишь 60% (рис. 3.2).
Из этой формулы наглядно видно, какие значительные потери генетического разнообразия могут происходить в малых популяциях, особенно обитающих на островах или во фрагментированных ландшафтах. Однако миграция особей между популяциями и регулярная мутация генов приводят к увеличению генетической изменчивости внутри популяции и уравновешивают влияние генетического дрейфа. Даже низкая частота перемешивания особей из разных популяций сводит к минимуму потерю генетического разнообразия, обусловленную малыми размерами популяции [Bryant

Рис.3.2. Генетическая изменчивость может быть со временем утрачена вследствии девиации генов. На графике представлен средний про-цент генетической изменчивости, оставшейся после десяти поколений для теоретической популяции величиной (Ne). За 10 поколений генетическая изменчивость уменьшается на 40% при размере популяции 10 особей, 65% при популяции, состоящей из 5 особей и 95% при величине равной 2 [из Meffe, Carroll 1997].
еt al., 1998]. Если в каждую изолированную популяцию размером около 100 особей в каждом поколении прибывает только один новый эмигрант, влияние генетического дрейфа будет пренебрежимо малым. Такой поток генов оказался важнейшим фактором, предотвращающим потерю генетического разнообразия в малых популяциях галапагосских вьюрков [Grant, Grant, 1992]. Хотя обычные темпы мутаций в природе – примерно от 1 из 1000 до 1 из 10 000 генов на генерацию – могут компенсировать случайные потери аллелей в больших популяциях, они не могут компенсировать эффект генетического дрейфа в малых популяциях численностью 100 и менее особей.
Дополняя собой теоретические положения и лабораторные эксперименты, полевые данные также показывают, что малый размер популяции ведет к более быстрой потере аллелей [Frankham, 1996]. Среди хвойных видов в Новой Зеландии популяции, образованные менее чем 1000 растениями, испытывают более сильную потерю генетического разнообразия, чем популяции, превышающие 10 000 экземпляров [Billington, 1991]. Обзор исследований генетической изменчивости растений показал, что только 8 из 113 видов имели генетическое разнообразие ниже предела обнаружения и что большинство из этих 8 видов обладали очень ограниченными ареалами [Hamrick, Godt, 1989].
Подверженные генетическому дрейфу малые популяции более восприимчивы к разрушительным генетическим эффектам, таким как инбредная депрессия, аутбредная депрессия и потеря эволюционной пластичности. Эти факторы могут вносить свой вклад в дальнейшее уменьшение размера популяции и увеличение вероятности вымирания.
Инбредная депрессия. В большинстве природных популяций существуют разнообразные механизмы, предотвращающие родственное спаривание (инбридинг). В крупных популяциях большинства видов животных особи обычно не спариваются с близкими родственниками. Обычно молодые животные покидают то место, где родились, или опасность их спаривания с родственниками подавляется уникальными индивидуальными запахами или другими подсказками органов чувств. У многих растений ряд морфологических и физиологических механизмов способствует перекрестному опылению и предотвращает самоопыление. Однако в некоторых случаях, особенно когда размер популяции мал и нет других партнеров, эти механизмы не могут предотвратить инбридинг. Спаривание между близкими родственниками, например между родителями и их потомством, внутри потомства и между двоюродными родственниками, самооплодотворение у гермафродитных видов могут привести к инбредной депрессии – состоянию, характеризующемуся сокращением числа потомков, появлением слабого или стерильного потомства [Ralls еt al., 1988]. Например, при воспроизводстве популяции гилии пурпурной (Ipomopsis aggregata), состоящей из менее чем 100 растений, семена становятся более мелкими, отличаются пониженной всхожестью и проявляют большую чувствительность к изменениям окружающей среды, чем растения из более крупных популяций (рис. 3.3).

Рис. 3.3. Семена в популяции пурпурной гилии (Ipomopsis aggregata) из горной Аризоны в малых популяциях (менее 150 представителей) прорастают хуже по сравнению с большими. Прорастание семян резко уменьшается в самых малых популяциях [по Heschel, Paige, 1995].
Эти симптомы, связанные с инбредной депрессией и потерей генетического разнообразия, ослабевают, когда растения из малых популяций подвергаются перекрестному опылению пыльцой растений их больших популяций. В Иллинойсе малые изолированные популяции лугового тетерева (Tympanuchus cupido pinnatus) обнаруживали признаки снижения генетического разнообразия и, соответственно, у них падала плодовитость и уменьшался процент вылупления цыплят [Westemeier еt al., 1998]. Когда в эти популяции запустили особей из больших популяций с высоким генетическим разнообразием, жизнеспособность яиц восстановилась, продемонстрировав необходимость поддержания генетической изменчивости. Наиболее очевидное объяснение инбредной депрессии состоит в том, что у потомства проявляется действие вредных аллелей, унаследованных от несущих их обоих родителей [Barrett, Kohn, 1991]. Инбредная депрессия может стать большой проблемой для малых популяций животных, содержащихся в зоопарках, и для реализации программ по селекции домашних животных.
Аутбредная депрессия. В дикой природе особи из географически разделенных и генетически различающихся популяций редко спариваются между собой не только из-за своей физической отдаленности, но и благодаря ряду поведенческих, физиологических и морфологических механизмов, которые гарантируют спаривание только между генетически сходными особями одного вида. Однако, когда вид редок, или его местообитание разрушено, случается аутбридинг – спаривание между представителями разных популяций. Особи, неспособные найти партнера в своей популяции, могут спариваться с особями из других популяций. В результате этого, из-за отсутствия комплементарности хромосом и ферментных систем, унаследованных от своих разных родителей, потомство бывает ослабленным или бесплодным, – ситуация, известная как аутбредная депрессия [Thornhill, 1993]. Гибридное потомство может уже не иметь ту необходимую комбинацию генов, которая позволяет особям выживать в конкретных местных условиях. В своем крайнем проявлении аутбредная депрессия может возникнуть при спаривании между особями близкородственных видов. Чтобы избежать аутбредной депресии, программы по разведению животных в неволе должны предусматривать исключение пар из близкородственных видов и из крайних областей их географического ареала. То есть при создании экспериментальных популяций редких видов следует их ограждать от возможного влияния аутбредной депресии.
Потеря эволюционной пластичности. Редкие аллели или необычные комбинации аллелей, не дающие сегодня никаких преимуществ, могут оказаться уникально полезными для будущих условий среды. Потеря генетического разнообразия в малой популяции может ограничить способность популяции адекватно откликаться на долговременные изменения в окружающей среде, такие как загрязнение, новые болезни или глобальное изменение климата. Без достаточного генетического разнообразия вид может исчезнуть.