Потребление атф
Роль АТФ в жизнедеятельности организма в целом трудно переоценить.Среди наиболее важных потребителей АТФ следует отметить такие, как:
1. Большинство анаболических реакций (anabolic reactions) в клетках проходят с использованием АТФ, т.е.:
-синтез белков из аминокислот,
-синтез ДНК и РНК из нуклеотидов,
-синтез полисахаридов,
-синтез жиров
2. АТФ необходима для активного трансмембранного транспорта молекул и ионов,
для индукции и проведения нервного импульса(nerve impulses),
поддержания клеточного объема через механизмы осмоса ( osmosis),
для мышечных сокращений (muscle contraction),
для осуществления биолюминесценции в тканях (bioluminescence).
3.АТФ является синаптическим передатчиком , широко распространенным в различ-
ных органах, особенно в пресинаптических окончаниях эффекторных нейронов. При стимуляции этих окончаний выделяются пуриновые продукты распада – аденозин и инозин. В эволюционной биологии вообще считается, что АТФ был единственным и общим для всех организмов медиатором на ранних этапах эволюции. В процессе эволюции, в связи с усложнением строения организмов, стали появляться новые специализированные синаптические медиаторы. Исследования на отдельных органах экспериментальных животных показали, что АТФ способствует релаксации гладких мышц органов пищеварения
Нарушения энергопотребления в сердце: причины и последствия
Одним из важнейших каналов энергопотребления в организме человека является деятельность сердца. Непрерывная работа сердца (С) требует устойчивого и надежного энергопотребления. Закупорка одной из питающих само С артерий прекращает кровоснабжение участка сердечной мышцы и возникает ишемия ткани. Продолжительный период ишемии приводит к гибели клеток сердечной мышцы, кардиомиоцитов – тогда развивается инфаркт миокарда. Но, если спазм сосудов был непродолжительным, и кровоток в нем восстанавливается, то сократительная работа миокарда может быть полностью восстановлена. Особенно важной эта проблема стала в связи с развитием технологии пересадок сердца. Как же осуществляется снабжение энергией клеток сердечной мышцы и ее использование?
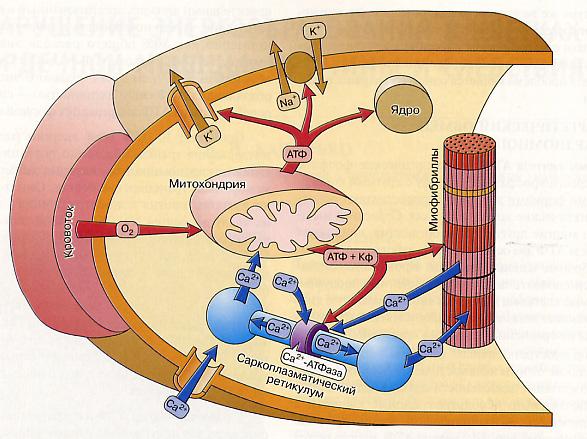
Рис.45 Основные энергопотребляющие структуры кардиомиоцита.
Регуляция сократительной функции кардиомиоцитов происходит посредством ионов кальция. Они поступают в клетку снаружи и вызывают высвобождение ионов кальция, содержащихся в цистернах саркоплазматического ретикулума. Эти ионы связываются с миофибриллами и вызывают их сокращение.
Основным потребителем АТФ в кардиомиоцитах является сократительный аппарат миофибрилл( Рис 45); его потребность в энергии оценивается около 80% общего расхода энергии. Примерно 10-15% энергии расходуется на поддержание трансмембранного потенциала и возбудимости кардиомиоцитов. И около 5% энергии клетки используется для синтетических процессов. Помимо АТФ,источником энергии в клетках может быть другое высокоэнергетичное соединение – креатин-фосфат( Кф) ,более эффективно используемое клеткой, чем АТФ.
Процесс энергообразования в кардиомиоцитах нарушается по разным причинам. При внезапной ишемии миокарда прекращается синтез АТФ в митохондриях, быстро снижается содержание Кф и АТФ. При этом, наиболее глубоко нарушаются функции сократительного аппарата.
Защитные механизмы при ишемии.
В процесс развития ишемии в сердечной мышце включаются защитные механизмы, снижающие деструктивные процессы.
1.Происходит открытие АТФ-зависимых калиевых каналов( рис 46).В норме они закрыты а при недостаточном ре-синтезе АТФ они открываются и калий активно выходит из клеток. Это сопровождается снижением мембранного потенциала и возбудимости клеток.
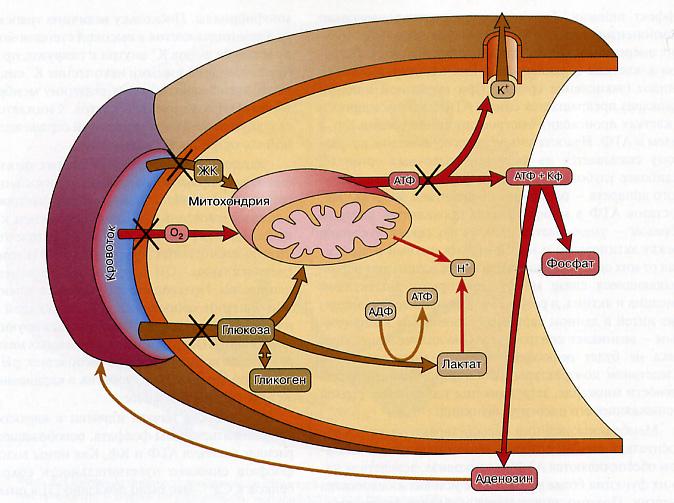
Рис.46. Метаболические последствия ишемии миокарда
2. Происходит закисление цитоплазмы клеток – развивается ацидоз. Прекращение окисления в митохондриях при ишемии приводит к активации гликолиза,накоплению недоокисленных продуктов,
увеличению концентрации ионов водорода и сдвигу рН.
3. Распад АТФ и Кф сопровождается накоплением фосфата в сердечных клетках. Это снижает чувствительность сократительных белков к ионам Са+2.
4. Накопление аденозина как результат распада АТФ блокирует аденорецепторы на кардиомиоцитах. В результате нейромедиатор норадреналин не активирует сердечные клетки и предотвращает снижение запасов АТФ и Кф.
Таким образом, уже в начале стадии ишемии активируются несколько защитных механизмов, снижающих вход ионов кальция в кардиомиоциты и чувствительность сократительного аппарата к действию ионов кальция. При ишемии уровень сократительной функции падает очень быстро( в пределах 30 сек) примерно до 5-10% от исходного, тогда как содержание АВТФ и Кф снижается умеренно.Это и позволяет клеткам сердца экономично расходовать энергию и переживать неблагоприятный период. При длительной ишемии ( неск.часов) энергодефицит усугубляется, усиливается ацидоз – это приводит к разрушению клеточных органелл и некрозу клеток.
Внезапное подавление синтеза АТФ при ишемии могло бы вызвать гибель кардиомиоцитов в течение неск.минут, если бы в сердце не работали естественные защитные механизмы. Они быстро подавляют сократительную активность и обеспечивают экономичное расходование запасов энергии в течение десятков минут.Устранение причин,вызвавших ишемию, в этот период может восстановить сократимость кардиомиоцитов, работу сердца.
Зона ишемии может быть уменьшена- с помощью введения аденозина, ионов калия а также окисью азота NO, обладающей сосудорасширяющим действием. Оказалось, что именно окись азота является посредником действия многих сосудорасширяющих веществ, таких например, как нитроглицерин.
.
Клеточное ядро
Термин “ядро “ был введен Брауном в 1833 году, когда он впервые описал постоянные структуры шаровидной формы в растительных клетках. Позднее такие же структуры были обнаружены во всех клетках высших организмов, в том числе и у человека.
Клеточное ядро, обычно одно в клетке, состоит из ядерной оболочки, отделяющей его от цитоплазмы,хроматина, ядрышка, ядерного белкового матрикса( остова) икариоплазмы( ядерного сока) ( рис 27 Ченцов).
Зернистый
эндоплазматический ретикулум Ядерная
пора ядрышко рибосомы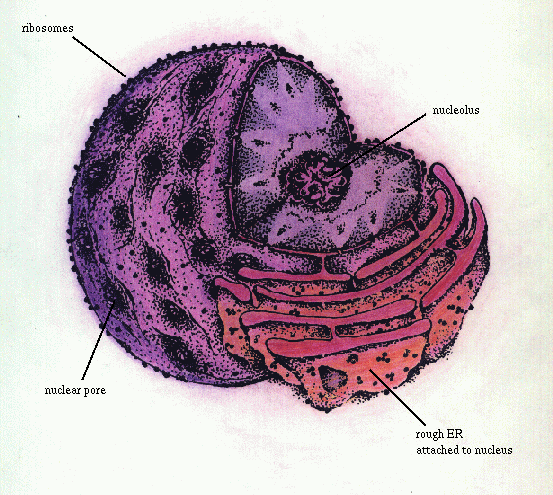
Рис.47. Клеточное ядро
Эти компоненты ядра имеются во всех клетках эукариотов- одно- и многоклеточных.
Ядро(nucleus) клетки – структура, содержащая генетическую информацию о клетке и целостном организме. Ядро осуществляет две группы общих функций: 1- хранение генетической информации., 2- ее реализация в виде синтеза белка.
1. Хранение и поддержание наследственной информации в виде неизменной структуры ДНК связано с работой т.н. ферментов репарации, ликвидирующих спонтанно возникающие повреждения молекул ДНК. Ферменты репарации работают и в клетках, поврежденных радиацией, способствуя более или менее эффективному восстановлению клеток от радиационных повреждений. Открыл этот вид репарации известный обнинский радиобиолог проф. Лучник Н.В. в 70-х годах 20 столетия.
2. Другая сторона деятельности ядра – работа аппарата белкового синтеза. В ядре также синтезируются компоненты рибосом. Из общей схемы белкового синтеза( рис 16 Ченцов) видно, что источником информации для начала биосинтеза является ДНК
Структура и химический состав клеточного ядра
В подавляющем большинстве клеток высших млекопитающих содержится лишь одно ядро, хотя есть и многоядерные клетки – например, клетки мышечного волокна –миосимпласты.
Ядерный хроматин представляет собой плотное вещество, заполняющее почти весь объем ядра. В неделящихся( интерфазных) клетках оно диффузно распределено по объему ядра, в делящихся клетках оно уплотняется( конденсируется) и образует плотные структуры – хромосомы. Хроматин хорошо прокрашивается основными красителями, поэтому и получил свое название (от греч.chroma– цвет, краска). В составе хроматина – ДНК в комплексе с белками – гистоновыми (щелочными) и негистоновыми. Диффузный хроматин интерфазных ядер генетики называютэухроматином, конденсированный хроматин –гетерохроматином. В обоих формах хроматин представляет собой фибриллы толщиной 20-25 нм.
Известно, что длина отдельных молекул ДНК может достигать сотен микрон и даже приближаться к сантиметру. В хромосомной наборе человека самая длинная хромосома – первая, длиной до 4 см. В хромосомах имеется множество мест независимой репликации ( удвоения) – репликонов. Таким образом, ДНК представляет собой цепочку тандемно расположенных репликонов различного размера.
Белки хроматина составляют 60-70% от его сухого веса. Гистоны (щелочные белки) расположены вдоль молекулы ДНК не равномерно, а в виде блоков, в каждый из них входит по восемь молекул гистона, формируя структуру нуклеосому. Процесс формирования нуклеосомы сопровождается сверхспирализацией ДНК и укорочением ее длины примерно в 7 раз.
В ядре, кроме ДНК, присутствуют также и молекулы РНК- информационной, связанной с белками.
Хромосомный цикл
Общеизвестно, что половые женские и мужские клетки несут одинарный набор хромосом и следовательно, содержат в 2 раза меньше ДНК, чем остальные клетки организма. Половые клетки с одинарным набором хромосом называются гаплоидными . Плоидность, т.е.кратность, обозначается в генетике буквойn. Так, клетки с набором 1nгаплоидны, с 2n-диплоидны, с 3n–триплоидны. Соответственно, количество ДНК в клетке( обозначается буквой с) зависит от ее плоидности: клетки с 2n-количеством хромосом содержат 2с количество ДНК. При оплодотворении сливается две гаплоидные клетки, каждая из которых несет набор 1nхромосом, поэтому образуетсядиплоидная( 2n,2c) клетка – зигота. Затем, за счет деления диплоидной зиготы и последующего деления диплоидных клеток будет развиваться организм,клетки которого будут диплоидными , а часть из них ( половые) снова будут гаплоидными.
Однако, процессу деления диплоидных клеток предшествует фаза синтеза ДНК-редупликация, т.е. появляются клетки с количеством ДНК, равным 4с, у них количество хромосом – 4 n. И только после деления такой тетраплоидной( 4с) клетки возникают две новые диплоидные клетки.
В ядрах интерфазных( покоящихся) клеток хромосомы увидеть трудно. Они появляются в ядре незадолго до клеточного деления. В интерфазе, тем не менее, происходит удвоение, редупликация хромосом. В этом периоде происходит синтез ДНК, поэтому он называется синтетическим, или S-периодом. В этот период в клетках обнаруживается количество ДНК, большее чем 2с. После окончанияS-периода количество ДНК в клетке составляет 4с( полное удвоение хромосомного материала). Если подсчитать количество хромосом в профазе, то их будет 2n, но это ложное впечатление., т.к. в это время каждая хромосома двойная( как результат редупликации).На этой стадии пара хромосом тесно соприкасается друг с другом, они закручиваются одна вокруг другой. Следовательно, уже в начале профазы хромосомы состоят из двух сестринских хромосом – хроматид. Они остаются связанными друг с другом и в следующей фазе – метафазе, когда хромосомы выстраиваются в экваториальной плоскости клетки. В следующей стадии – анафазе, происходит расхождение пар гомологичных хромосом к противоположным полюсам клетки, после чего клетка делится. Затем в телофазе разошедшиеся диплоидные наборы( 2n) хромосом начинают деконденсироваться, т.е. разрыхляться. Так заканчивается один хромосомный цикл и начинается следующий.( Рис 31 Ченцов). Хромосомный( клеточный ) цикл у многоклеточных эукариотов длится1-1,5 суток.
Ядрышко (Nucleolus)
В ядре всех эукариотических клеток видно одно или несколько телец округлой формы – ядрышки. Они хорошо окрашиваются основными красителями, поскольку богаты РНК. Ядрышко – это производное хромосом, при этом оно является самостоятельной органеллой, функция которой заключается в образовании рибосомных РНК и рибосом. Ядрышко неоднородно по строению – центральная часть фибриллярная, где сосредоточены предшественники рибосом, и периферия – гранулярная, где концентрируются созревающие субъединицы рибосом.
Ядерная оболочка (кариолемма)
Представляет собой структуру, ограничивающую клеточное ядро. Она разделяет два внутриклеточных компонента друг от друга – ядро от цитоплазмы. Значение такого разделения структур в пространстве важно: это создает дополнительные (в сравнении с прокариотами) возможности для регуляции генной активности при синтезе специфических белков.
Ядерная оболочка состоит из двух мембран – внешней и внутренней, между которыми располагается перинуклеарное пространство (Рис106 Ченцова). В общем виде ядерная оболочка может быть представлена как двуслойный мешок, отделяющий содержимое ядра от цитоплазмы. Однако ядерная оболочка имеет характерную особенность, отличаюшую ее от других мембранных структур клетки – это особые ядерные поры, образующиеся за счет слияния двух ядерных мембран.
Внешнюю мембрану ядерной оболочки относят к мембранной системе эндоплазматического ретикулума – на ней расположены многочисленные полирибосомы, и сама ядерная мембрана переходит в мембраны ретикулума. Внутренняя мембрана ядерной оболочки не имеет на своей поверхности рибосом. Однако, она связана с хроматином и это является характерной особенностью внутренней ядерной мембраны.
Еще одна функция ядерной оболочки – создание внутриядерного порядка, архитектуры, фиксация хромосомного материала в трехмерном пространстве.
Ядерные поры – результат слияния двух ядерных мембран. Отверстия пор – диаметром около 90 нм. Комплекс ядерных пор, включающий 8 периферических белковых гранул и одну центральную, участвует в транспорте молекул белков и нуклеопротеидов, в распознавании этих молекул. Этот процесс транспорта активный и требует затрат АТФ. В среднем на одно ядро приходится несколько тысяч поровых комплексов.( рис 109 Ченц).
Ядерный белковый матрикс
Процессы репликации (удвоения) и транскрипции (считывания информации) хроматина осуществляется в ядре строго упорядоченно. Для реализации этих процессов существует внутриядерная система, объединяющая все ядерные компоненты – хроматин, ядрышко, ядерную оболочку. Такой структурой является ядерный белковый остов, или матрикс (ЯБМ). При этом, он не представляет собой четкой морфологической структуры. По морфологической композиции ЯБМ состоит из трех компонентов: сетчатого белкового слоя- ламины, внутренней сети- остова и “ остаточного “ ядрышка. Основной компонент структур ЯБМ – фибриллярные белки, близкие по аминокислотному составу к промежуточным микрофиламентам.
Роль ядерной оболочки в ядерно-цитоплазматическом обмене.
Ядерная оболочка служит регулятором в ядерно-цитоплазматическим обмене. Обмен продуктами между ядром и цитоплазмой очень велик: все ядерные белки поступают в ядро из цитоплазмы, и все РНК выводятся из ядра. Комплексы ядерных пор в этом процессе выполняют роль не только механизма переноса (транслокатора), но и роль сортировщика переносимого материала. Через поры в ядро поступают пассивным транспортом ионы, сахара, нуклеотиды, АТФ и гормоны. Методом пассивного транспорта в обе стороны проникают через ядерную мембрану высокомолекулярные соединения с массой не более 5.103Да. Через ядерные поры осуществляется и активный транспорт макромолекул в обе стороны.
Немембранные органеллы.
Рибосомы. Эти специализированные органеллы клетки обеспечивают синтез белков и полипептидов. Присутствуют у всех типов животных клеток и представляют собой высокомолекулярные рибонуклеопротеиды. В состав рибосом (Р) входят белки и особый вид РНК, называемый рибосомальной РНК(р-РНК). Размер Р – 20 х 20 х 20 нм. Состоит из большой и малой субъединиц. Каждая из субъединиц сформирована из рибонуклеопротеидного тяжа. В клетках имеются отдельные Р и их комплексы – полирибосомы. Они могут свободно располагаться в гиалоплазме или быть связанными с мембранами эндоплазматического ретикулума . Обычно, свободные Р содержатся в малоспециализированных и быстрорастущих клетках, тогда как связанные с ретикулумом – в специализированных клетках. Кроме того, свободные Р синтезируют белок для собственных нужд клетки, а связанные – на экспорт.
Рис. “Грозди “ рибосом
Цитоскелет. Это опорно-двигательная система в клетке, создающая действительно клеточный скелет( Рис). Система содержит белковые нитчатые образования. Нитчатые и фибриллярные структуры цитоскелета являются динамичными образованиями, возникающими и исчезающими в зависимости от функционального состояния клетки. Основные компоненты цитоскелета –микротрубочки и микрофиламенты.
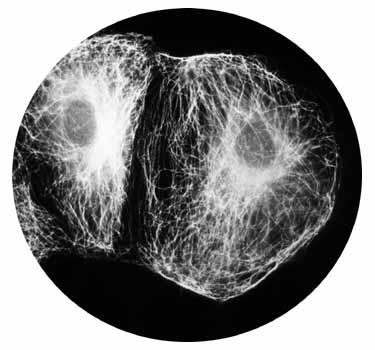
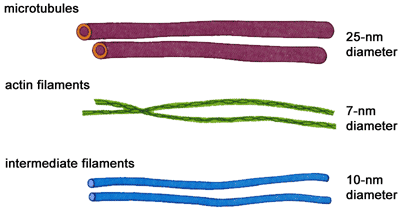
Методами иммунофлуоресценции установлено, что в состав микрофиламентов входят сократительные белки –актин, миозин, тропомиозин. Т.е.,микрофиламенты – это ни что иное, как сократительный аппарат клетки, обеспечивающий подвижность как самой клетки, так и органелл внутри нее. Микрофиламенты имеют толщину 5-7 нм.
Микротрубочки принимают участие в создании временных ( веретено деления, цитоскелет интерфазных клеток ) и постоянных структур ( центриоли, реснички, жгутики). Они представляют собой прямые неветвящиеся полые цилиндры диаметром 24 нм, толщина стенки цилиндра – 5 нм. В электронном микроскопе на поперечном срезе микротрубочек видны 13 субъединиц белка тубулина.
Клеточный центр ( центросома).Состоит из центриолей и связанных с ними микротрубочек. Методами электронной микроскопии удалось исследовать тонкое строение центриолей. Основу этой структуры составляют 9 триплетов микротрубочек, образующих полый цилиндр.( рис). Его ширина около 2 нм, длина – 3-5 нм.
Обычно, в интерфазных клетках присутствуют две центриоли, образуя единую структуру – диплосому. В ней центриоли расположены под прямым углом друг к другу. Из двух центриолей различают материнскую и дочернюю. Торец дочерней центриоли направлен к поверхности материнской центриоли.
При подготовке к митотическому делению в клетке происходит удвоение центриолей. Интересно то, что увеличение числа центриолей не связано с их делением, почкованием или фрагментацией, но есть результат формирования зачатка рядом с исходной центриолью.
Перед митозом центриоли служат центром формирования веретена деления из микротрубочек.
Помимо названных структур, в состав некоторых клеток входят реснички и жгутики, представляющие собой выросты цитоплазмы. Внутри этих выростов присутствует сложная сократительная система из микротрубочек и сократительных белков типа тубулин и динеин. С помощью ресничек и жгутиков клетки осуществляют движения.
Лекция 5. Целостные реакции клеток
Все многообразие превращения веществ в клетках складывается из цепей биохимических реакций . Для реализации биохимических реакций необходимо поступление веществ в клетку – эндоцитоз,превращение веществ в клетке –метаболизм, и выведение конечных продуктов обмена в виде ненужных шлаков или необходимых для организма биологически-активных веществ –экзоцитоз.
Эндоцитоз. Существует несколько способов реализации эндоцитоза:
Трансмембранный пассивный и активный транспорт веществ в клетку. Об этой форме эндоцитоза рассказано в соответствующей главе.
Пиноцитоз – захват клеткой жидких коллоидных частиц.
Фагоцитоз – захват клеткой плотных и крупных корпускулярных частиц вплоть до захвата других клеток.
Вообще, поступление в клетку твердых или жидких субстанций извне называется общим термином – гетерофагия. Этот процесс важен для организма человека в таких органах и системах, как защитная ( фагоцитарная активность нейтрофилов крови, макрофагов), перестройки в костной ткани( остеокласты), образование гормона тироксина в фолликулах щитовидной железы, обратное всасывание белка и других макромолекул в канальцах нефрона почек.
Клеточный метаболизм , или обмен веществ, представляет собой совокупность процессов биосинтеза сложных биологических молекул из более простых (ассимиляция) и реакций расщепления сложных макромолекул с высвобождением тепловой энергии, используемой клетками для различных целей( диссимиляция).
Клетка эффективно использует энергию, заключенную в химических связях поступающих с пищей белков, углеводов и жиров, и высвобождающуюся в процессе их расщепления ( гидролиза) в пищеварительном тракте. Т.е., клеточный метаболизм осуществляется по правилам первого закона термодинамики – энергия не возникает и не уничтожается, она переходит из одного вида в другой, пригодный для выполнения работы.
Схематично процессы диссимиляции питательных веществ происходят таким образом, что на начальном этапе в пищеварительном тракте они расщепляются до мономеров ( белки – до аминокислот, жиры – до жирных кислот, углеводы – до моносахаров), после чего, вне зависимости от природы питательных веществ, дальнейшее
Экзоцитоз.Выведение веществ из клеток осуществляется также с помощью нескольких механизмов. Так же, как и эндоцитоз, имеют место активный и пассивный транспорт веществ из клетки. Активным транспортом выводятся ионы и мелкие молекулы, пассивным – большинство неорганических веществ и конечных продуктов метаболизма ( т.н. шлаки).
Еще один способ выведения имеется для выведения из клеток крупномолекулярных соединений. Они накапливаются в цитоплазме в аппарате Гольджи в виде транспортных пузырьков и с помощью системы микротрубочек концентрируются у плазматической мембраны клетки. Мембрана пузырька встраивается в плазматическую мембрану, и содержимое пузырька выводится за пределы клетки. Слияние пузырька с плазмолеммой может происходить без каких-либо дополнительных сигналов – такой экзоцитоз называется конститутивным. Таким образом выводятся ,как правило, продукты собственного метаболизма клетки, или шлаки. Но значительная часть клеток синтезирует специальные вещества, необходимые организму для жизнедеятельности –секреты.Для того, чтобы пузырек с секретом слился с плазмалеммой, необходим сигнал извне. Такой экзоцитоз называетсярегулируемым.Сигнальные молекулы, стимулирующие выведение секретов, называются либеринами, тормозящие выведение – статинами .Весьма распространен этот способ экзоцитоза в нейро-эндокринной системе при выработке гормонов и нейро- медиаторов.
Межклеточные взаимодействия
Функции клеток
Воспроизведение клеток. Клеточный цикл.
Согласно одному из постулатов клеточной теории, размножение клеток, т.е. увеличение их численности, происходит путем деления исходной клетки. Это правило справедливо как для эукариотических, так и для прокариотических, клеток. Время жизни клетки как таковой, от деления до следующего деления, или от деления до ее гибели, называется клеточным циклом.В организме клетки разных тканей и органов имеют неодинаковую способность к делению. Например, во всех органах имеются клеточные популяции, полностью потерявшие способность к делению. Это специализированные ,или дифференцированные, клетки. Они выполняют , как правило, особые функции, присущие только этому типу клеток и входят в состав паренхимы органов.
Но в организме есть и постоянно обновляющиеся ткани – эпителиальные, кроветворные ткани. В таких тканях существует довольно большая доля активно делящихся клеток, заменяющих отжившие. Длительность цикла обычно составляет в активно делящихся популяциях клеток 10-30 ч. Делящиеся клетки обладают различным количеством ДНК в зависимости от стадии клеточного цикла. Это характерно как для половых, так и соматических клеток. Известно, что мужские и женские половые клетки несут единичный ( гаплоидный) набор хромосом и содержат ДНК в два раза меньше, чем соматические диплоидные клетки всего организма. Плоидность в генетике обозначают буквой n . Так, половые клетки несут 1nнабор, соматические клетки обладают 2nнабором, т.е. они диплоидны, существуют клетки с набором хромосом 3n- это триплоидный набор.
В течение клеточного цикла в популяции диплоидных клеток встречаются как диплоидные, так и тетраплоидные наборы хромосом и промежуточные количества ДНК в период покоя клетки ( интерфаза). Такая гетерогенность обусловлена тем, что удвоение количества ДНК происходит до начала деления ( митоза).
Весь клеточный цикл( анимация в Интернет) состоит из четырех временных отрезков: собственно митоза (М), пресинтетического (G1), синтетического (S) и постсинтетического (G2)периодов интерфазы.
В G1периоде, который следует сразу после митоза, клетки имеют диплоидное содержание ДНК( 2с; с- содержание ДНК, соответствующее плоидности). В этот период начинается рост клеток за счет накопления клеточных белков и происходит подготовка клетки к синтетическому периодуS. Именно в этот период активизируется синтез ферментов,
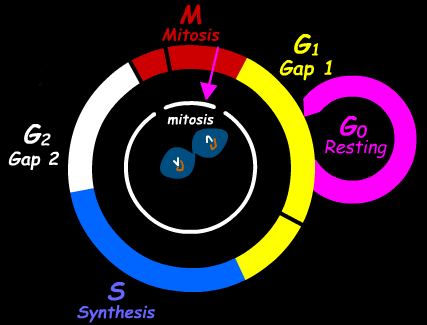
необходимых для образования предшественников ДНК. Резко повышается энергетическая потребность в клетке.
В S- период происходит удвоение количества ДНК ( редупликация) и удваивается количество хромосом ( 1n--- 2n).В разных клетках, находящихся вS- периоде, можно обнаружить и различное количество ДНК – от 2 с до 4с. В этом периоде содержание РНК возрастает в соответствии с количеством ДНК.
Постсинтетическая фаза G2также называется пре-митотической. В этот период активизируется синтез информационной РНК, необходимой для осуществления митоза
В конце этого периода ,перед митозом, синтез РНК падает.
В растущих тканях животных и растений есть некоторое количество клеток, которые находятся как бы вне клеточного цикла. Эти клетки называют клетками G0- периода, или покоящимися. Они не вступают в следующую стадию –G1после митоза, а перестают делиться. При этом, они и не дифференцируются, оставаясь в состоянии, готовом к митозу. Например, большинство клеток печени находятся именно вG0 – периоде – они не синтезируют ДНК и не делятся. Однако, после удаления части печени,как было показано в экспериментальных условиях на животных, большая часть клеток печени включается в митотический цикл. Многие клетки полностью теряют способность возвращаться в митотический цикл – например ,нейроны головного мозга.
Клеточный цикл и радиочувствительность
Интересно отметить, что разные стадии клеточного цикла значительно различаются по чувствительности к внешним воздействиям. Например, наиболее чувствительны к химическим агентам и физическим воздействиям, таким как ионизирующая радиация, G1- период а также собственно митоз, тогда как основная часть интерфазы ( периодыG2иS) менее чувствительна. Экспериментальные исследования на клеточных культурах, полученных от облученных животных, показали, что различия в радиочувствительности могут достигать 40 и более раз между стадиями клеточного цикла. Кроме того, у подвергнутых облучению клеток изменяются временные показатели отдельных стадий цикла; например, отмечается задержка наступления митотической стадии и удлинение стадииG2.
В радиационной цитологии известны феномены самопроизвольного восстановления жизнедеятельности облученных клеток от потенциально летальных повреждений. Этот эффект был обнаружен русским исследователем В.И.Корогодиным в 50-х годах 20-го столетия и имел существенное значение для оценки истинной радиочувствительности клеток и организма в целом.
Деление клеток: митоз.
Митоз ( кариокинез, непрямое деление) – универсальный способ деления любых эукариотических клеток ( анимационный фильм в Интернет). Уже синтезированные в предыдущий пре-митотический периодG2хромосомы ( двойной набор- 4n) переходят в конденсированную компактную форму , образуется веретено деления в клетке и происходит расхождение гомологичных хромосом к противоположным полюсам клетки, после чего разделяется и цитоплазма клетки( цитокинез, цитотомия).
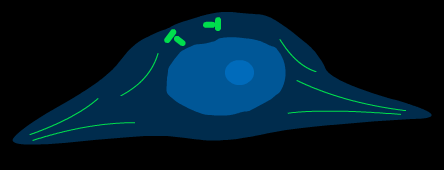
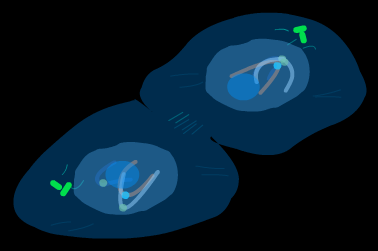
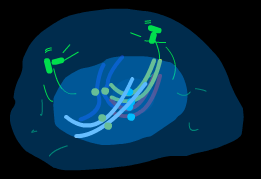
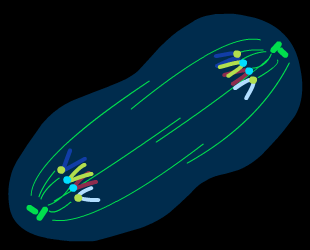
Процесс митоза условно подразделяют на несколько основных фаз: профаза, метафаза, анафаза, телофаза.( рис )
Профаза.Как уже отмечалось, по окончанииS- периода в интерфазном ядре клетки количество ДНК составляет 4 с, поскольку уже произошло удвоение хромосомного материала. Однако морфологически в световом микроскопе число хромосом в профазе различается как 2n, хотя каждая из них уже удвоилась. Однако, к концу профазы двойственность набора хромосом уже морфологически различима за счет активно идущего процесса конденсации ( уплотнения) хромосом. Число хромосом 4nточно соответствует количеству ДНК – 4 с.
![]()
профаза
прометафаза телофаза
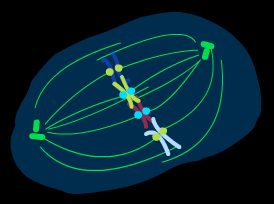
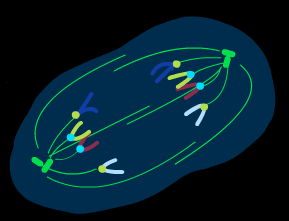
метафаза
анафаза
Рис. Митоз
В профазе значительно снижается уровень синтетических процессов в клетке, и образуется веретено деления – аппарат для разведения генетического материала к двум полюсам клетки.
Метафаза. Эта фаза занимает примерно треть всего времени митоза. Ее тличительной особенностью является то, что в это время заканчивается образование веретена деления и хромосомы выстраиваются в экваторе веретена деления в середине клетки. Клетка в этой фазе имеет характерный вид, называемый “ метафазной пластинкой”, или материнской звездой. К концу метафазы уже хорошо различимы в световом микроскопе удвоенные и конденсированные хромосомы в виде близко прилегающих друг к другу сестринских хроматид. Их плечи располагаются параллельно друг другу, однако между хроматидами уже имеется разделительное пространство.
Анафаза.Выстроившиеся в центре клетки гомологичные хромосомы теряют связь друг с другом и синхронно начинают расходиться к противоположным полюсам пока еще не разделившейся клетки. Это самая короткая по длительности фаза митоза .Тем не менее, в это время происходят важные события : обособление двух идентичных наборов хромосом и их перемещение к двум полюсам клетки.
Телофаза.Два набора хромосом ( 2nх 2) формируют два ядра клетки; одновременно с этим идет процесс разделения исходной клетки на две дочерние –цитокинез, цитотомия. В подмембранном слое цитоплазмы располагаются сократительные белки типа актиновых фибрилл, ориетированные в зоне экватора клетки. Эти белки и осуществляют “ перетяжку” клетки в центре и ее разделение на две дочерние. Митоз завершается.
При повреждении митотического аппарата может происходить задержка митоза в метафазе или даже рассеивание хромосом. Кроме того, могут возникать многополюсные и ассиметричные митозы. При нарушениях процессов цитотомии образуются гигантские ядра или многоядерные клетки.Такие эффекты наблюдаются при злокачественной трансформации клеток как под действием внешних источников( химические агенты, лекарства, ионизирующая и неионизирующая радиация, вирусы ), так и под действием внутренних факторов(например- некоторых гормонов, биологически активных веществ).
Мейоз
Помимо митотического деления соматических клеток, происходящего во всех органах и тканях организма, существует особая, уникальная форма клеточного размножения, приводящая к образованию половых клеток с гаплоидным набором хромосом. Такая форма получила название мейоза и происходит она у высших животных (и человека) и высших растений в первичных генеративных органах. У человека образование половых клеток( гамет) происходит в яичках( у мужчин) и яичниках ( у женщин).
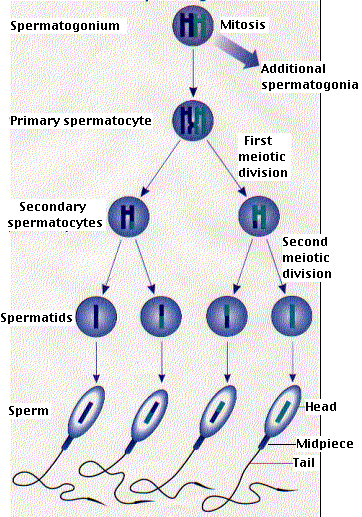
Образование мужских половых клеток ( сперматогенез) протекает в ткани извитых семенных канальцев яичек и включает 4 последовательные стадии: размножения, роста, созревания и формирования( рис).
Начальная фаза сперматогенеза – размножениесперматогенного эпителия и формирование более зрелых клеток – сперматогоний. Среди сперматогоний имеется пул стволовых клеток, являющихся источником образования новых клеток ,и другая часть сперматогоний продолжает дальнейшее созревание, или дифференцировку.
Результатом такого созревания( фаза роста) является потеря клеткой способности к делению формирование клетки, называемой первичным сперматоцитом, или сперматоцитом 1-го порядка. В этот период сперматоцит 1-го порядка увеличивается в объеме и вступает в стадию первого деления мейоза( редукционное деление). Эта стадия длительная и состоит из 5 этапов: лептотены, зиготены, пахитены, диплотены и диакинеза.После деления в каждой из двух дочерних клеток, называемых сперматоцитами 2-го порядка, или вторичными сперматоцитами, содержится уже гаплоидное число хромосом( 23 у человека).
Следующая фаза – второе деление( фаза созревания), происходящее как обычный митоз во вторичных сперматоцитах без редупликации( удвоения) хромосом. В результате, из двух вторичных сперматоцитов образуется четыре клетки с гаплоидным хромосомным набором-сперматиды. Таким образом, каждая исходная сперматогония дает начало 4 сперматидам, у которых гаплоидный(одинарный) набор хромосом.
Сперматиды больше не делятся и после сложных морфологических изменений превращаются в зрелые сперматозоиды. Эта трансформация происходит в заключительную стадию сперматогенеза – фазу формированияспермиев.
Сперматогония (диплоидная
клетка)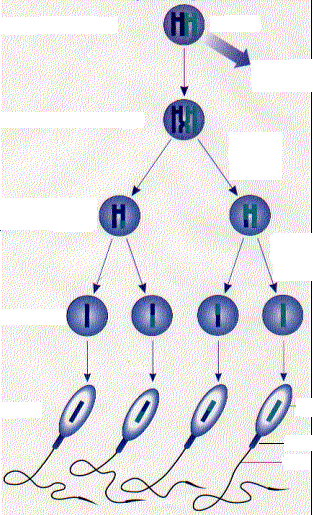
митоз
Дополнительная
сперматогония
Первичный
сперматоцит
Первое
мейотическое деление
Вторичный
сперматоцит
Второе
мейотическое деление
Сперматиды(гапло-идные
клетки)
Сперматозоид Головка Шейка Хвост
Рис. Сперматогенез
Процесс сперматогенеза у человека длится около 75 суток и протекает на протяжении извитого семенного канальца волнообразно. В определенном отделе канальца имеется определенный набор клеток сперматогенного эпителия.
Лекция 6 . Реакции клеток на внешние воздействия.
Клетки организма постоянно подвержены воздействию разнообразных факторов внешней среды – химических, физических и биологических, а также внутренним влияниям – нервным и нейро-гуморальным. Эти факторы вызывают первичные нарушения в клеточных структурах, следствием чего наблюдается как правило функциональное нарушение в органе или системе. Судьба клеток зависит от интенсивности воздействия, его характера и длительности. После нарушений клетки могут адаптироваться, приспособиться к воздействующему фактору, восстанавливаться после отмены повреждающего действия, или необратимые изменения в конце концов приведут к гибели клеток.
При обратимом повреждении клетки отвечают рядом функциональных и морфологических изменений. Одним из наиболее общих критериев клеточного повреждения является изменение способности клеток взаимодействовать с различными красителями. Нормальные клетки поглощают растворенные в воде красители и откладывают их в виде гранул в цитоплазме; ядро при этом не прокрашивается. Поврежденные клетки ( от нагревания, изменения давления, рН среды, воздействия денатурирующего агента) теряют эту способность и краска диффузно распределяется не только в цитоплазме, но и в ядре. В случае обратимого характера повреждений , при условии отмены действия внешнего фактора, гранулообразование в цитоплазме клеток восстанавливается.
Другим характерным признаком повреждения клеток является падение дыхательных процессов в клетке, при этом значительно падает окислительной фосфорилирование, необходимое для синтеза АТФ. Для поврежденных клеток характерно усиление гликолитических процессов ( закисление ), падение количества АТФ и активация протеолиза ( денатурация белков ). Вся совокупность неспецифических обратимых изменений в клетках, возникающих под действием различных агентов, называется “ паранекрозом”. В этой начальной и обратимой стадии изменений процессы клеточной ассимиляции и диссимиляции существенно не изменены.
Однако, в случае необратимого повреждения клеточного метаболизма развертываются события, затрагивающие не только цитоплазму, но и ядерный аппарат. Наиболее значительным проявлением необратимых изменений в поврежденной клетке служит конденсация ( уплотнение, аггрегация) хроматина, падение ядерных синтетических процессов. При гибели клетки хроматин образует грубые сгустки внутри ядра ( пикноз), а само ядро распадается на части или даже растворяется ( кариорексис).
В поврежденных клетках резко снижается митотическая активность, задержки клеток на разных стадиях митоза. Нарушается проницаемость клеточных мембран, в результате происходит вакуолизация мембранных органелл клетки. При этом, происходит интенсивное накопление в клетке каких-то отдельных продуктов нарушенного метаболизма. В общей патологии такие изменения в структуре клеток называются дистрофиями. Так, например, при жировой дистрофии в клетках накапливаются жировые включения, при углеводной –гликогена, при белковой – отложение белковых гранул, пигментов и т.д. Конечным этапом необратимых изменений в клетках является их гибель, на уровне ткани и органа это проявляется в виденекроза, или отмирания ткани ( органа).
Особой формой нарушений регуляции метаболизма клеток является нарушение клеточной дифференцировки, приводящее чаще всего к развитию опухолевого процесса. Опухолевым клеткам присуще неконтролируемое размножение, автономность поведения в организме и нарушение межклеточных взаимодействий. Все эти свойства опухолевых клеток сохраняются от поколения к поколению, т.е. являются наследуемыми. В связи с этими особенностями, раковые клетки считают мутантами с точки зрения неподчиненности их поведения регулирующим воздействиям организма.
Здесь важную роль играют молекулярные процессы поддержания и регуляции ионного гомеостаза клеток в норме и при злокачественной трансформации. Понятие ионный гомеостаз клетки( ИГК) включает систему регуляции активности ионов, обеспечивающих нормальную внутриклеточную среду , и воды. К числу наиболее важных для жизнедеятельности клетки ионов относятся К+ ,Na+,Ca2+,H+,PO2-, и ионы таких энергоемких молекул, как АТФ и АДФ ( аденозиндифосфат). ИГК может управлять поведением клетки, меняя прежде всего количественные соотношения между основными системами клетки – к примеру, интенсивность синтеза белка и РНК, масса цитоплазмы и ядра и т.д. В случае нарушения работы ИГК в клетках изменяются прежде всего молекулярные механизмы их взаимодействий друг с другом, в результате чего опухолевые клетки и приобретают автономность поведения и низкую чувствительность к регулирующим воздействиям.( Маленков А.Г., 1976 г.).
Гибель клеток.
Различают две формы гибели клеток – некрозиапоптоз.
Некрозвызывается преимущественно различными внешними факторами, которые прямо или опосредованно влияют на проницаемость мембран и клеточный метаболизм. Во всех случаях происходит цепь морфо-функциональных нарушений, приводящая в конце концов к растворению клетки – лизису.
Итак, некроз –это форма гибели клеток, характеризующаяся:
функционально – необратимым прекращением их жизнедеятельности,
морфологически –нарушением целостности мембран, изменениями в ядре( пикноз, кариорексис, лизис), цитоплазме ( отек), разрушением клеток, воспалительной реакцией,
биохимически – нарушением выработки энергии, коагуляцией и гидролитическим расщеплением белков, нуклеиновых кислот, липидов,
генетически – потерей генетической информации.( Лушников Е.Ф.,Абросимов А.Ю., 2001).
Апоптоз– это процесс клеточной гибели, который может происходить и без первичного нарушения клеточного метаболизма. Чаще апоптоз называют ещепрограммированной клеточной гибелью; имеется в виду тот факт, что эта форма гибели клеток действительно развивается по программе, заложенной в генетическом аппарате клеток. При этом, в результате действия на клетку различных стимулов в ядре происходит активация определенных генов, стимулирующих самоуничтожение клетки. Программа самоуничтожения может реализовываться под действием некоторых сигнальных молекул самого организма – например, гормонов или белковых молекул. Так, размножающиеся в огромных количествах в вилочковой железе ( тимусе) лимфоциты почти все погибают в течение нескольких часов не покидая вилочковой железы и в кровяном русле под действием глюкокортикоидов – гормонов коры надпочечников. Смысл этого феномена совершенно неясен, но тем не менее он происходит именно по пути апоптоза.
К активации генов самоуничтожения может привести прекращение какого-то регулирующего сигнала. Например, после удаления семенников полностью погибают клетки предстательной железы у мужчин.
Гибель клеток как бы без видимой причины встречается часто при нормальном эмбриональном развитии организма. Например, клетки хвоста головастика погибают в результате активации гормонов щитовидной железы на определенной стадии развития головастика. Во взрослом организме апоптозу подвергаются клетки молочной железы при ее инволюции ( обратном развитии).
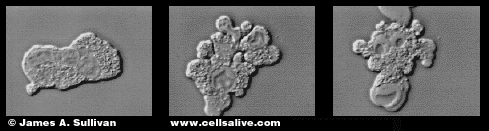
Фиг. Стадии развития апоптоза.Фото слева – нормальная клетка;
В центре – начало фрагментации цитоплазмы; справа – конечный этап
фрагментации цитоплазмы и ядра.
Процесс апоптоза существенно отличается от некроза. Эта форма клеточной гибели характеризуется:
функционально – необратимым прекращением жизнедеятельности клеток,
морфологически – потерей микроворсинок и межклеточных контактов, конденсацией хроматина и цитоплазмы, сморщиванием клеток, образованием пузырьков из плазматических мембран, фрагментацией клеток и образованием микроядер,
биохимически – гидролизом белков цитоплазмы и распадом ДНК,
генетически – структурно-функциональной перестройкой генетического аппарата, завершающейся поглощением клеточных фрагментов макрофагами тканей без воспалительной реакции.( Лушников Е.Ф.,Абросимов А.Ю., 2001).
Сущность апоптоза , его место в норме и при патологии.
В многоклеточном организме гибель клеток происходит постоянно, но в разных тканях и органах она наблюдается при определенных условиях. Биологическая сущность гибели клеток неоднозначна. Самое главное и существенное, это то, что апоптоз имеет разное значение для нормальной жизни позвоночных животных и беспозвоночных животных ( а теперь стало известно, что и для растительного мира):
в эмбриогенезе апоптоз является элементом развития и связан с формированием тканей и органов,
при самостоятельном существовании организма апоптоз является составной частью метаморфоза ( у насекомых и амфибий),
у животных процессы гистогенеза и органогенеза протекают с участием апоптоза,
в зрелом организме апоптоз является частью гомеостатического механизма смены клеток,
при старении апоптоз отражает естественную гибель клеток.
Репродуктивная гибель клеток.
В связи со стадиями клеточного цикла существует две формы гибели клеток от внешнего воздействия. Одна из них – репродуктивная гибель, касающаяся только делящихся популяций клеток. Эта форма гибели клеток была обнаружена в радиобиологических экспериментах на лабораторных животных в 70-е годы.
У облученных животных при цитологических исследованиях была обнаружена потеря некоторыми клетками способности к делению после прохождения нескольких клеточных циклов, с последующей гибелью. Причем, такая форма гибели наблюдается и у клеток в культуре “ин витро”.
Интерфазная гибель клеток.
Другой феномен, обнаруженный также в радиобиологических работах- немедленная гибель клеток после облучения ,без деления – в интерфазе. Эта форма поражения клеток характерна для лучевых последствий в системе кроветворения в костном мозге, в иммунной и нервной системах . Согласно современным представлениям, интерфазная гибель клеток происходит по пути апоптоза.
Лекция 7. Общая эмбриология. Основные понятия и феномены.
Эмбриология( от греч.embryon– зародыш) – наука о закономерностях развития зародышей. Медицинская эмбриология (Э) изучает закономерности развития зародыша человека, причины развития уродств и другие отклонения от нормы, возможные пути и методы воздействия на эмбриогенез( процесс формирования зародыша).
В настоящее время медицинские технологии позволяют во многих случаях бороться с бесплодием, осуществлять рождение “ пробирочных” детей, проводить трансплантации зародышевых органов и тканей. Сейчас уже разработаны методы культивирования ин витро яйцеклеток, вне- организменное оплодотворение и имплантация зародыша в матку.
Процесс эмбрионального развития человека претерпел длительную эволюцию и в значительной степени отражает стадии развития других представителей животного мира. Поэтому, ранние стадии развития зародыша человека сходны с аналогичными стадиями эмбриогенеза менее организованных хордовых животных.
Репродуктивная система человека включает несколько взаимосвязанных этапов развития мужских и женских половых клеток, слияния этих клеток при оплодотворении и формирование нового организма из зародыша. Эти этапы следующие:
Прогенез –развитие и созревание половых клеток-яйцеклеток и сперматозоидов. В результате прогенеза в зрелых половых клетках возникает гаплоидный ( одинарный) набор хромосом, формируются структуры, способствующие их взаимному слиянию( оплодотворению) и развитию нового организма.
Эмбриогенез-часть онтогенеза человека ( его индивидуального развития), ввключает основные стадии : 1- оплодотворение с образованием зиготы, 2- дробление зиготы и образование бластулы( бластоцисты), 3 –гаструляцию- образование зародышевых листков и комплекса осевых органов, 4- образование зародышевых и внезародышевых тканей и органов ,5- формирование систем органов( системогенез).
Пост-эмбриональный период –функционирование нового индивида после рождения вне организма матери, включая и продолжение формирования органов и тканей после рождения.
Прогенез.
Половые клетки( гаметы) , в отличие от соматических клеток,содержат не двойной, а одинарный набор хромосом. У человека этот двойной набор составляет 46 хромосом, следовательно в гаметах содержится по 23 хромосомы.
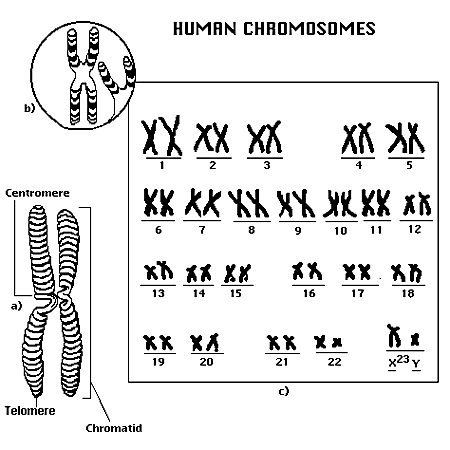
Фиг.Диплоидный хромосомный набор( 46 хромосом) в соматических клетках у человека
Все хромосомы в гаметах называются аутосомами-их в гамете 22, за исключением одной – двадцать третьей , которая называетсяполовой хромосомой. Вмужскихполовых клетках половина гамет( сперматозоидов)(С)содержит половую хромосому с женским генетическим материалом ( Х- хромосома), и половина – хромосому с мужским генетическим материалом –Y- хромосома. В женских гаметах( яйцеклетках) ( Я) все половые хромосомы являются Х-несущими. Характерной особенностью гамет является невысокий уровень метаболизма в них . Кроме того, зрелые гаметы утрачивают способность делиться.
Мужские половые клетки – сперматозоиды –образуются в огромном количестве у большинства высших животных.У млекопитающих и человека сперматозоиды образуются и созревают в течение всего активного полового периода в генеративных органах – яичках , из первичных половых клеток сперматогенного эпителия извитых канальцев. В процессе митотического деления, часть сперматогоний ( следующая стадия дифференцировки первичных половых клеток) вступает в процесс дифференцировки до состояния зрелых сперматозоидов, а часть не дифференцируется и продолжает митотически делиться, тем самым поддерживая пул стволовых( исходных) клеток .(Далее нужно кратко описать мейоз.)