

Биоэнергетические процессы в клетке (Атомная энергетика клетки).
Жизнедеятельность клеток требует энергетических затрат. Живые системы( организмы) получают ее из внешних источников, например, от Солнца( фототрофы, каковыми являются растения, некоторые виды простейших и микроорганизмы), или производят ее сами( аэробные аутотрофы) в результате окисления различных веществ(субстратов).
В обоих случаях клетки синтезируют универсальную высокоэнергетичную молекулу АТФ( аденозинтрифосфорную кислоту),при разрушении которой выделяется энергия. Эта энергия расходуется для выполнения всех видов функций- активного транспорта веществ, синтетических процессов, механической работы и т.д.
Сама по себе молекула АТФ достаточно проста и представляет собой нуклеотид, состоящий из аденина, сахара рибозы и трех остатков фосфорной кислоты.( Рис). Молекулярная масса АТФ невелика и составляет 500 дальтон. АТФ является универсальным переносчиком и хранителем энергии в клетке, которая заключена в высокоэнергетичных связях между тремя остатками фосфорной кислоты.
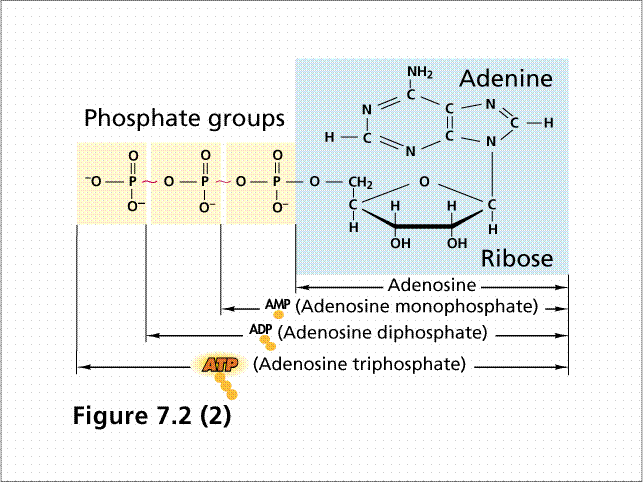
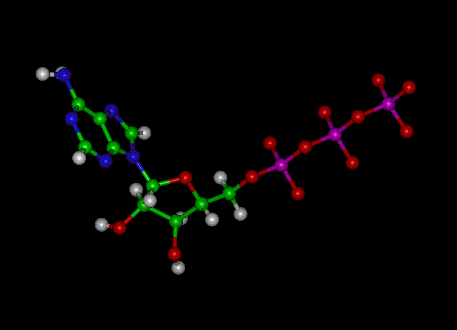
структурная формула пространственная формула
Рис 37. Аденозин-трифосфорная кислота ( АТФ)
Цвета для обозначения молекул(пространственная формула):белый –водород,красный – кислород, зеленый –углерод, голубой –азот,темно-красный - фосфор
Отщепление одного лишь остатка фосфорной кислоты от молекулы АТФ сопровождается высвобождением значительной порции энергии – около 7,3 ккал.
Как же происходит процесс запасания энергии в виде АТФ? Рассмотрим это на примере окисления( сгорания) глюкозы – распространенного источника энергии для перевода в энергию химических связей АТФ.
НОСН2 С
О Н
Н Н С
С ОН
ОН Н С
С ОН
Н
ОН Рис
38. Структурная формула глюкозы
( содержание в крови человека- 100 мг%)








Окисление одного моля глюкозы( 180 г) сопровожда-
ется выделением около 690 ккал свободной энергии.
 С6Н12О6+ 6О2 6СО2+6Н2О + Е( около 690 ккал)
С6Н12О6+ 6О2 6СО2+6Н2О + Е( около 690 ккал)
В живой клетке это огромное количество энергии высвобождается не сразу, а постепенно в виде ступенчатого процесса и регулируется целым рядом окислительных ферментов. При этом, высвобождаемая энергия переходит не в тепловую энергию, как при горении, а запасается в виде химических связей в молекуле АТФ( макроэргические связи) в процессе синтеза АТФ из АДФ и неорганического фосфата. Этот процесс можно сравнить с работой аккумулятора, который заряжается от различных генераторов и может обеспечивать энергией множество машин и аппаратов. В клетке роль унифицированного аккумулятора выполняет система аденозин-ди и три-фосфорных кислот. Зарядка аденилового аккумулятора состоит в соединении АДФ с неорганическим фосфатом ( реакция фосфорилирования) и образовании АТФ:
 АДФ + ФнеоргАТФ +
Н2О
АДФ + ФнеоргАТФ +
Н2О
Для образования всего 1 молекулы АТФ требуется затрата энергии извне в количестве 7,3 ккал. И наоборот, при гидролизе АТФ( разрядке аккумулятора) это же количество энергии выделяется. Оплата этого энергетического эквивалента, называемого в биоэнергетике “ квантом биологической энергии “ происходит из внешних ресурсов – то есть за счет пищевых веществ. Роль АТФ в жизнедеятельности клетки может быть представлена так:
АТФ
Энергети-
Система Система Функции ческие
ре- аккумуляции использова-
клетки сурсы
энергии ния энергии
АДФ
+ Фн
Рис.39
Общий план энергетики клетки














Синтез молекул АТФ происходит не только за счет расщепления углеводов( глюкозы), но и белков( аминокислот) и жиров( жирных кислот). Общая схема каскадов биохимических реакций такова( Рис ).
1.Начальные этапы окисления происходят в цитоплазме клеток и не требуют участия кислорода. Эта форма окисления называется анаэробным окислением , или проще – гликолизом.Основной субстрат при анаэробном окислении –гексозы, преимущественно глюкоза. В процессе гликолиза происходит неполное окисление субстрата: глюкоза распадается до триоз ( две молекулы пировиноградной кислоты). При этом, для осуществления реакции в клетке затрачивается две молекулы АТФ, но и синтезируется 4 молекулы АТФ. То есть, методом гликолиза клетка “ зарабатывает” всего две молекулы АТФ при окислении 1 молекулы глюкозы. С точки зрения эффективности энергетики это
маловыгодный процесс.При гликолизе высвобождается всего 5% энергии химических связей молекулы глюкозы.
 С6
Н12 О6+ 2Фнеорг+2АДФ
2 С3Н4О3+2АТФ + 2Н2О
С6
Н12 О6+ 2Фнеорг+2АДФ
2 С3Н4О3+2АТФ + 2Н2О
Глюкоза пируват
Несмотря на низкую эффективность, гликолиз широко используется в природе: у микроорганизмов, у паразитических кишечных анаэробных простейших, у многих опухолевых клеток.Эритроциты млекопитающих, например, получают энергию за счет гликолиза и у них отсутствуют митохондрии.
2. Образовавшиеся в процессе гликолиза триозы ( в основном-пировиноградная кислота, пируват) использу-
ются для дальнейшего более эффективного окисления, но уже в органеллах клетки – митохондриях. При этом, высвобождается энергия расщепления всех химических связей, что приводит к синтезу большого количества АТФ и потреблению кислорода.
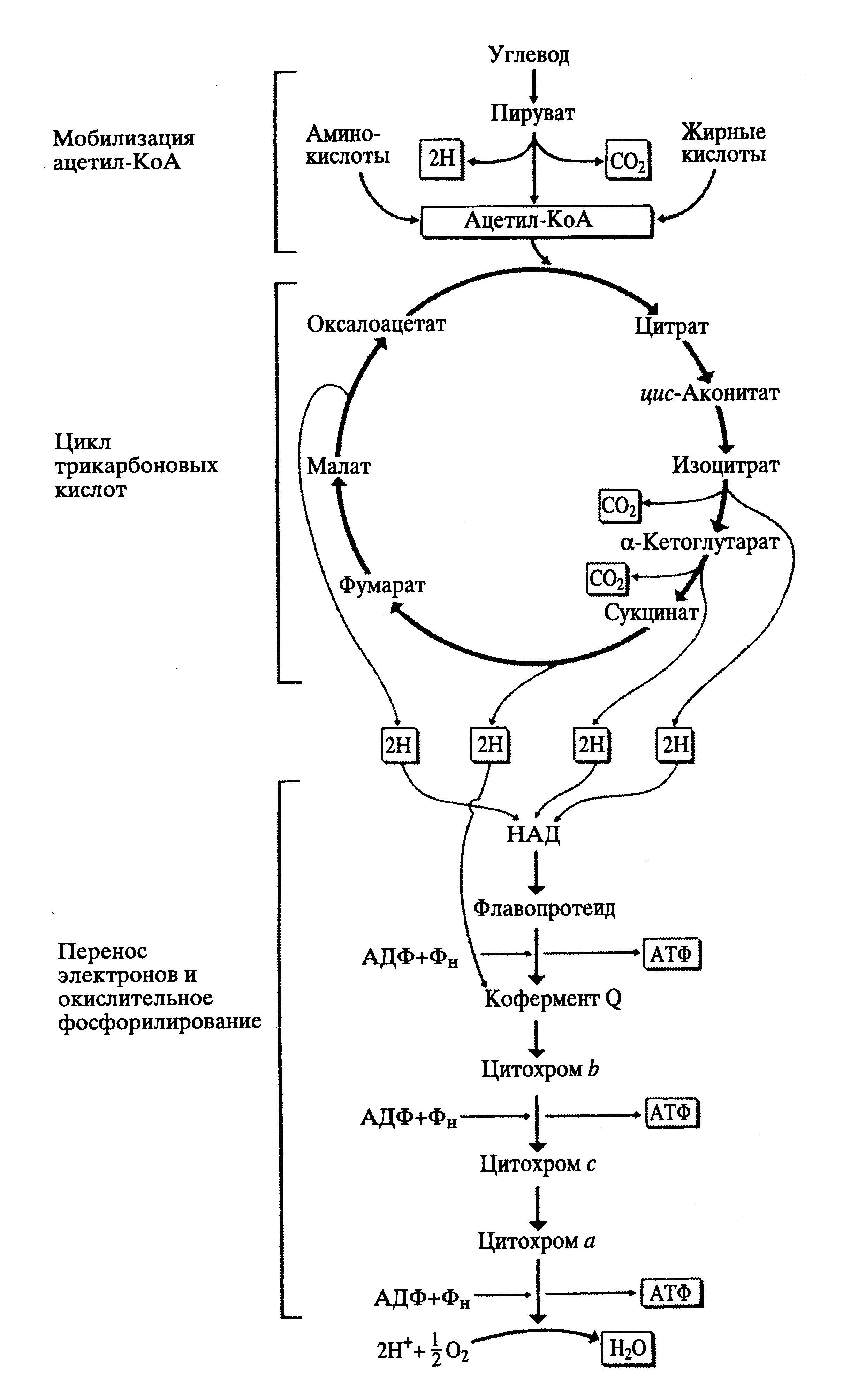
Рис.40 Схема цикла Кребса( трикарбоновых кислот) и окислительного фосфорилирования( дыхательной цепи)
Эти процессы связаны с окислительным циклом трикарбоновых кислот (синонимы:циклом Кребса, циклом лимонной кислоты) и с цепью переноса электронов с одного фермента на другой ( дыхательная цепь), когда из АДФ образуется АТФ путем присоединения одного остатка фосфорной кислоты( окислительное фосфорилирование).
Понятием “окислительное фосфорилирование“ определяют синтез АТФ из АДФ и фосфата за счет энергии окисления субстратов ( питательных веществ).
Под окислениемпонимают отнятие электронов от вещества, соответственно – восстановление – присоединение электронов .
Какова роль окислительного фосфорилирования у человека? Представление об этом может дать следующий грубый расчет:
Взрослый человек при сидячей работе потребляет в день около 2800 ккал энергии с пищей. Для того, чтобы такое количество энергии было получено методом гидролиза АТФ, потребуется 2800/7,3 = 384 моль АТФ, или 190 кг АТФ. Тогда как известно, что в организме человека содержится около 50 г АТФ. Поэтому ясно, что для удовлетворения потребности в энергии в организме эти 50 г АТФ должны тысячи раз расщепиться и заново синтезироваться. Кроме того, сама скорость обновления АТФ в организме меняется в зависимости от физиологического состояния – минимальная во время сна и максимальная – при мышечной работе. А это означает, что окислительное фосфорилирование – не просто непрерывный процесс, но и регулируемый в широких пределах.
Суть окислительного фосфорилирования заключается в сопряжении двух процессов, когда окислительная реакция с привлечением энергии извне( экзэргическая реакция) увлекает за собой другую, эндэргическую реакцию фосфорилирования АДФ неорганическим фосфатом:
 Ав АДФ + Фн
Ав АДФ + Фн
окисление фосфорилирование

 Ао АТФ
Ао АТФ
Здесь Ав–восстановленная форма вещества, подвергающегося фосфорилирующему окислению,
Ао – окисленная форма вещества.
В цикле Кребса образовавшийся в результате гликолиза пируват ( СН3СОСООН) окисляется до ацетата и соединяется с коферментом А, образуя ацетил-коА. После нескольких этапов окисления образуется шестиуглеродное соединение лимонная кислота( цитрат), также окисляющееся до оксал-ацетата; затем цикл повторяется(Cхема цикла трикарб. Кислот). При этом окислении выделяются две молекулы СО2и электроны, которые переносятся на акцепторные( воспринимающие) молекулы ко-ферментов( НАД – никотинамиддинуклеотид) и затем вовлекаются в цепь переноса электронов с одного субстрата( фермента) на другой.
При полном окислении одного моля глюкозы до СО2и Н2О в цикле гликолиза и трикарбоновых кислот образуется 38 молекул АТФ с энергией химических связей 324 ккал, а общий выход свободной энергии этого превращения , как отмечалось ранее, составляет 680 ккал. Эффективность выхода запасенной энергии в АТФ составляет 48%( 324/680 х100%= 48%).
Cуммарное уравнение окисления глюкозы в цикле Кребса и гликолитическом цикле:
 C6Н12О6 +6О2+36 АДФ +Фн 6СО2+36АТФ +
42Н2О
C6Н12О6 +6О2+36 АДФ +Фн 6СО2+36АТФ +
42Н2О
3. Освободившиеся в результате окисления в цикле Кребса электроны соединяются с ко-ферментом и транспортируются в цепь переноса электронов( дыхательную цепь) с одного фермента на другой, где в процессе переноса и происходит сопряжение( трансформация энергии электронов в энергию химических связей) с синтезом молекул АТФ.
Существует три участка дыхательной цепи, в которых энергия процесса окисления-восстановления трансформируется в энергию связей молекул в АТФ. Эти участки называются пунктами фосфорилирования:
1.Участок переноса электронов от НАД-Н к флавопротеиду, синтезируется 10 молекул АТФ за счет энергии окисления одной молекулы глюкозы,
2.Перенос электронов на участке от цитохрома б к цитохрому с1, фосфорилируется 12 молекул АТФ на одну молекулу глюкозы,
3. Перенос электронов на участке цитохром с – молекулярный кислород, синтезируется 12 молекул АТФ.
Итого, на этапе дыхательной цепи происходит синтез( фосфорилирование) 34 молекул АТФ. А общий выход АТФ в процессе аэробного окисления одной молекулы глюкозы составляет 40 единиц.
Таблица 1
Энергетика окисления глюкозы
|
Механизм накопления энергии |
Выход АТФ на одну молекулу глюкозы |
|
Гликолиз
Цикл Кребса( трикарбо- новых кислот)
Дыхательная цепь
|
4
2
34
Итого: 40 С вычетом энергозатрат на запуск гликолиза 38 АТФ
|
На каждую пару электронов, передающихся по цепи от НАД –Н+к кислороду, синтезируется три молекулы АТФ
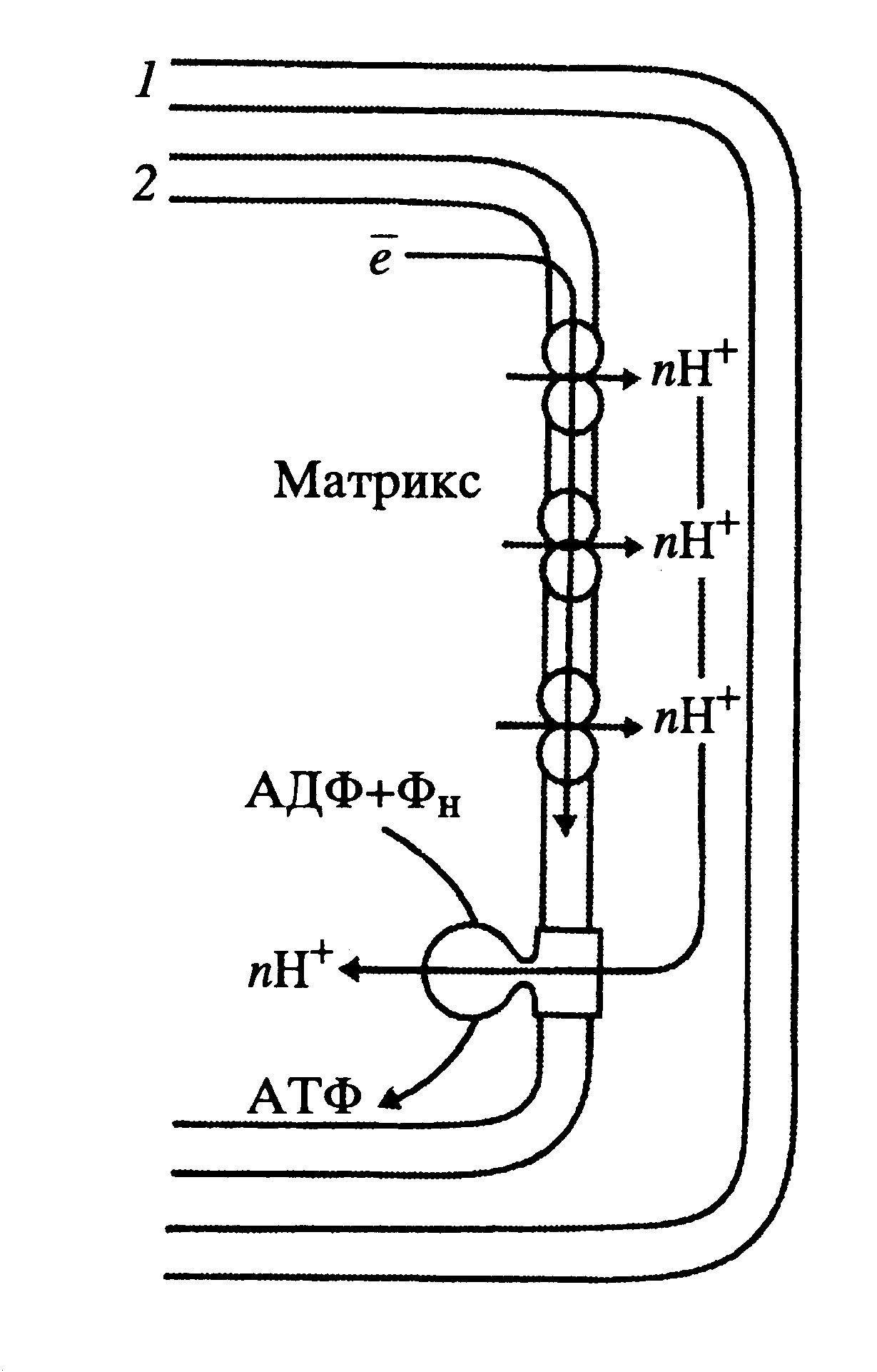
Дыхательная цепь представляет собой ряд белковых комплексов, встроенных во внутреннюю мембрану митохондрий( Рис 41 ).
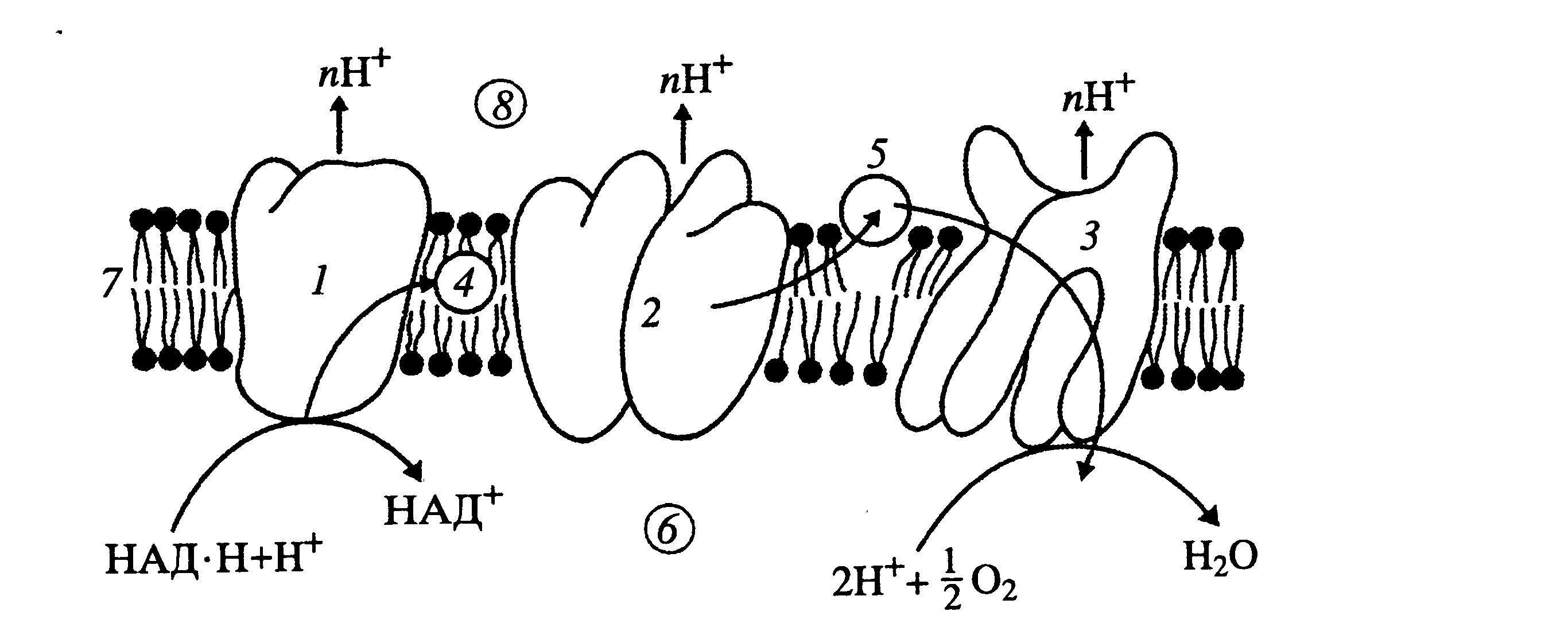
Рис.41 Схема расположения ферментов дыхательной цепи во внутренней мембране митохондрий:
1-НАД-Н-дегидрогеназный комплекс, с1-комплекс, 3-цитохром-оксидазный комплекс, 4-убихинон, 5-цито-
хром-с, 6-матрикс митохондрии, внутренняя мембрана митохондрии,8- межмембранное пространство.
Итак, полное окисление исходного субстрата завершается высвобождением свободной энергии, значительная часть которой ( до 50%) расходуется на синтез молекул АТФ, образованием СО2и воды.Другая половина свободной энергии окисления субстратов идет на следующие нужды клетки:
1. Для биосинтеза макромолекул( белков, жиров, углеводов),
2. Для процессов движения и сокращения,
3. Для активного транспорта веществ через мембраны,
4.Для обеспечения передачи генетической информации.
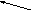
3
Рис.42 Общая схема процесса окислительного фосфорилирования в митохондриях.
1- наружная мембрана митохондрии, 2- внутренняя мембрана, 3- встроенный во внутреннюю мембрану фермент АТФ-синтетаза.
Синтез молекул АТФ
Синтез АТФ происходит во внутренней мембране митохондрий, смотрящей в матрикс( Рис 42 выше).В нее встроены специализировавнные белки-ферменты, занимающиеся исключительно синтезом АТФ из АДФ и неорганического фосфата Фн-АТФ-синтетазы( АТФ-С). В электронном микроскопе эти ферменты имеют весьма характерный вид, за что и были названы “грибовидными тельцами”( Рис). Эти структуры сплошь выстилают внутреннюю поверхность мембраны митохондрии,направленную в матрикс.По образному
выражению известного исследователя биоэнергетики проф. Тихонова А.Н.,АТФ-С является “самым миниатюрным и совершенным мотором в природе”.
Рис.43
Локализация АТФ-синтетаз
в мембране мито- хондрий(
клетки животных) и хлоропластов( клетки
растений).
Голубые
участки -области с повышенной концентрацией
Н+(
кислотная зона),оранжевые участки
–области с низкой концентрацией Н+. Внизу:
перенос ионов водорода Н+
через мембрану при синтезе(а) и
гидролизе(б) АТФ
Эффективность работы этого фермента такова, что одна молекула способна осуществить 200 циклов ферментативной активации в секунду, при этом синтезируется 600 молекул АТФ.
Интересная подробность работы этого мотора в том, что он содержит вращающиеся детали и состоит из роторной части и статора, причем, вращение ротора происходит против часовой стрелки.( Рис. 44)
Мембранная часть АТФ-С, или фактор сопряжения F0,представляет собой гидрофобный белковый комплекс. Второй фрагмент АТФ-С – фактор сопряженияF1- выступает из мембраны в виде грибовидного образования. В митохондриях животных клеток АТФ-С встроена во внутреннюю мембрану, а комплексF1обращен в сторону матрикса.
Образование АТФ из АДФ и Фн происходит в каталитических центрах фактора сопряжения F1. Этот белок можно легко выделить из мембраны митохондрий, при этом он сохраняет способность гидролизовать молекулу АТФ, но теряет способность синтезировать АТФ. Способность синтезировать АТФ – это свойство единого комплексаF0F1в мембране митохондрии( рис1 а)Это связано с тем, что синтез АТФ в помошью АТФ-С сопряжен с транспортом через нее протонов Н+в направлении отF0rF1( рис 1 а). Движущей силой для работы АТФ-С является протонный потенциал, создаваемый дыхательной цепью транспорта электронов е-.
АТФ-С – это обратимая молекулярная машина, катализирующая как синтез, так и гидролиз АТФ. В режиме синтеза АТФ работа фермента осуществляется за счет энергии протонов Н+, переносимых под действием разности протонных потенциалов. В то же время, АТФ-С работает и как протонная помпа – за счет энергии гидролиза АТФ она перекачивает протоны из области с низким протонным потенциалом в область с высоким потенциалом ( рис 1б). Сейчас уже известно, что каталитическая активность АТФ-С непосредственно связана с вращением ее роторной части. Было показано, что молекулаF1 вращает роторный фрагмент дискретными скачками с шагом в 1200. Один оборот на 1200сопровождается гидролизом одной молекулы АТФ.
Замечательным качеством вращающегося мотора АТФ-С является его исключительно высокая КПД. Было показано, что работа, которую совершает мотор при повороте роторной части на 1200, почти точно совпадает с величиной энергии, запасенной в молекуле АТФ, т.е. КПД мотора близок к 100%.
В таблице приведены сравнительные характеристики нескольких типов молекулярных моторов, работающих в живых клетках. Среди них АТФ-С выделяется своими наилучшими свойствами. По эффективности работы и развиваемой ею силе она значительно превосходит все известные в природе молекулярные моторы и уж конечно – все созданные человеком.
Таблица 2 Сравнительные характеристики молекулярных моторов клеток( по: Kinoshitaetal, 1998).
|
Мотор/ Путь |
Шаг |
Макс.сила |
Макс. КПД |
Способ движения |
|
Миозин/ актин
Кинезин/ микротру- бочка
РНКполи- Мераза
F1-комплекс АТФ-С |
Перемен Ный
Смещение На 8 нм
Смещение на 0.34 нм
Поворот На 1200 |
3-5 пН
5 пН
14 пН
40 пН .нм |
Около 20%
Ок 50%
Ок. 20%
Ок 100% |
Шаги, прыжки Бег Шаги
Ползание
вращение |
Молекула F1комплекса АТФ-С примерно в 10 раз сильнее акто-миозинового комплекса – молекулярной машины, специализирующейся на выполнении механической работы. Таким образом, за многие миллионы лет эволюции до того, как появился человек,придумавший колесо, преимущества вращательного движения были уже реализованы природой на молекулярном уровне.
Объем работы, которую производит АТФ-С, поражает грандиозностью. Общая масса молекул АТФ синтезируемых в организме взрослого человека за сутки cоставляет около 100 кг. В этом нет ничего удивительного, поскольку в организме идут многочисленные
биохимические процессы с использованием АТФ. Поэтому, чтобы организм мог жить, его АТФ-С должны постоянно крутиться, своевременно восполняя запасы АТФ.
Яркий пример молекулярных электромоторов – работа жгутиков бактерий. Бактерии плавают со средней скоростью 25 мкм/с , а некоторые из них – со скоростью более 100 мкм/с. Это означает, что за одну секунду бактерия перемещается на расстояние в 10 и более раз большее, чем собственные размеры. Если бы пловец преодолевал за одну секунду расстояние, в десять раз больше его собственного роста б , то 100-метровую дорожку он проплывал бы за 5 секунд!
Скорость вращения электромоторов бактерий колеблется от 50-100 об/ сек до 1000 об/ сек, при этом они очень экономичны и расходуют не более 1% энергетических ресурсов клетки.
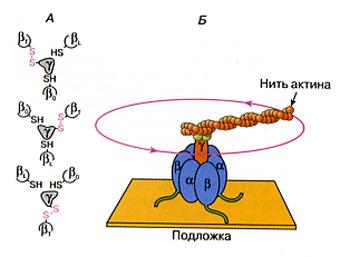
Рис 44. Схема вращения роторной субъединицы АТФ-синтетазы.
Таким образом, во внутренней мембране митохондрий локализованы как ферменты дыхательной цепи, так и синтеза АТФ.
Помимо синтеза АТФ, выделяющаяся при транспортировке электронов энергия запасается еще и в виде градиента протонов на мембране митохондрий.При этом, между наружной и внутренней мембранами возникает повышенная концентрация ионов Н+( протонов). Возникший протонный градиент из матрикса в межмембранное пространство служит движущей силой при синтезе АТФ( Рис.42). По существу,внутренняя мембрана митохондрий с встроенными АТФ-синтетазами является совершенной электростанцией на протонах, поставляющей с высокой эффективностью энергию для жизнедеятельности клетки.
При достижении определенной разности потенциалов ( 220 мВ) на мембране,АТФ-синтетаза начинает транспортировать протоны обратно в матрикс; при этом происходит превращение энергии протонов в энергию синтеза химических связей АТФ. Так осуществляется сопряжение окислительных процессов с синтетически-
ми в процессе фосфорилирования АДФ до АТФ.
Энергетика окислительного фосфорилирования
жиров
Еще более эффективным оказывается синтез АТФ при окислении жирных кислот и липидов. При полном окислении одной молекулы жирной кислоты, например, пальмитиновой, образуется 130 молекул АТФ. Изменение свободной энергии окисления кислоты составляет ∆G= -2340 ккал, а аккумулированная в АТФ энергия при этом составляет около 1170 ккал.
Энергетика окислительного расщепления аминокислот
Большую часть метаболической энергии, вырабатываемой в тканях, обеспечивают процессы окисления углеводов и особенно жиров; у взрослого человека до 90% всей потребности в энергии покрывается из этих двух источников. Остальную энергию ( в зависимости от рациона от 10 до 15%) поставляет процесс окисления аминокислот( рис цикла Кребса ).
Было подсчитано, что клетка млекопитающих содержит в среднем около 1 миллиона ( 106) молекул АТФ. В пересчете на все клетки тела человека ( 1016 –1017 ) это составляет 1023 молекул АТФ. Суммарная энергия, заключенная в этой массе АТФ, может достигать значений 1024 ккал! ( 1 Дж= 2,39х 10-4ккал). У человека весом 70 кг общее количество АТФ составляет 50 г, большая часть которого ежедневно расходуется и вновь синтезируется.