

Глава 5
РЕГУЛЯЦИЯ ДЕЯТЕЛЬНОСТИ
МЕХАНОРЕЦЕПТОРОВ
Способность животных изменять поток сенсорной информации, поступающей в организм, была замечена исследователями очень давно. Однако первоначально эта способность рассматривалась в основном как некоторый комплекс реакций, определяющих ту или иную ориентацию рецепторных поверхностей по отношению к источнику сигналов, а также изменяющих характеристики сигналов при их прохождении через вспомогательные структуры сенсорных образований к собственно воспринимающим элементам. Изменение положения тела или отдельных его частей (например, головы, глаза или ушных раковин), сокращение или расслабление мышц среднего уха, расширение или сужение зрачка и т. д. — все это типичные примеры такого рода реакций.
Наряду с этими наиболее явными формами регулирования сенсорного потока, как показали дальнейшие исследования, у организмов имеются и другие многочисленные способы воздействия на сигналы, поступающие в центральную нервную систему от рецепторных приборов. Можно утверждать, что в наиболее совершенных сенсорных аппаратах практически каждый их элемент, каждое звено переработки информации находится под эффективным контролем нервной системы. И то, что структура сенсорной информации претерпевает значительные изменения на своем пути к высшим нервным центрам, во многом определяется эфферентным регулирующим влиянием. Эфферентная регуляция деятельности рецепторов изучена в настоящее время весьма неравномерно. Если в отношении воспринимающих элементов органов чувств она исследована еще весьма недостаточно, то по отношению к мышечным рецепторам, играющим огромную роль в осуществлении координированных двигательных актов; эта проблема разработана достаточно подробно. При этом наиболее изученными оказались мышечные веретена млекопитающих и рецепторы растяжения ракообразных. Лишь в самое последнее время ряд важных фактов удалось получить и в отношении волосковых механорецепторов.
В данном разделе будут изложены данные, касающиеся регулирования деятельности основных видов механорецепторных приборов в коже, мышцах, органах чувств. Основное внимание уделяется рассмотрению фактов по эфферентной регуляции мышечных веретен млекопитающих, рецепторов растяжения у ракообразных, волосковых механорецепторов органов чувств позвоночных.
Как показывают существующие в литературе данные, имеются важные особенности в эфферентном регулировании деятельности различных механорецепторов. Анализ этих особенностей, представленный в заключении главы, показывает, что основное различие в характере регулирования наблюдается между первичными и вторичными механорецепторными приборами позвоночных.
Регуляция деятельности первичных механорецепторов у позвоночных
Регуляция активности рецепторов кожи
Можно отметить несколько способов регулирования нервной системой афферентного потока сигналов, поступающих от тканевых механорецепторов. Во-первых, это влияния, изменяющие механическое состояние тканей (например, изменения растяжения кожных покровов, достигаемые движениями). Во-вторых, это влияния, сказывающиеся на состоянии трофики ткани (спазм или расширение сосудов данной области, изменения метаболизма). В-третьих, это прямые влияния нервной системы на свойства отдельных рецепторов или групп рецепторных элементов. Далее, нервная система оказывает суммарное воздействие на тканевые механорецепторные структуры с помощью различных гуморальных факторов. Наконец, тканевые механорецепторы могут и сами взаимодействовать между собой на основе механизма аксон-рефлекса, а также с помощью гуморальных факторов, высвобождаемых из нервных и тканевых элементов в ходе раздражения сенсорных структур.
Действие симпатических волокон и катехоламинов. Деятельность механорецепторов кожи регулируется в основном с помощью симпатической нервной системы. Изменение кожной чувствительности после экстирпации шейного симпатического ганглия у кошки отмечалось еще Клодом Бернаром (Bernard, 1851). Однако далее этот вопрос длительное время специально не рассматривался, пока в связи с развитием учения об адаптационно-трофической функции симпатической нервной системы и ее влиянии на органы чувств он не стал предметом пристального внимания в школе Л. А. Орбели (Тонких, 1925; Юрьева, 1927; Кунстман, 1928; Сперанская-Степанова, 1930; Орбели, 1932; Загорулько, Лебединский, 1935). В этих исследованиях было твердо установлено существование влияния симпатической нервной системы на кожную чувствительность.
Как показали в дальнейшем многочисленные исследования (Жирмунская, 1940; Пахомов, Проппер-Гращенков, 1941; Landgren, 1952b; Landgren, Liljestrand, Zotterman, 1952; Landgren, Neil, Zotterman, 1952; Douglas, Gray, 1953; Witzleb, 1953; Diamond, 1955; Loewenstein, 1956a; Paintai, 1959; Leitner, Perl, 1964; Дмитриева, Есаков, 1969; Nilsson, 1972, 1974; Sharma et al., 1972, и др.), раздражение симпатических нервов, а также подведение адреналина, норадреналина и других симпатомиметических аминов оказывают на тканевые (в том числе и кожные) механорецепторы весьма сложное влияние. Это объясняется тем, что адреналин влияет и на все части рецепторного аппарата, и на сосуды, и на различные контрактильные элементы ткани, причем иногда противоположно. Даже в пределах одного и того же звена адреналин обладает сложным фазным действием. Естественно, что результирующий эффект зависит от способа подведения вещества, его дозы и длительности действия (соответственно интенсивности и продолжительности раздражения симпатических нервов).
Наиболее обстоятельное изучение действия адреналина и других сходных веществ на тканевые механорецепторы было проведено в экспериментах на тельцах Пачини брыжейки кошки. Первые исследования этого вопроса (Gammon, Bronk, 1935) показали, что после введения адреналина в сосудистое русло мезентериальные тельца Пачини, возбуждающиеся в такт с пульсовыми толчками, снижают свою активность. В противоположность этому Ильинский (1966а) нашел, что адреналин, инъецированный в мелкий мезентериальный сосуд в направлении группы телец Пачини, приводит к увеличению величины их разрядов, синхронных с пульсовыми колебаниями сосудов. Снижение импульсации от телец Пачини в опытах ГэммонаиБронкабыло обусловлено, по-видимому, уменьшением величины пульсовых толчков сосудов, а не действием адреналина непосредственно на рецепторы.
Изучение действия адреналина на изолированных тельцах Пачини (Loewenstein, Altamirano-Orrego, 1956) показало, что аппликация адреналина или норадреналина (1.0—0.1 мкг) вызывает понижение порога возникновения ПД при механическом раздражении изолированного одиночного рецептора. Порог медленно падал и в течение 2—7 мин. достигал 90—70% исходного значения. Если в начале эксперимента рецептор отвечал одним импульсом на механический толчок пороговой силы, то при максимальном развитии действия адреналина наблюдалось появление нескольких спайков. Аналогичный результат был получен Черепновым (1969).
В самое последнее время вопрос о действии адреналина и норадреналина на тельца Пачини был рассмотрен Акоевым и др. (1974а, 19746) в условиях методики непрерывной внешней перфузии декапсулированного тельца Пачини. Оказалось, что добавление
в нормальный перфузионный раствор небольших концентраций (1 • 10“7—1 • 10-5 г/мл) адреналина и норадреналина приводит к фазным изменениям возбудимости рецептора (рис. 72). Вначале наблюдается незначительное повышение порога возникновения ПД, которое длится 0.5—1 мин. В дальнейшем, по мере пропускания раствора с адреналином порог четко снижался на 10—15%. Это повышение возбудимости длилось обычно 5—7 мин. и затем сменялось резким снижением возбудимости вплоть до наступления полного блока генерации ПД. Скорость развития и продолжитель-
усл.ед
-
L1 . . 11 I . , i
1
Ю
20 30 мин.
имп.
мкб
60-
40
-
20-![]()
![]()

![]()

![]()


5 10 15
мин.
Рис.
72. Изменение порога возбуждения (И),
числа импульсов в разряде (Б)
и
амплитуды рецепторного потенциала
(В)
телец Пачини под влиянием катехоламинов
(пр: Акоевой др., 1974а).![]()


_1 1 I | |
40 80 120 160 200 усл. ед.
А — изменение возбудимости рецептора при действии адреналина (Ь10~7 г/мл). По оси абсцисс — время (мин.); по оси ординат — величина порогового механического раздражения (усл. ед.). Стрелки: первая — переход к перфузии адреналином, вторая — переход к перфузии раствором без адреналина. В — изменение количества импульсов в разряде при действии адреналина (5-10-8 г/мл). По оси абсцисс — время (мин.); по оси ординат — число импульсов в разряде. Стрелка — переход к перфузии адреналином. В — амплитуда РП тельца Пачини, находящегося в растворе без (черные кружки) и с добавлением норадреналина (2 «10-^ г/мл) (белые кружки). По оси абсцисс — величина механического воздействия (усл. ед.);- по оси ординат — амплитуда РП (мкв).
ность наблюдаемых эффектов определялись степенью декапсуля- ции рецепторов. Часто под влиянием адреналина в фазе повышенной возбудимости возникала спонтанная активность рецептора, которая затем исчезала по мере снижения возбудимости. Наряду с изменением порогов менялось и количество импульсов вЪтвете рецептора при действии механического стимула (рис. 72).
Изменение возбудимости рецептора, определяемой по импульсной активности, зависит как от свойств собственно нервного о он- чания (нерегенеративной системы генерации), так и от свойств его волокна (регенеративной системы генерации). Под влиянием адреналина наблюдалось возрастание критического уровня РП и амплитуды ПД.
Исследование изменений РП (Loewenstein, Altamirano-Or- rego, 1956) обнаружило, что под действием адреналина и норадреналина амплитуда и скорость нарастания РП увеличиваются. В ряде случаев возрастала и длительность РП. При удалении катехоламинов и после отмывания рецептора физиологическим раствором наблюдалось восстановление исходной реакции. Более детальные исследования (Черепнов, 1969; Акоев и др., 1974а, 19746) выяснили, что изменения РП под действием адреналина и норадреналина носят фазный характер. Вначале наблюдалось увеличение (на 13—25%) амплитуды и скорости нарастания РП, вслед за которым имело место уменьшение этих параметров. Как видно из рис. 72, в период положительной фазы действия симпатомиметических аминов кривая зависимости амплитуды РП от силы раздражения смещалась вдоль оси ординат, следовательно, в этот период под влиянием адреналина (норадреналина) происходит увеличение мембранного потенциала нервного окончания, но нет изменений в пороге возбуждения собственно механочувствитель- ного субстрата рецептора (см. стр. 274). Каков конкретный механизм влияния адреналина и норадреналина на возбудимые мембраны, пока остается неясным. Возможно, что часть эффектов (гиперполяризация мембран) связана с работой натриевого насоса, который может активироваться под влиянием катехоламинов (Bulbring, Kuriyama, 1963; Somlyo, Somlyo, 1969; Kuba, 1970).
Приведенные данные дают основание заключить, что изменения в деятельности телец Пачини под влиянием умеренных доз адреналина и норадреналина протекают следующим образом. Вначале происходит повышение критического уровня деполяризации, т. е. возбудимость регенеративной системы рецептора несколько снижается. За этим кратковременным периодом следует более длительный отрезок времени, в течение которого благодаря возрастанию амплитуды и скорости нарастания РП чувствительность рецептора к действию механического стимула увеличивается (изменения РП превалируют над изменениями критического уровня деполяризации). Наконец, возбудимость рецептора вновь снижается вплоть до полного блока генерации ПД (амплитуда и скорость нарастания РП уменьшаются, и эти изменения суммируются с прогрессивным повышением критического уровня деполяризации, при котором происходит возникновение ПД). Если учесть еще возможность действия адреналина на сосуды, то легко понять ту фазность в действии этого вещества (равно как и возбуждения симпатических нервов) на деятельность тканевых механорецепторов in vivo, которая^отмечалась, как это указывалось выше, многими авторами.
Эффект адреналина следует сопоставить с влиянием на деятельность телец Пачини раздражения тонкого добавочного (эфферентного, по мнению ряда авторов) С-волокна, иннервирующего в ряде случаев эти рецепторы. В кратких сообщениях (Goto, Loewenstein, 1961; Loewenstein et al., 1962) отмечается, что данное волокно (оно встречалось лишь у 13% мезентериальных телец) в отличие от основного не возбуждается механическими толчками, а также не обладает чувствительностью к действию термических и химических стимулов. Оно проводит возбуждение со скоростью примерно 0.7 м/сек. (основное волокно в этих опытах обладало скоростью проведения 25—35 м/сек. при 32°). При изолированном раздражении С-волокна наблюдалось увеличение (на 10%) амплитуды РП, возникавшего в рецепторе при механической стимуляции. Чувствительность рецептора, определяемая по генерации ПД, при этом повышалась примерно на ту же величину. Эффект длился всего несколько минут, явлений депресии отмечено не было. Не наблюдалось и возникновения ПД при изолированном раздражении тонкого волокна.
Происхождение и функция тонкого дополнительного волокна, встречающегося у телец Пачини (и других инкапсулированных рецепторов) широко обсуждаются в литературе (стр. 21). Однако учитывая противоречивый характер имеющихся к настоящему времени данных (Черепнов, 1968а; Santini, 1968, 1969; Santini et al., 1971; Spencer, Schaumburg, 1973; Челышев, 1975), следует заключить, что в целом этот вопрос требует еще дополнительного изучения.
В экспериментах на кожных механорецепторах лягушки при регистрации импульсной активности было показано, что раздражение симпатических нервов, а также подведение адреналина или норадреналина вызывает четкое изменение деятельности рецепторов. По данным Левенстейна (Loewenstein, 1956а), раздражение симпатической цепочки (нанесение адреналина или норадреналина) сказывалось лишь на ответах фазного типа и практически не изменяло реакции медленно адаптирующихся рецепторов. Количество импульсов в ответе фазного рецептора на тактильный стимул резко возрастало (достигая увеличения в 15 раз). Так как при этом порог ответа снижался незначительно (редко более чем на 10%), следует заключить, что в основном уменьшалась скорость адаптацииУрецепторов. Раздражение симпатических нервов могло вызвать даже появление импульсов от кожных механорецепторов.
Увеличение числа импульсов в ответе на механическое раздражение, которое проводится на фоне активации симпатических нервов, отмечал также Чернетски (Chernetski, 1964). Наиболее выраженное влияние симпатических нервов (и адреналина) наблюдалось при пороговых значениях адекватного раздражения. Другие авторы (Дмитриева, Есаков, 1969) в условиях нерастянутого кожного лоскута лягушки (при отсутствии спонтанной активности) отмечали наибольшее влияние раздражения симпатической цепочки на медленно адаптирующийся ответ, который значительно возрастал по длительности и числу импульсов. Симпатические влияния очень четко выступали на фоне явлений угнетения тактильных рецепторов, вызванных ритмическим раздражением (Дмитриева, Есаков, 1969). Такого рода влияния вообще характерны для действия симпатической нервной системы на возбудимые образования (Орбели, 1948).
Были представлены морфологические данные, показывающие наличие адренергической иннервации в слоях кожи, где залегают свободные нервные окончания, причастные к механорецепции (Fuxe, Nilsson, 1965). Однако трудно сказать, обусловлены ли результаты физиологов прямым действием симпатических волокон на рецепторы или же наблюдавшиеся эффекты являются следствием сокращения контрактильных структур кожи. Во всяком случае, чем больше была исходно натянута кожа, тем более выраженным был симпатический эффект. По мнению авторов (Douglas, Gray, 1953), отмечавших возбуждающее действие адреналина на кожные рецепторы млекопитающих, оно было обусловлено сокращением мышечных элементов окружающих тканей. Аналогично возбуждались, по-видимому, и некоторые механорецепторы внутренних органов (Pearce, Henry, 1955; Kolatat et al., 1957), а также барорецепторы каротидного синуса (Landgren et al., 1952; Witzleb, 1953; Diamond, 1955).
Изменение механических свойств механорецепторных приборов кожи под влиянием симпатической нервной системы наблюдалось Антоновой (1967). Отмечалась также возможность подобного влияния симпатической нервной системы на вибриссы (Fuxe, Nilsson, 1965). Гистологические исследования несомненно свидетельствуют о тесной связи симпатических нервных волокон и тканевых механорецепторов, хотя данные, говорящие о возможности существования синаптических контактов между ними, кажутся недостаточными и требуют дальнейшей проверки (Тимофеев, 1896; Fuxe, Nilsson, 1965; Chouchkov, 1968; Andres, 1969; Santini et al., 1971). Все изложенные материалы заставляют склониться к точке зрения, что симпатическая нервная система в нормальных условиях скорее всего лишь изменяет ответы механорецепторов (прежде всего облегчает их возникновение) при действии адекватных для них стимулов.
Наряду с облегчающим действием симпатической нервной системы на механорецепторы были описаны и эффекты угнетения. Так, при раздражении симпатического ствола на медленно адаптирующихся рецепторах вибрисс (Nilsson, 1972, 1974) показано, что динамический ответ при этом несколько угнетался, а статический практически не изменялся. При тактильном раздражении кожи Есаков и Дмитриева (1968) отмечали торможение деятельности рецепторов, расположенных на симметричном участке кожной поверхности. В первую очередь изменялась частота импуль- сации и в меньшей степени — длительность ответа. Часть этих эффектов угнетения авторы относят за счет соматической нервной системы (ее влияния реализуются с меньшим латентным периодом и являются более краткими), а часть — за счет симпатических рефлекторных влияний.
Существенной особенностью симпатического эффекта во всех случаях являлся крайне большой латентный период (от нескольких секунд до 1 минуты, по данным разных авторов) и значительная длительность (от нескольких секунд до нескольких минут). В специальных экспериментах удалось показать, что при раздражении симпатических нервов выделяется адренолиноподобное вещество, которое может быть перенесено на другой участок кожи и оказывает на его тактильные рецепторы симпатический эффект (Loewenstein, 1956а; Дмитриева, Есаков, 1969). Следует подчеркнуть, что эффект раздражения нервов можно было воспроизвести нанесением на препарат небольших доз адреналина или норадреналина.
Гуморальные и другие периферические взаимодействия. К процессам эфферентной нервной регуляции деятельности механорецепторов тесно примыкают явления периферического взаимодействия. Речь идет о влиянии возбужденных рецепторных приборов на функциональное состояние соседних невозбужденных рецепторов. Эти влияния носят тормозный характер и могут рассматриваться как явления латерального торможения, аналогичные тем, которые были впервые обнаружены в органе зрения мечехвоста Limulus (Hartline, 1949; Hartline et al., 1952; Hartline, Ratliff, 1957).
Такого рода влияния в коже могут осуществляться двумя путями: нервным — с помощью аксон-рефлекса, и гуморальным — при участии биологически активных веществ, выделяющихся из активируемых структур ткани в ходе действия механического раздражения (Есаков, Дмитриева, 1971). Понятно, что во втором случае тормозные влияния могут не ограничиваться только тем рецептивным полем, которое подвергается раздражению, но могут и распространяться на соседние рецептивные поля.
Угнетение деятельности механорецепторов под влиянием антидромно посланных импульсов неоднократно отмечалось в литературе (Cattell, Hoagland, Д931; Eyzaguirre, Kuffler, 1955b; Diamond et al., 1958a; Lindblom, 1958; Loewenstein, 1958; Loewenstein, Altamirano-Orrego, 1958; Lindblom, Tapper, 1967; Есаков, Дмитриева, 1971). При этом затруднялось возникновение нормальных ПД (благодаря наличию абсолютного и относительного рефрактерных периодов в регенеративной системе генерации после «вхождения» в рецептор антидромного импульса), амплитуда же РП при этом мало изменялась (Loewenstein, Cohen, 1959b; Sato, Ozeki, 1963). По-видимому, по аналогичному механизму изменяется деятельность рецепторов одного и того же рецептивного поля, т. е. окончаний одного и того же разветвляющегося нервного волокна, когда один или несколько составляющих его элементов приходят в состояние возбуждения и начинают генерировать импульсную активность (Adrian et al., 1931; Katz, 1950a; Есаков, Дмитриева, 1971).
Характер тормозного влияния возбужденных рецепторов на неактивные рецепторы определяется прежде всего параметрами раздражающего воздействия. При легком ритмическом, но длительном (до 1 мин.) тактильном раздражении кожи соседние чувствительные точки оказывались заторможенными на 4—5 мин. (Есаков, Дмитриева, 1967). При этом максимум торможения возникал через 1—2 мин. после окончания раздражения. Ответ рецептора под влиянием такого торможения мог превратиться из тонического в фазный.
Вопрос о высвобождении из кожных структур веществ, изменяющих возбудимость рецепторных элементов, также существует в литературе. Известное повышение чувствительности нервных окончаний в коже после ее легкого повреждения (поскабливания) давно трактовалось (Echlin, Ргоррег, 1937) как следствие выделения каких-то веществ или ионов (например, калия) из клеточных элементов ткани.
С помощью более адекватной методики антидромного раздражения кожных нервов (Habgood, 1950) действительно показано выделение из кожи гуморальных факторов, которые могли распространяться от одного участка ткани к другому и повышать чувствительность его рецепторных структур. Таким веществом, по мнению Хебгуда, может служить гистамин.
Возможность гуморального влияния, оказываемого возбужденными нервными элементами кожи на покоящиеся рецепторы, была продемонстрирована также и другими авторами (Дмитриева, 1967; Есаков, Дмитриева, 1967) в опытах с антидромной стимуляцией афферентных нервов. В экспериментах на десимпатизированном ухе кролика (Holton, Holton, 1954; Holton, 1959) обнаружено, что при антидромном раздражении чувствительного нерва могут выделяться различные биологически активные вещества, такие, например, как аденозинтрифосфат (АТФ). Правда, действие именно АТФ, по-видимому, иное — тормозящее (Дмитриева и др., 1973). Оказалось, что при антидромном раздражении кожного нерва или адекватном механическом воздействии на кожу в наружном перфузате появляется активное начало, оказывающее тормозное влияние на чувствительность и развитие адаптационных явлений в кожных механорецепторах. В перфузате обнаруживаются различные вещества (нуклеотиды, ионы кальция и т. д.), но, по мнению авторов, основным активным началом является АТФ. Подведение АТФ к коже приводило к отчетливому торможению ответов механорецепторов на адекватное раздражение.
Полученные факты представляют несомненный интерес и при анализе механизмов нервно-тканевых взаимоотношений (см. главу 9). Несомненно, что именно химические факторы являются определяющими в такого рода взаимодействии. Природа же этих факторов сейчас еще совершенно неясна.
Регуляция активности проприоцепторов
Эфферентная регуляция мышечных веретен изучена наиболее детально. Первоначальные сведения о существовании нервных волокон, проводящих эфферентные влияния от центральной нервной системы к мышечным веретенам, были получены морфологами (стр. 41), которые показали, что пластинчатые окон чания в полярных зонах интрафузальных волокон у млекопитающих являются окончаниями двигательных нервов. В дальнейшем были получены и физиологические доказательства существования эфферентной регуляции веретен. Изучение афферентных разрядов веретен (Matthews, 1933) показало, что при раздражении двигательного нерва, превышающем на 10—20% максимальное, удается получить увеличение афферентной импульсации. Этот эффект, как стало ясно позднее, был обусловлен сокращением интрафузальных мышечных волокон, наступающим в результате возбуждения тонких нервных волокон при супрамаксимальных раздражениях моторного нерва.
Эфферентные гамма-волокна. Первые прямые электрофизиологические исследования эфферентной импульсации, приходящей к мышечным веретенам, были проведены в Швеции в лаборатории Гранита (Leksell, 1945). На основании этих экспериментов можно было заключить, что тонкие нервные волокна двигательного нерва (гамма-волокна) являются моторными для интрафузальных мышечных волокон. Эти нервные волокна обладали очень высоким порогом возбуждения (в 3.5—4.2 раза превышающим порог для двигательных альфа-волокон экстрафузальной мускулатуры). Полное вовлечение их в процесс возбуждения наступало лишь при силах воздействия, превышающих в 12—23 раза порог (все альфа- волокна нерва возбуждались уже при интенсивностях в 2.3— 3.5 раза выше порогового). Помимо своей низкой чувствительности к электрическому току, гамма-волокна оказались менее чувствительными и к сдавливанию, что позволяло избирательно блокировать активность альфа-волокон и изучать деятельность только гамма-системы. Дальнейшие исследования эфферентной системы мышечных веретен, начиная с работы Каффлера и сотр. (Kuf- fler et al., 1951), проводились преимущественно в условиях изоляции одиночных нервных волокон.
В настоящее время можно считать установленным, что гамма- волокна у млекопитающих являются основным эфферентным путем к мышечным веретенам. Их раздражение приводит к сокращению интрафузальных волокон и растяжению центральной зоны веретена. В результате нервные окончания, заложенные в этой области, деформируются, что и приводит к возникновению возбуждения в афферентных волокнах. Перемещение витков нервных окончаний, намотанных вокруг ядерной сумки мышечного веретена кошки, при активации эфферентных двигательных волокон удалось наблюдать непосредственно под микроскопом (Boyd, 1966). Перемещение и деформация нервных окончаний веретена в результате сокращения интрафузальных волокон были такими же, как при растяжении всей мышцы, в которой расположено веретено. Как показали уже первые работы по изучению эфферентной (фу- зимоторной) иннервации веретен, раздражение гамма-волокон может существенно изменять ответы рецепторов, вызванные адекватным раздражением.
Гамма-волокна составляют 30% общего количества волокон передних корешков. Они часто обладают тонической активностью, которая даже у спинальных животных может достигать 20— 60 имп./сек. (Hunt, 1951). Деэфферентация интрафузальных волокон приводит к установлению чрезвычайно регулярного ритма афферентного разряда (Matthews, 1933). У животных же с интактными эфферентными путями такая регулярность в активности рецепторов отсутствовала. Нерегулярность появления афферентных импульсов от веретен резко возрастает после децеребрации, что было обусловлено существенным повышением активности гамма-системы у таких животных (Kobayashi et al., 1952; Eldred et al., 1953; Granit, Kaada, 1953).
Наряду с аритмичностью разрядов для интактных веретен был установлен более высокий уровень активности. Наибольшей активностью обладали веретена у децеребрированных животных, у которых практически все рецепторы даже в состоянии покоя выявляли спонтанную импульсацию. У децеребрированных кошек помимо увеличения фоновой активности и появления пачечного режима генерации возрастает также и величина ответа на раздражение. Кроме того, замедляется и скорость адаптации. Порог возбуждения активных веретен был, естественно, значительно ниже, чем неактивных. Все это лишний раз свидетельствует о важной функциональной роли эфферентных влияний в деятельности веретен.
Последующие детальные исследования показали, что имеются определенные различия в поведении первичных и вторичных окончаний. У интактных веретен первичные окончания обладали значительно менее регулярным разрядом, чем вторичные окончания (Pascoe, 1965; Stein, Matthews, 1965; Matthews, Stein, 1969b; Eysel, Griisser, 1970). Это положение хорошо иллюстрирует рис. 73, на котором представлены изменения мгновенной частоты разрядов первичного и вторичного окончаний при наличии фузи- моторной активности и после ее устранения. Ответы вторичных окончаний были менее вариабельны по сравнению с первичными окончаниями даже после деэфферентации веретен. Большая стабильность в деятельности вторичных окончаний говорит о том, что они могут давать в центральную нервную систему более точные сведения о длине мышцы.
В целом влияние гамма-системы на веретена оказалось весьма сложным, что определялось рядом обстоятельств. Реакция веретена возрастала по мере увеличения частоты эфферентной импуль- сации, увеличения частоты раздражения, а также при возрастании фонового растяжения мышцы (Leksell, 1945; Kuffler et al., 1951). Как отмечалось выше (стр. 159), при сокращении мышцы в активности веретена наблюдается пауза. Под влиянием раздражения гамма-волокон происходит уменьшение паузы (рис. 74). Заполнение паузы импульсами веретена, обусловленными активностью гамма-волокон, было тем значительнее, чем больше было исходное напряжение мышцы и соответственно веретена. Интересной особенностью ответов некоторых веретен было существование явления послевозбудительного облегчения: активность веретена сохранялась и после прекращения раздражения гамма-волокон (Hunt, Kuffler, 1951а; Kuffler et al., 1951), что, по-видимому, зависит

Рис. 73. Вариабельность разрядов первичного (Пера.) и вторичного (Втор.) окончаний при наличии (Б) и в отсутствие (Л) фузимоторной активности (по: Matthews, Stein, 1969b).
Мгновенные значения частоты разряда (имп./сек.) двух окончаний (1, 2) в камбаловидной мышце децеребрированной кошки до (Б) и после перерезки переднего корешка (А). Слева — шкалы частоты импульсов в ответах. Отметка времени — 1 сек.
от тонических свойств интрафузальных мышечных волокон, изучавшихся в этих экспериментах, т. е. от особенностей организации вспомогательных структур рецепторов.
Исследования, проведенные Мэтьюзом с сотрудниками (Matthews, 1962; Crowe, Matthews, 1964а, 1964b; Brown et al., 1965; Brown, Matthews, 1966), а затем и другими авторами (Bessou et al., 1965; Appelberg et al., 1966; Bessou, Laporte, 1966; Bessou et al., 1966; Emonet-Denand et al., 1966; Jansen, 1966), позволили выявить ряд важных особенностей в деятельности гамма-системы. Оказалось, что все гамма-волокна в зависимости от их действия на мышечные веретена можно подразделить на две группы. Волокна первой группы обладали способностью возбуждать первичные окончания веретен в период динамического действия раздражения (растяжения мышцы). Они получили название динамических волокон. Проводники же второй группы действовали возбуждающим образом в период статической фазы растяжения и получили
наименование статических волокон. Следует отметить, что при постоянной длине мышцы раздражение волокон обоих типов приводит к возбуждению первичного окончания. Хотя в этом случае возбуждающее действие статического волокна было более выражено, тем не менее основные различия между разными типами
А Б в
/Ж11ПШ
1ЦШ11Ш11Н1Ш11шшш|щщ||
^IIIB ]£ЦИИ
Рис. 74. Влияние сокращения мышцы на разряд веретена в ответ на раздражение фузимоторного волокна (по: Hunt, Kuffler, 1951).
I — фоновый разряд; II — раздражение гамма-волокна с частотой 100/сек. (9 стимулов наносятся в начале каждой записи); III — одновременное раздражение гамма-волокна и альфа-волокон, вызывающих сокращение мышцы; IV — изолированное раздражение альфа-волокон. Сверху вниз', импульсная активность веретена длинного сгибателя; изменение напряжения мышцы (исходное натяжение для А — 2, для Б — 15, для В — 35 г). Максимальное напряжение при тетанусе — 140 г. Отметка времени — 200 мсек.
гамма-волокон выделяются наиболее четко при динамических воздействиях (рис. 75 и 76).
Динамические волокна вызывают увеличение способности веретена отвечать на действие динамической составляющей раздражения, статические ее уменьшают, несмотря на то, что в целом и статические волокна обладают значительным возбуждающим действием. Способность реагировать на динамический компонент воздействия оценивалась по динамическому индексу, т. е. по относительной величине изменения частоты разряда, наблюдаемого в первые 0.5 сек. после окончания динамической фазы линейного растяжения (Jansen, Matthews, 1962; Crowe, Matthews, 1964a).
Как видно из рис. 77, увеличение динамического индекса при раздражении динамического гамма-волокна наблюдалось во всем диапазоне используемых скоростей растяжения и возрастало при увеличении частот стимуляции. Напротив, статическое волокно вызывало падение динамического индекса. Частота раздражения волокна в этом случае мало сказывалась на эффекте. Оценивая динамический индекс, можно было легко отдифференцировать динамические волокна от статических (рис. 76). Волокон промежуточного типа обнаружено не было.
Действие динамических волокон находится в зависимости от скорости растяжения, что хорошо видно из рис. 77. Влияние же статических волокон мало изменяется при варьировании скорости изменения раздражающего воздействия на рецепторы.
Известно (см. стр. 44), что одно гамма-волокно может иннер вировать несколько различных веретен. Как показали специальные исследования, влияния волокон одного типа на деятельность всех иннервируемых ими рецепторов было одинаковым, т. е. динамические волокна оказывали во всех случаях динамическое действие, а статические — статическое (Crowe, Matthews, 1964b; Brown et al., 1965; Bessou et al., 1966). Так как в любом мышечном веретене имеется лишь одно первичное окончание, полученные результаты позволяют говорить о существовании истинной специфичности динамических или статических фузимоторных волокон, а не о наличии каких-то случайных соотношений, имеющих место в том или ином рецепторе.
При снятии статической нагрузки с рецептора на фоне активности фузимоторных волокон различия в их «динамическом» действии выявляются весьма четко. В период деятельности динамического волокна прекращение растяжения приводит к резкому уменьшению частоты импульсации, за которым следует постепенный возврат к фоновому уровню. При раздражении статического волокна афферентная импульсация после снятия нагрузки с мышц снижается постепенно по мере возвращения мышцы в исходное состояние (рис. 78).
Как показало изучение активности вторичных окончаний (Crowe, Matthews, 1964b; Appelberg et al., 1966; Brown et al., 1967), статические фузимоторные волокна оказывают на них влияние, сходное с их действием на первичные окончания. Влияние же динамических волокон, по-видимому, ограничивается только первичными окончаниями. Следует учесть, что вторичные окончания у кошек, на которых проводились исследования, как правило, расположены на ЯЦ-волокнах. Действуют ли статические волокна на вторичные окончания, относящиеся к ЯС-волокнам, остается неясным.
6 IIIII lllllllllll 11111111
6 11111II11 111 III II llllllllllllllllllll lllllllllllllllllllllll
Рис. 75. Эффекты раздельной стимуляции статических и динамических фузи- моториых гамма-волокон у кошки (по: Crowe, Matthews, 1964а).
а — растяжение мышцы на 6 мм со скоростью 30 мм/сек.; б — разряд веретена в отсутствие раздражения гамма-волокна; в, г — изменения разряда веретена при раздражении одного эфферентного статического (в) и одного динамического (г) волокна с частотой 70 имп./сек. на протяжении всей записи. Отметка времени — 0.1 сек.
имп./сек
200 г
юо
50
имп./сек.![]()


![]()

-50
100
Рис. 76. Влияние одиночных динамических (белые кружки) и статических (черные кружки) фузимоторных волокон на динамический индекс и статический разряд первичных окончаний в передней мышце голени у кошки (по: Brown et al., 1965).
По оси абсцисс — величина статического компонента импульсного ответа (имп./сек.); по оси ординат — динамический индекс (имп./сек.). Каждая точка — действие отдельного моторного волокна на отдельное первичное окончание. Мышца растягивалась на 3 мм со скоростью 5 мм/сек.; частота раздражения фузимоторных волокон — 100/сек.; увеличение статической активности измерялось при начальной длине.
Было рассмотрено также влияние динамических фузимоторных волокон на различные компоненты ответной реакции первичных окончаний (Lennerstrand, Thoden, 1968b, 1968с). Было установлено, что при активации динамических волокон чувствительность рецепторов к смещению (положению) остается либо без изменений, либо понижается. Чувствительность первичных окончаний оценивалась по наклону кривой, связывающей частоту разряда с величиной стационарной длины мышцы. Напротив, динамические компоненты реакции изменялись существенно больше. При этом медленный компонент реакции на скорость изменялся в большей саепени, чем быстрая составляющая. Статические волокна несколько усиливают медленный компонент реакции. В действии этих волокон на чувствительность к положению удалось выявить два типа реакций. При эффекте одного типа чувствительность к положению либо не меняется, либо даже снижается, а при другом типе влияний чувствительность возрастает и тем больше, чем выше частота стимулирования фузимоторных волокон.
При одновременном раздражении двух фузимоторных волокон характер ответной реакции зависел от исследуемой комбинации волокон (Crowe, Matthews, 1964b; Lennerstrand, 1968; Brown et al., 1969). При раздражении статического и динамического волокон ответ веретена был несколько больше, чем при возбуждении каждого волокна в отдельности, но меньше суммы величин этих двух ответов. Помимо частичной суммации при такой комбинации чаще встречалась окклюзия меньшего ответа большим как применительно к стационарному компоненту реакции, так и к чувствительности первичного окончания к смещению.
При раздражении двух динамических волокон наблюдалась частичная суммация стационарного ответа, а чувствительность к положению имела промежуточную величину. При комбинации из двух статических волокон (а в отличие от предыдущих случаев ее можно было получить не только для первичных, но и для вторичных окончаний) имела место частичная суммация эффектов в отношении стационарного разряда и чувствительность к скорости и положению изменялась примерно одинаково, характер же изменений варьировал. При раздражении нескольких статических волокон, что больше приближалось к естественной ситуации, наблюдалось повышение чувствительности к положению.
При раздражении первичного нервного окончания с помощью синусоидальных раздражений (исследование вибрационной чувствительности) можно было зарегистрировать афферентный разряд, приуроченный в случае низкочастотных воздействий к моменту восходящей фазы стимула. Способность веретена отвечать на вибрационные стимулы разной частоты, а также характер самого ответа сильно изменяются под влиянием фузимоторной активности. Фузимоторные волокна увеличивают частоту раздражения, воспроизводимую первичными окончаниями (примерно, в два раза — с 200 до 400 гц) (Granit, Henatsch, 1956). Вибрационный стимул

Рис. 77. Влияние динамического (Л) и статического (Б) фузимоторных волокон на динамический индекс при различных скоростях растяжения камбаловидной мышцы у кошки (по: Crowe, Matthews, 1964а).
По оси абсцисс — скорость растяжения (мм/сек.); по оси ординат — динамический индекс (имп./сек.). Частота раздражения волокон указана на кривых (б/с — без стимуляции). Амплитуда растяжения — 6 мм.

Рис. 78. Мгновенная частота импульсного ответа первичных окончаний на снятие растяжения в период фузимоторной активности (по: Crowe, Matthews, 1964b).
Реакция двух различных рецепторов (А, Б) в камбаловидной мышце у кошки на прекращение растяжения. Горизонтальная линия — раздражение динамического (А) и статического (Б) гамма-волокон с частотой 70/сек. Уменьшение растяжения со скоростью 5 мм/сек. при общем укорочении 6 мм. Отметка времени — 1 сек. Слева —» шкала частоты импульсов в разряде (имп./сек.). жении динамического волокна она становилась высокой (Crowe, Matthews, 1964b). При более высоких частотах воздействия (свыше 1 гц) чувствительность первичного окончания возрастала во всех случаях. При этом при частотах воздействия выше 5 гц увеличение чувствительности во время раздражения статического волокна происходило быстрее, чем при действии динамического волокна. Наконец, при частоте вибрации порядка 100 гц чувствительность рецептора во время раздражения обоих типов гамма-волокон становилась одинаковой. Это было обусловлено скорее свойствами самих рецепторов, чем особенностями фузимоторных влияний (Matthews, 1972).
dll ■! ...Ml 111 -111 !■ H ■
,
IIIIIIHl
llllllin ■ 1НМИ1М1ММИИ
Рис. 79. Влияние фузимоторной стимуляции на ответ первичного окончания при действии синусоидального растяжения средней силы (по: Crowe, Matthews, 1964b).
1 — действие динамического и 2 — статического гамма-волокон. Сверху вниз: биоэлектрическая активность; растяжение мышцы (амплитуда растяжения — 1 мм, частота — 3/сек.); стимуляция волокон (частота 100/сек.).
При раздражении фузимоторных волокон с разной частотой можно было наблюдать, как афферентный разряд веретена воспроизводит ритм раздражения. Способность первичных окончаний веретена следовать за ритмом фузимоторной стимуляции (driving) исследовалась в ряде работ (Kuffler et al., 1951; Besson, Laporte, 1962; Crowe, Matthews, 1964b; Brown et al., 1965; Appel- berg et al., 1966; Emonet-Denand et al., 1966; Besson et al., 1968; Emonet-Denand, Laporte, 1969). Было установлено, что «драйвинг» первичных окончаний имеет место только при раздражении статических волокон. Динамические волокна не обладают способностью вызывать ответ первичных окончаний в ритме своего раздражения. Стимулирование статических волокон, оканчивающихся на вторичных окончаниях, как правило, приводило к появлению афферентного ответа в ритме раздражения. Так как большая часть статических гамма-волокон оканчивается на ЯЦ-волокнах веретен, то можно заключить, что именно эти волокна обладают способностью к быстрым сокращениям. Однако, по-видимому, воспроизведение ритма фузимоторного раздражения не является исключи
тельной особенностью этого типа интрафузальных волокон. Считается более вероятным (Granit, 1970), что указанной особенностью обладают те сенсорные окончания, около которых вплотную располагаются двигательные окончания фузимоторных волокон. В этом случае даже крайне незначительные сокращения интрафу- зальной мускулатуры будут вызывать в области разветвлений моторных волокон возбуждение нервных окончаний и появление афферентного разряда.
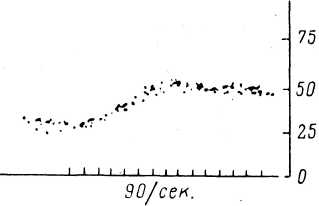
Для анализа свойств веретена использовался своеобразный метод графического усреднения афферентного отве1а (Besson et al., 1968). Смысл этого метода заключается в том, что на один кадр записывается ряд последовательных ответов рецептора на раздражение в виде мгновенных частот. Благодаря тому, что моменты появления спайков относительно раздражения флуктуируют, удается получить картину непрерывного изменения частоты, получившую название частотограмма. Такой способ регистрации ответной реакции позволил получить дополнительную информацию о свойствах веретен, и в частности о скорости интрафу- зального сокращения. В этом случае раздражение статического фузимоторного волокна стимулами умеренной частоты (22 и 47 в 1 сек.) вызывает ответы первичного окончания, которые сходны с миографической записью серии быстрых мышечных сокращений (рис. 80). При увеличении частоты раздражения наблюдается постепенное слияние отдельных ответов, и реакция принимает вид, сходный (тетаническим сокращением. Это позволяет предположить, что статическое гамма-волокно влияет на мышечные волокна, обладающие способностью к быстрым сокращениям. Напротив, при раздражении динамического фузимоторного волокна ответная реакция первичного окончания имеет вид, позволяющий предположить, что данный тип гамма-волокон влияет на медленно сокращающиеся интрафузальные волокна (рис. 79).
При одновременной регистрации с помощью частотограммы реакции первичного и вторичного окончаний одного и того же веретена на раздражение статического фузимоторного волокна было выявлено (Besson, Pages, 1969а), что частота стимулирования, при которой наблюдается слияние ответов, у вторичного окончания ниже. Этого следовало ожидать, так как вторичное окончание располагается в области интрафузального волокна, обладающей большей вязкостью.
При использовании частотограммы необходимо учитывать, что хотя она и позволяет делать определенные заключения о характере сокращений интрафузальных волокон, но отнюдь не является прямым отображением этих сокращений.
Изучение скоростей проведения в фузимоторных волокнах не выявило существования каких-либо резких отличий между волокнами, относящимися к разным типам (Brown et al., 1965; Besson et al., 1965; Emonet-Denand et al., 1966). И динамические,
75
-
. v-л* Щи к/-
I
I
-,200
-100
i -100
•
A
9
180/сек,
Рис.
80. Частотограммы, полученные для
первичного окончания m. tenis- simus при
раздражении статического (Л) и
динамического (Б) фузимоторных волокон
(по: Besson et al., 1968).
Каждая
частотограмма — результат наложения
30 записей мгновенных частот разряда
за один и тот же временной интервал. По
горизонтали —
импульсы стимуляции фузимо- торного
волокна с указанной частотой (цифры
под каждым кадром). Шкалы
(справа от каждого кадра) — мгновенные
значения частот (имп./сек.).
![]()
![]()

t
i.l 1 I I I I I I I J t-L
■ J Q
90/сек.11111111
U11IIH1.1LI
I 1 I 11U-LX1
J 0
и статические волокна обладали скоростями проведения порядка 25—50 м/сек. В отдельных случаях статические волокна обладали более медленной скоростью проведения. Так, например, в задней большеберцовой мышце кошки все фузимоторные волокна со скоростями проведения меньше 25 м/сек. относились к статическим. Общее количество статических волокон превышает число динамических: у кошки соотношение волокон составляет для разных мышц 3 : 1 или 2:1, а у кролика — 6:1 (Crowe, Matthews, 1964а; Emonet-Denand et al., 1966). Это позволяет говорить о большей функциональной значимости именно статических волокон.
При анализе механизмов действия фузимоторных волокон важно знать, какого рода потенциалы возникают под их влиянием в интрафузальной мускулатуре. Первые работы, выполненные в этом направлении (Kuffler et al., 1951; Eyzaguirre, 1960a, 1960b), были осуществлены до открытия статических и динамических гамма-волокон и дали весьма противоречивые результаты. В дальнейшем при использовании раздельного раздражения разных типов фузимоторных волокон было показано, что динамические волокна вызывают появление нераспространяющихся потенциалов, а статические вызывают потенциалы, возникающие по закону «все или ничего», обладающие овершутом и частично распространяющиеся от места своего возникновения (Bessou, Laporte, 1965; Bessou, Pages, 1969a, 1969b).
Сейчас еще нет исчерпывающей ясности в вопросе о причинах различного действия динамических и статических фузимоторных гамма-волокон (см. обзоры: Granit, 1970; Matthews, 1972). Вместе с тем ряд особенностей их влияния уже может получить вполне удовлетворительное объяснение. Очевидно, что различия в действии разных типов фузимоторных волокон следует искать в особенностях организации и распространения их концевых аппаратов, свойствах разных интрафузальных волокон, особенностях строения и функции элементов первичных и вторичных окончаний. Резюмируя данные литературы, Гранит (Granit, 1970) пишет, что ведущую роль играют две комбинации: 1) динамическое гамма- волокно—ЯС-волокно и 2) статическое гамма-волркно—ЯЦ-во- локно. Последняя комбинация объясняет, почему только статические волокна оказывают влияние как на первичные, так и на вторичные окончания. Остается неисследованным ряд возможностей, связанных с взаимодействием между кустиковидными и пластинчатыми окончаниями. Резкие различия между первичными и вторичными окончаниями в отношении их чувствительности к скорости и к положению, по-видимому, лучше всего объясняются различиями в свойствах интрафузальных волокон. ЯЦ-волокна обладают фазическими свойствами и почти лишены демпфирующей вязкости, а ЯС-волокна обладают вязкими, демпфирующими свойствами и вообще более сходны с медленными волокнами тонических пучков, способными сокращаться может рассматриваться как специфический стимул для изучения динамической чувствительности веретен. С этой точки зрения, следовало бы считать, что лишь раздражение динамических волокон должно оказывать влияние на способность рецепторов реагировать на действие вибрации. Можно было ожидать, что вторичные окончания, которые мало чувствительны к вибрационным стимулам (Bessou, Laporte, 1962; Bianconi, Meulen, 1963), не приобретут большую чувствительность под влиянием активности гамма-волокон. Последнее предположение было подтверждено (Brown et al., 1968). В отношении же воздействия разных типов фузимоторных волокон на первичное окончание дело оказалось сложнее. Во всех случаях максимальная частота, воспроизводимая первичными окончаниями под влиянием фузимоторных волокон, возрастала (Crowe, Matthews, 1964b; Brown et al., 1967), и это возрастание было примерно одинаковым при раздражении как динамических, так и статических волокон. Существует мнение (Bianconi, Meulen, 1963), которое разделяют и другие (Granit, 1970), что такое действие могли оказывать те фузимоторные волокна, которые оканчиваются в сравнительно недемпфирующем (невязком) участке веретена, вероятнее всего в его экваториальной части, где содержатся эластические элементы. Следует учесть, что у кошек, на которых проводились исследования, статические волокна, как правило, оканчиваются именно на ЯЦ-волокнах, содержащих в центральной области эластические структуры.
При использовании низкочастотных синусоидальных раздражений достаточно большой амплитуды ответы рецепторов были аналогичны тем, которые наблюдались при включении и выключении значительного линейного растяжения. В этом случае (рис. 79) динамическое фузимоторное волокно увеличивает ответ первичного окончания на фазе нарастания стимула и не оказывает влияния на фазе спадения (расслабления мышцы). Статическое же волокно вызывает увеличение средней частоты афферентной импульсации, и ответы на обе фазы раздражения становятся примерно одинаковыми. Пачечный режим активности окончания может полностью нарушиться, и при этом устанавливается непрерывная, довольно ритмичная импульсация веретена.
При небольших амплитудах воздействия ответная реакция первичного окончания под влиянием фузимоторной активности меняется несколько иначе. Теперь раздражение динамического волокна приводит к установлению непрерывной импульсации рецептора, хотя частота разряда продолжает быть больше на фазе нарастания растяжения. При раздражении же статического волокна частота раздражения возрастает и остается неизменной вне зависимости от фаз раздражения.
При небольших частотах раздражения (0.5—5 гц) первичное окончание по-разному изменяет свою чувствительность при возбуждении разных фузимоторных волокон: в ходе раздражения статического волокна чувствительность была низкой, а при раздра-
лишь при тетаническом раздражении. Первичные окончания ведут себя так, как если бы они отражали свойства ЯС-волокон, а вторичные — как если бы они в основном находились на ЯЦ- волокнах. Промежуточные первичные окончания реагируют таким образом, как если бы они были особенно хорошо снабжены концевыми веточками, получающими информацию от ЯЦ-волокон, что придавало бы им способность отвечать также на смещение, вызываемое уменьшением длины. Подробное обоснование связи динамических гамма-волокон с ЯС-волокнами, а статических
Таблица 5
Характеристика влияний динамических и статических типов фузимоторных волокон на первичные и вторичные окончания мышечных веретен (по: Matthews, 1972)
Характеристики
Динамические
волокна
Статические
волокна

