

Глава 2
ДЕПОЛЯРИЗАЦИОННЫЕ РЕЦЕПТОРНЫЕ ПОТЕНЦИАЛЫ
Существование локальных электрических реакций рецепторных структур отмечалось исследователями давно. Так, например, было описано (Frohlich, 1914, 1921) появление длительной деполяризации чувствительных структур глаза головоногих моллюсков во время освещения. Такого рода факты, а также данные общей нейрофизиологии, полученные на разнообразных сенсорных приборах, говорящие о тесных связях локальных и распространяющихся электрических реакций в различных нервных элементах, позволили шведским физиологам сформулировать теорию генераторного потенциала (Bernhard et al., 1942; Bernhard, Granit, 1946; Granit, 1947, 1955). Согласно этой теории, во всех сенсорных структурах при действии на них адекватного стимула возникает локальный деполяризационный ответ — генераторный потенциал, который в свою очередь обусловливает появление в отходящем нервном волокне бегущих волн возбуждения, т. е. вызывает генерацию потенциалов действия. Указанная концепция оказалась очень плодотворной для развития физиологии рецепторов. Хотя отдельные положения данной теории в дальнейшем подверглись уточнению, тем не менее в целом она получила полное подтверждение и повсеместное признание.
В современной литературе для обозначения локального ответа рецептора употребляют два основных термина: генераторный потенциал и рецепторный потенциал (Granit, 1955; Черниговский, 1960; Davis, 1961; Ильинский, 1964а, 1966г; Fuortes, 1971). Согласно предложению Девиса (Davis, 1961), которое принято большинством авторов, термин генераторный потенциал следует употреблять только в тех случаях, когда локальный электрический ответ рецептора, точнее локальные токи, лежащие в его основе, своим непосредственным действием вызывают появление импульсной активности в отходящем нервном волокне. В случае же, когда локальная электрическая реакция рецептора не ведет непосредственно к появлению импульсной активности в нерве (во вторичных рецепторах) или даже угнетает ее (например, при гиперполяризационном ответе), термин генераторный потенциал применять нельзя, ибо это противоречит смыслу, исходно вложенному в данное понятие. Более правильным будет использование в этих случаях термина рецепторный потенциал (РП), который может быть по знаку как негативным {деполяризационный, возбуждающий рецепторный потенциал}, так и позитивным (гиперполяризацион- ный, тормозный рецепторный потенциал) (см.: Ильинский, 1964а, 1965, 1966г). Так как негативные и позитивные локальные реакции первичных рецепторов часто наблюдаются в одной непрерывной реакции и следуют одна задругой, некоторые авторы в этих случаях пользуются обозначениями деполяризационная и гиперполяризационная фазы рецепторного потенциала. Однако локальная деполяризация и гиперполяризация могут появляться изолированно друг от друга, т. е. существовать самостоятельно. Поэтому лучше оперировать понятиями деполяризационный (возбуждающий) РП п синонимом его в случае первичных рецепторов — генераторный потенциал (ГП), а также термином гиперполяризационный (тор- мозпый) РП. Наконец, когда знак локальной реакции рецептора несуществен для изложения, мы будем пользоваться обычным названием: рецепторный потенциал (РП).
Как следует из теории генераторного потенциала, локальный электрический ответ является чрезвычайно важным звеном в цепи явлений, возникающих в сенсорном приборе при действии на него адекватного раздражения. Многие особенности в реакции рецепторов на раздражение связаны именно со свойствами РП. Поэтому исследователи уделяют его изучению очень большое внимание.
Как показали многочисленные исследования, электрические процессы, протекающие в рецепторах, являются неспецифическими. Это проявляется в том, что качество воспринимаемых сенсорной структурой стимулов не сказывается на характеристиках локальных и распространяющихся электрических ответов. Следовательно, выводы, сделанные на основе изучения электрических явлений, имеющих место в тех или иных рецепторных образованиях (например, механо- или фоторецепторах), могут быть распространены и на другие рецепторы. Следует признать, что значительная часть сведений об общих свойствах РП, была получена в опытах именно на механорецепторах (см. обзоры: Granit, 1955; Kuf- I’ler, 1958, 1960; J. А. В. Gray, 1959а, 1959b; Loewenstein, 1959a, 1961a, 1965, 1966, 1971; Черниговский, 1960, 1967; Eyzaguirre, 1961; Inman, 1962; Ильинский, 1966д, 1966e, 1967, 1971, 1972a; Sato, Ozeki, 1966; Flock, 1971a, 1971b; Fuortes, 1971; Ottoson, Shepherd, 1971a).
Одной из основных проблем, существующих в области физиологии рецепторов, является проблема первичных трансформационных процессов. Следует отметить, что сейчас трудно сказать, имеет ли место во всех случаях подлинная в строгом смысле слова трансформация одного вида энергии в другой. Именно эти процессы, предшествующие развитию локальных электрических явлений, т. е являющиеся промежуточным звеном между адекватным! стимулом и РП, и определяют специфичность сенсорной структуры. Анализ этих процессов во многих рецепторах, в том числе* и механорецепторах, в настоящее время ведется еще очень медленно, что связано с огромными техническими трудностями. Поэтому сведения о трансформационных процессах приходится черпать из ряда косвенных показателей, в частности из особенностей РП (их характеристик, изменений их свойств под влиянием' различных физических и химических факторов и т. д.).
В некоторых чувствительных приборах, таких как хемо- и: фоторецепторы, существование процессов трансформации очевидно. Это первичные и весьма специфические химические реакции, которые протекают в рецепторах под влиянием адекватных внешних стимулов (соответственно химических или -световых).. Совершенно ясно, что эти химические реакции следует различать» от последующих явлений и, в частности, от перемещений ионов; через мембрану рецептора, которые ведут к возникновению РП„ т. е. от процессов его генерации.
Цепь явлений, связанных с перемещением ионов через поверхностную- мембрану сенсорной структуры, можно называть процессами генерации' (Ильинский, 1964а, 1966г, см. также: Loewenstein, 1971). Несомненно, что явления, развивающиеся и на иных специализированных хемочувствитель- ных поверхностях клеток, таких как постсинаптические мембраны, также- можно разделять на процессы трансформации и генерации. Однако для избежания терминологической путаницы при рассмотрении вторичных рецепторов, где имеет место синаптическая передача, мы не будем использовать, термины трансформации и генерации в отношении постсинаптических образований.
Процессы трансформации менее очевидны в рецепторах, воспринимающих иные, нежели световые или химические, стимулы,, в частности в механорецепторах, однако осуществление трансформации в механорецепторах сейчас не вызывает сомнения.. Говоря о механорецепторах, под трансформационными следует понимать процессы, возникающие в механочувствительном субстрате рецепторов при действии деформирующих механических сил и приводящие к изменению проницаемости поверхностной рецепторной мембраны для ионов. Вероятно, благодаря существованию трансформационных процессов в рецепторах свойства мембран сенсорных элементов существенно отличаются от|свойств; обычных мембран нервных клеток. В отношении механорецепторов это будет показано при дальнейшем изложении.
В этой главе будет рассмотрен деполяризационныиУрецептор- ный потенциал. Это основной вид локальных ответов рецептора. Его свойства изучены в настоящее время весьма детально. Иначе обстоит дело с гиперполяризационным рецепторным потенциалом. Он был обнаружен позднее, и его систематическое изучение только начинается. Исследования гиперполяризационного РП в механорецепторах тесно связаны с изучением явлений чувствительности данных сенсорных приборов к действию направления механического стимула, дирекционной чувствительности, й эти вопросы целесообразно излагать совместно. Изучение дирекционной чувствительности позволило понять некоторые важные механизмы действия механического раздражения на механорецепторы. Поэтому рассмотрение дирекционной чувствительности в книге будет непосредственно предшествовать главе о механизмах действия механического раздражителя. В соответствии с данными главы 1 свойства РП будут рассматриваться раздельно для первичных и вторичных механорецепторов.
Деполяризационные рецепторные потенциалы тканевых механорецепторов у позвоночных
При адекватном раздражении первичного механорецептора в отходящем нервном волокне (при отведении активности вблизи от рецептора) наряду с импульсной активностью (потенциалами действия — ПД) удалось зарегистрировать и градуальные деполяризационные ответы (рис. 21). Эти ответы отсутствовали, если регистрирующие электроды размещались на некотором расстоянии от рецепторов, и были максимальными при отведении потенциалов непосредственно от самого рецепторного образования (Katz, 1950b; Alvarez-Buylla, Ramirez de Arellano, 1953; J. A. B. Gray, Sato, 1953; Eyzaguirre, Kuffler, 1955a, 1955b). Гилло очевидно, что данный вид электрической активности является проявлением деятельности собственно рецептора, его рецепторным потенциалом (РП). Различные особенности РП первичных механорецепторов были тщательно изучены в экспериментах на тельцах Пачини, мышечных веретенах, рецепторах растяжения у ракообразных, волосковых механорецепторах у насекомых и ряде других образований.
Следует заметить, что хотя число работ, посвященных изучению свойств РП в механорецепторах, прогрессивно нарастает от года к году, тем не менее оно составляет лишь малую часть всех исследований, проводимых на этих рецепторных образованиях, и, конечно, является совершенно недостаточным с точки зрения важности самой проблемы. Это хорошо видно на примере моханочувствительных нервных окончаний кожи, внутренних органов и особенно волосковых рецепторов органов чувств, где при общем чрезвычайно большом числе работ анализу особенностей РП посвящены лишь отдельные исследования, что является отражением прежде всего тех методических трудностей, с которыми сталкиваются экспериментаторы при изучении деятельности одиночных механорецепторов.
Рецепторные потенциалы в тельцах Пачини
При раздражении телец Пачини механическим стимулом в нервном волокне вблизи рецептора может быть зарегистрировано два типа активности: локальный ответ (РП) и рас
пространяющаяся волна возбуждения (ПД). В случае телец Пачини, так же как и ряда других механорецепторов, ПД сильно затрудняет изучение РП, а иногда даже полностью его маскирует (рис. 21, В). Однако в рецепторах свойства структур, ответственных за генерацию ПД {регенеративных структур), и структур,

Рис. 21. Локальные и распространяющиеся ответы различных механорецепторов (Л — по: Eyzaguirre, Kuffler, 1955а; Б — по: Shepherd, Ottoson, 1965; В — по: Ильинский, 1969д).
А — ответ медленно адаптирующегося рецептора растяжения у ракообразных (внутриклеточное отведение). Стрелки — начало и конец растяжения. Отметка времени — 1 сек.; калибровка — 25 мв. Б — ответ мышечного веретена лягушки. Сверху вниз: биоэлектрическая активность; отметка раздражения (скорость растяжения — 130 мм/сек.). Отметка времени — 50 мсек. В — ответы телец Пачини на адекватное механическое раздражение небольшой длительности, а, б — отведение потенциалов в 2—3 мм от выхода нервного волокна из капсулы рецептора (а — амплитуда раздражения, пороговая для возникновения ПД, б — амплитуда раздражения увеличена, РП полностью замаскирован развитием ПД); в — отведение потенциалов в области 2—3-го перехватов Ранвье (видны как РП, так и ПД); г — отведение потенциалов вблизи тельца Пачини (ПД блокированы новокаином). Отметка времени: на а, б — 2, на в — 1.5, на г — 1 мсек. Калибровка: на а, б — 100, на в — 50, на г — 20 мкв.
ответственных за возникновение РП (нерегенеративных структур), оказались существенно различными.V Это позволило с помощью различных методов блокировать ПД, сохраняя при этом РП. Блокировать ПД можно, например, с помощью небольших доз новокаина (рис. 21, В, г), тетродотоксина, с помощью механического сдавления начальной части мякотного волокна или аноди- ческой поляризации рецептора, а также нанося на рецептор через
короткий интервал два последовательных коротких стимула (.1. Л. В. Gray, Sato, 1953; Diamond, Gray, Sato, 1956; Loewenstein, Altamirano-Orrego, 1958; Loewenstein, Rathkamp, 1958; Ильинский, 1963). В последнем случае второй стимул не вызывает появления спайка, так как он попадает в рефрактерный период после первого ПД. Однако при этом сохраняется РП (в этом состоит одно из существенных отличий РП от ПД). В подобного рода экспериментах весьма удобно сравнивать оба вида активности нервного окончания телец Пачини: ПД, вызванный на первое воздействие, и РП, вызванный на второе (рис. 22, 1-^3).


Рис. 22. Соотношение рецепторного потенциала (РП) и потенциалов действия (ПД) в тельцах Пачини (1—3 — по: Ильинский, 1963; 4 — по: Nishi, Sato, 1968).
Раздражение двумя последовательными толчками (записи с наложением). 1,2 — интактный рецептор (интервал между стимулами уменьшается); з — частично декапсулирован- пый рецептор (первый стимул отсутствовал; в этом случае второй стимул вызывал появление ПД); 4 — ответы декапсулированного тельца Пачини на стимулы возрастающей амплитуды. Сверху вниз: биоэлектрическая активность; отметка раздражения. Отметка времени: на 1—3 — 1, на 4 — 5 мсек.; калибровка: на 1—з — 120, на 4 — 100 мкв.
В ответ на механический стимул в тельце Пачини возникает РП, который распространяется вдоль нервного волокна с декрементом, из-за чего он на расстоянии далее чем 3—5 мм от рецептора не регистрируется. При раздражении тельца Пачини кратким толчком умеренной силы деполяризационный РП имеет довольно типичный вид (рис. 21, В, г). Он возникает после небольшого скрытого периода, плавно нарастает и затем плавно падает до нуля; фаза нарастания короче фазы спадения.
Изучение места возникновения РП в тельцах Пачини показало (Loewenstein, Rathkamp, 1958), что ни элементы наружной капсулы, ни элементы внутренней колбы не являются тем субстратом, который ответствен за появление РП. Удаление практически всей капсулы рецептора (на 99.99%), окружающей немиелинизи- рованное нервное окончание, не изменяло характера ответной реакции. При манипуляциях же с собственно немиелинизирован-
нои частью нервного окончания ответ резко изменялся или даже полностью блокировался. Так, передавливание окончания между зоной, куда наносилось раздражение, и точкой отведения полностью блокировало появление РП. Дегенерация нервного окончания в результате предварительной перерезки уже через 24— 36 час. приводила к исчезновению РП, хотя элементы капсулы рецептора при этом сохранялись без заметных изменений. Не наблюдалось появления активности и при механическом раздражении миелинизированного нервного волокна, отходящего от рецептора.
пки
НО
-
30-
20-
Рис. 23. Зависимость амплитуды рецепторного потенциала от силы раздражения (по: Ильинский, 1966а).
I | i | I По оси абсцисс — сила раздраже-
п 500 1ППП 1^пп 9ППО нт л ния <УСЛ- ед-); по оси ординат —
и JUU 7UUU 7jUU ZUUUусл.е,',. амплитуда РП (мкв).
ных фактов пришли и другие исследователи (J. А. В. Gray, Sato, 1953; J. А. В. Gray, 1959а, 1959b; Ильинский, 1963, 1966а; Sato, Ozeki, 1966; Ильинский и др., 1968; Nishi, Sato, 1968).
Основной отличительной особенностью РП является его гра- дуальность. При увеличении силы раздражения амплитуда РП постепенно нарастает (рис. 22, 4 и 23). Существует представление, согласно которому в собственно рецепторной части нервного окончания имеется много независимых элементарных структур, каждая из которых может работать по принципу «все или ничего» (Loewenstein, 1961а; Ильинский, Фикс, 1963). В свете этого представления градуальность РП отражает постепенное вовлечение в процесс возбуждения все новых элементов рецепторной мембраны.
Поскольку градуальное возрастание РП возможно лишь до определенного предела (рис. 23), то наличие этого предела говорит о состоянии возбуждения всех рецептирующих элементарных структур нервного окончания. При очень больших величинах раздражения амплитуда РП снижается, что связано уже с необратимым повреждением рецептора. Диапазон смещений от пороговыхVI до повреждающих для телец Пачини весьма велик: от сотых долей микрона до десятков микрон (J. А. В. Gray, Sato, 1953; Loewenstein, Rathkamp, 1958; J. A. В. Gray, 1959a; Loewenstein, 1965; Ильинский, 1966a). При усилении раздражения наряду с, повышением амплитуды увеличивается и скорость нарастания РП. Амплитуда и скорость нарастания РП при прочих равных условиях зависят и от скорости нанесения механического раздражения. Такая зависимость наблюдается при скорости нарастания стимула менее 1—3 мкм/сек. (J. А. В. Gray, Sato, 1953; J. А. В. Gray, 1959а; Ильинский, 1965, 1966а; Nishi, Sato, 1968). При этом между скоростью нарастания РП и скоростью нарастания раздражения имеется линейная зависимость. Скорость нарастания РП продолжает увеличиваться и тогда, когда амплитуда ответа достигает максимума. Это имеет место как в интактных, так и де- капсулированных рецепторах (Nishi, Sato, 1968).
Измерение абсолютной величины РП в тельцах Пачини произвести не удается из-за малого диаметра немиелинизированной части нервного окончания. Сопоставление РП и ПД нервного волокна показало, что величина деполяризационного РП может достигать 80—95% от амплитуды спайка или даже равняться ПД (рис. 22) (Ильинский, 1963; Nishi, Sato, 1968). Следует учесть, что величина спайка в области 1—2-го перехватов Ранвье обычно несколько меньше, чем на некотором расстоянии от рецептора. С другой стороны, истинная величина РП в точке возникновения больше, чем в месте отведения, так как РП распространяется с декрементом и приходит к регистрирующим электродам, расположенным в области первых перехватов Ранвье, несколько уменьшенным. Учитывая связь РП с мембранным потенциалом, исследователи склонны считать величину максимального деполяризационного РП примерно равной потенциалу покоя нервного окончания (Loewenstein, Altamirano-Orrego, 1958; J. А. В. Gray, 1959а; Ильинский, 1963).
При увеличении амплитуды и скорости нарастания раздражения наряду с изменением амплитуды и скорости нарастания РП изменяется его латентный период Д который постепенно укорачивается. Минимальный латентный1период появления РП (при отведении потенциала в месте выхода нервного волокна из капсулы рецептора) составляет при комнатной температуре 0.20 мсек. (J. А. В. Gray, Sato, 1953), а при температуре тела животного — примерно 0.15 мсек. (Ильинский, 1966е). Минимальное же время возникновения РП в точке раздражения равняется всего лишь 0.05—0.06 мсек. (Ильинский, 1966г), что существенно меньше минимального времени синаптической задержки (Eccles, 1957, 1964). Короткий латентный период возникновения РП является одним из доводов для подтверждения, что тельца Пачини являются первичными рецепторами, т. е. рецепторами, у которых восприятие механического раздражения осуществляется непосредственно нервным окончанием, а не окружающими его элементами внутренней колбы (J. А, В, Gray, 1959а; Ильинский, 1966г, 1966е),
При раздражении телец Пачини механическими стимулами с крутым передним фронтом возникает РП с весьма короткой фазой нарастания (1—2 мсек, при комнатной температуре и 0.6—1.0 мсек, при температуре тела животного) и более длительной фазой спадения. Разрушение РП происходит экспоненциально. Длительность РП у разных рецепторов варьирует и равняется 2—8 мсек. (J. А. В. Gray, Sato, 1953; J. А. В. Gray, 1959а; Loewenstein, 1958; Ильинский, 1962, 1963). Как длительность, так и фаза нарастания РП существенно зависят от скорости нарастания раздражающего стимула (рис. 24) и практически не зависят от длительности статического компонента воздействия (рис. 25). При раздражении рецептора как кратким, так и продолжительным прямоугольным стимулом временное течение РП было примерно одинаковым. Такая особенность телец Пачини обусловлена некоторыми свойствами капсулы рецептора, благодаря которым к нервному окончанию проходит только динамическая составляющая стимула. После удаления капсулы нервное окончание начинает генерировать деполяризационные РП, длящиеся в течение всего периода действия стационарной деформации (Mendelson, Loewenstein, 1964; Loewenstein, Mendelson, 1965; Ozeki, Sato, 1965; Ильинский, 1966a; Nishi, Sato, 1968). Подробнее этот вопрос, относящийся к проблеме адаптации, будет рассмотрен в главе 4.
Если тельца Пачини с неповрежденной капсулой раздражали длительными стимулами, то возникновение ответа наблюдалось не только в момент включения раздражения (рп-ответ), но и при его выключении (pff-omeem) (рис. 25, 2). Механизмы данного явления, также зависящие от свойств вспомогательной структуры рецептора (свойств капсулы), будут рассмотрены ниже (стр. 120).
При нанесении на интактный рецептор двух последовательных стимулов можно зарегистрировать два РП. Меняя силу толчков и интервалы между ними, удалось обнаружить суммацию обоих РП, а также депрессию второго РП. Это же можно продемонстрировать и в опытах с изменением продолжительности достаточно долго длящегося стимула при взаимодействии on- и off-ответов (J. А. В. Gray, Sato, 1953; Diamond et al., 1958a; Loewenstein, Altamirano-Orrego, 1958; Ильинский, 1966д).
Анализ этих явлений проводился на декапсулированных рецепторах. При раздражении интактного тельца Пачини воздействие более или менее равномерно распределяется на всю поверхность немиелинизированного нервного окончания. При декапсуляции рецептора можно с помощью соответствующих устройств, вибраторных систем (на базе пьезокристаллов или электромагнитов), имеющих на конце стержень с различной площадью кончика (вплоть до 1 мкм), локально раздражать отдельные участки нервного окончания. Такая возможность избирательного воздействия на отдельные области рецепторной структуры отсутствует при работе с другими механорецепторами. Поэтому значительная часть аналитических исследований на тельцах Пачини проводилась после удаления капсулы рецептора. Детали техники экспериментирования с одиночными тельцами Пачини приведены в ряде

Рис. 24. Изменения рецепторного потенциала в зависимости от скорости нанесения механического стимула (по: Ильинский, 1966а).
< >'г а до е скорость нанесения стимула уменьшается. Длительность ответа увеличивается. Негативный ответ наблюдается лишь в момент действия заднего фронта толчка. Передний фронт либо не вызывает никаких изменений потенциала (д, е), либо вызывает появление позитивных ответов (а—г). Сверху вниз: отметка раздражения; РП. Отметка времени — !» мсек.; калибровка — 30 мкв.

Рис. 25. Влияние длительности раздражения на рецепторные потенциалы телец Пачини (ПД блокирован новокаином) (по: Ильинский, 1966а).
_ один, е — другой рецептор. От а до д длительность раздражения возрастает. Сверху вниз — то же, что на рис. 24. Отметка времени: на а—д — 5, на е — 4 мсек.; калибровка — 40 мкв.
Q Физиология механорецепторов
источников (Gray, Malcolm, 1950, 1951a, 1951b; J. A. B. Gray, Sato, 1953; Loewenstein, Altamirano-Orrego, 1958; Loewenstein, Rathkamp, 1958; Ильинский, 1963, 19646, 1966a, 19666; Mendelson, 1964; Ильинский, Кудрин, 1967).
Проведенный анализ показал, что в основе суммации и депрессии лежат эффекты, связанные с пространственным взаимодействием возбужденных участков рецепторной мембраны, а не какие-либо особенности вспомогательных структур (Loewenstein, 1959b). Как видно из рис. 26, раздражение в различной последовательности двумя стимулами, прикладываемыми к нервному окончанию, может привести как к суммации обоих РП — при раздражении различных участков Й>рецепторной поверхности
* У - у 7 2 j \ 1
Л— riflag \ / дав 1 дпд
■ЧЕЗ® четг -лете
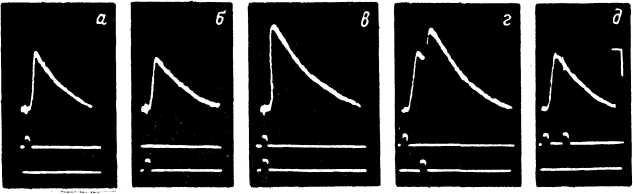
Рис. 26. Пространственная суммация рецепторных потенциалов в нервном окончании телец Пачини (по: Loewenstein, 1959а).
А — схема независимой стимуляции двух участков (7, 2) мембраны декапсулированного нервного окончания тельца Пачини; В — РП в области миелинизированного нервного волокна, а — изолированное раздражение участка 1; б — изолированное раздражение участка 2', в — одновременное раздражение участков 1 и 2\ г — последовательное раздражение участков 1 и 2 с интервалом в 1 мсек.; д — повторное раздражение участка 1 с интервалом в 1 мсек. Отметка времени — 1 мсек.; калибровка — 50 мкв.
(рис. 26, в—г), — так и к депрессии второго РП — при воздействии на одно и то же место рецептора (рис. 26, д). Эти данные говорят о том, что процессы, развертывающиеся на рецепторной мембране Л носят локальный характер.
В пользу такого представления говорят и другие факты, полученные на декапсулированных рецепторах. Если раздражать один из участков мембраны ритмическими стимулами большой частоты, можно через некоторое время получить значительное снижение величины РП. Соседний же участок мембраны, который ритмически не раздражался, при этом сохраняет все свои свойства неизмененными. Наконец, при раздражении одного конца нервного окончания тонкой микроиглой и отведении потенциалов с помощью микроэлектрода, помещаемого на различном расстоянии от точки приложения механического стимула, выяснилось, что амплитуда РП закономерно уменьшается по мере увеличения расстояния (Loewenstein, 1961а). Логарифм амплитуды деполяризационного
Pit и расстояние оказались связанными линейно, т. е. величина РП с расстоянием уменьшалась экспоненциально. Точно так же уменьшалась амплитуда электрического сигнала, который в специальных опытах с помощью микроэлектрода наносился на обнаженное нервное окончание вместо механического толчка (рис. 27). Такой характер изменения сигнала означает, что структуры немиелинизированной части нервного окончания в тельцах Пачини, ответственные за возникновение РП, не обладают регенеративными свойствами (не взаимодействуют между собой) и являются электрически невозбудимыми (подробнее см. стр. 275).
Величина РП зависит от раздражаемой площади нервного окончания (Loewenstein, 1961а), что естественно вытекает из данных, согласно которым возбуждение одного участка мембраны
 не
сказывается на соседних. При уменьшении
раздражаемой поверхности
не
сказывается на соседних. При уменьшении
раздражаемой поверхности
Рис. 27. Зависимость амплитуды рецепторного потенциала от расстояния до точки раздражения (по: Loewenstein, 1965). По оси абсцисс — расстояние между точкой раздражения и точкой регистрации ответов (мм); по оси ординат — амплитуда потенциалов (мв). Точки — амплитуда РП, вызванного механическим стимулом; крестики — амплитуда элек- тротонического потенциала, вызванного электрическим импульсом.
величина максимального РП снижается, характер же зависимости амплитуды РП от силы воздействия сохраняется. Используя понятие площади, эту зависимость можно охарактеризовать так: генераторный ток возрастает нелинейно, его рост замедляется с увеличением площади возбужденной мембраны (Loewenstein, 1961а, 1971).
Представление, согласно которому рецепторную поверхность следует рассматривать как «мозаичное» образование, составленное из отдельных элементов (конечно, в функциональном отношении, а не в прямом смысле этого слова), позволило провести рассмотрение деятельности рецептора методами математического моделирования (Loewenstein, 1961а; Ильинский, Фикс, 1963). При этом аналог каждого элемента функциональной «мозаики», ее элементарная ячейка (ею мог быть, например, отдельный канал через мембрану), наделялся следующими свойствами: он работал независимо от соседних элементов по принципу «все или ничего» и, следовательно, мог находиться либо в состоянии покоя, либо в состоянии возбуждения. Результаты теоретических расчетов и экспериментальные данные оказались в удовлетворительном соответствии. Однако следует признать, что сделанные оценки явля-
раздражения
имел симметричный

лись приближенными и основывались на моделях, носящих феноменологический характер.
При нанесении на тельце Пачини повторных раздражений небольшой частоты (один толчок в секунду и реже, так что взаимодействие между последовательными ответами отсутствовало) оказалось, что несмотря на строго постоянную величину механических стимулов, амплитуда и скорость нарастания РП варьировали (Ishiko, Loewenstein, 1959; Loewenstein, 1961а; Ильинский, Фикс, 1963; Ильинский, 1966а). Такие флуктуации параметров РП носили случайный, хаотический характер, и график распределения индивидуальных значений амплитуды РП для каждой силы
вид. Величина флуктуаций была весьма значительной и намного превышала обычные флуктуационные явления в нервном волокне (величина стандартного отклонения суб-
Рис. 28. Соотношение между уровнем длительного обусловливающего рецепторного потенциала (Fc) и параметрами быстрого тестирующего ответа (Vs) (по: Ozeki, Sato, 1965).
По оси абсцисс — уровень обусловливающего РП (мкв); по оси ординат: слева — амплитуда тестирующего РП (мкв), справа — время полуразрушения (мсек.)- Черные кружки — У , белые кружки — t. На врезке — схема измеряемых параметров РП.
максимального РП от среднего значения могла, например, достигать 8%). По мере увеличения интенсивности раздражения величина флуктуаций нарастала, а затем, достигнув максимума, снижалась, что объясняется нелинейным характером связи между амплитудой РП и силой раздражения: в области больших величин воздействия прирост амплитуды ответов уменьшается (Loewenstein, 1961а).
Явление депрессии второго РП, возникающего в разные периоды после начала первого ответа, исследовалось более подробно японскими авторами (Ozeki, Sato, 1965). Эти исследователи показали, что величина депрессии зависит от амплитуды первого РП. Если же опыт проводился на декапсулированном рецепторе и в ответ на первое раздражение достаточной длительности возникал определенный деполяризационный РП, то второй РП, появляющийся на фоне этой деполяризации, уменьшался не только по величине, но и по длительности (рис. 28). При этом укорачивалась длительность именно периода разрушения РП (время полуразрушения, оцениваемое по нисходящей фазе РП, уменьшалось на 1/4—1/5). Если учесть, что время разрушения РП определяется постоянной времени рецепторной мембраны, и считать, что емкость мембраны не претерпевает существенных изменений в ходе возбуждения, то можно заключить, что в период развития РП сопротивление мембраны значительно снижается, т. е. проницаемость мембраны для ионов увеличивается.
При раздражении тельца Пачини стимулами большой амплитуды с крутыми фронтами нарастания и спада можно было отметить появление множественных деполяризационных РП, каждый из которых мог вызвать появление ПД (Sato, 1961; Ильинский, 1965, 1966в). По мнению Сато, данные реакции обусловливаются свойствами рецепторной мембраны. Наряду с этой возможностью, которая действительно имеет место в тельцах Пачини (возникно-
отмечалось уменьшение^амплитуды РП (рис. 29). Величина й длительность этой посттетанической депрессии были тем больше, чем длительнее раздражался рецептор и чем больше была амплитуда и частота стимулов. При этом увеличение частоты с 1 до 200 стимулов в 1 сек. вызывало крутое снижение амплитуды РП. Максимум депрессии РП достигался при частоте 800 стимулов в 1 сек. (Loewenstein, Cohen, 1959b). Вместе с тем в части экспериментов величина РП после тетанизации увеличивалась. Такая посттетаническая потенциация наблюдалась у рецепторов, в ответах которых была мало выражена следовая негативность, а также можно было обнаружить явления гиперполяризации (например, следовую позитивность). Детально указанное явление не исследовалось, однако имеются основания полагать, что увеличение амплитуды РП может быть следствием гиперполяризационных сдвигов мембранного потенциала нервного окончания. Подробное рассмотрение гиперполяризационных ответов телец Пачини и причин их возникновения будет проведено в главе 10.
Антидромная ритмическая стимуляция рецептора (электрическое раздражение наносилось на афферентное нервное волокно) не вызывала, по данным одних авторов (Loewenstein, Cohen, 1959b), сколько-нибудь заметных последующих изменений в величине деполяризационного РП. По другим же данным (Sato, Ozeki, 1963), механический стимул, нанесенный сразу вслед за антидромным ПД, вызывал появление РП несколько меньшей амплитуды, чем в норме.
РП рецепторов по многим своим параметрам сходны с постсинаптическими потенциалами (ПСП). Явления посттетанического облегчения, в основе которых также лежат явления гиперполяризации, неоднократно отмечались при исследовании синапсов (см. обзоры: Hughes, 1958; Eccles, 1964). В этих же исследованиях было установлено, что при ритмическом раздражении синапсов наблюдаются и эффекты депрессии, которые носят более выраженный характер, чем в рецепторах. По мнению Экклса, происходит одновременное функционирование двух антагонистических факторов: кратковременной потенциации и более длительной депрессии. Из представленных данных следует, что сходные явления потенциации и депрессии достаточно хорошо проявляются и в механизме деятельности рецепторов, которые, однако, возбуждаются адекватными механическими стимулами.
Рецепторные потенциалы в мышечных веретенах
/
Хотя изучение физиологических особенностей данного типа рецепторов протекает весьма интенсивно, тем не менее следует признать, что литература содержит значительно меньшее количество фактов о функционировании этих сенсорных приборов, чем об их строении. И в первую очередь это касается особенностей локальных явлений, развертывающихся при раздражении мышечных веретен.
Впервые РП мышечных веретен был описан в опытах на лягушках (Katz, 1950b). Затем РП веретен был зарегистрирован у млекопитающих (котят) (Lippold et al., 1960а). В дальнейшем основные работы по изучению свойств РП проводились почти исключительно на лягушках (Ottoson, 1961, 1964, 1965а, 1965b; Ottoson, Shepherd, 1965, 1968, 1969, 1970b, 1970с, 1971b; Shepherd, Ottoson, 1965; Husmark, Ottoson, 1970, 1971b; см. также обзор: Ottoson, Shepherd, 1971a). В последнее время появились первые, пока еще предварительные данные и на изолированных веретенах у млекопитающих (Hunt, Ottoson, 1973). Так же, как и в случае телец Пачини, исследования РП веретен удобно проводить на рецепторах, импульсная активность которых блокирована новокаином или другим достаточно избирательно действующим на импульсную активность агентом.
По сравнению с деполяризационным РП тельца Пачини, РП мышечного веретена обладает существенным отличием: этот потенциал поддерживается в течение всего времени стационарного воздействия на рецепторы (рис. 21, Б). И в течение всего периода существования деполяризации рецептора возникает импульсная активность. Таким образом, мышечное волокно является структурой, прекращение активности которой (адаптация) развивается относительно медленно.
Хотя в настоящее, время нет еще прямых данных, говорящих о том, что РП генерируется именно в разветвлениях немиелинизированных нервных окончаний веретена, тем не менее практически все авторы склоняются к этой точке зрения. Так, например, локальное раздражение различных участков веретена с помощью микроиглы показывает, что основное афферентное нервное волокно и его миелинизированные разветвления являются относительно нечувствительными к механическому воздействию, в то время как полярные области веретена обладают высокой чувствительностью (Ottoson, Shepherd, 1971а). По мнению4* Катца (Katz, 1961), механические воздействия сказываются прежде всего па сенсорных колбах, которые погружены в интрафузальные мышечные волокна. Тонкие нервные волокна (порядка 0.15 мкм), соединяющие сенсорные колбы, по-видимому, выполняют роль только проводников. В результате электротонического распространения по немиелинизированным проводникам локальный ответ рецептора достигает мякотного нервного волокна, где и может быть зарегистрирован. Естественно, что при таком способе регистрации абсолютная величина РП в зоне его возникновения пока остается неизвестной.
В РП мышечного веретена, как и в локальных ответах других медленно адаптирующихся рецепторов, различают два компонента: динамический и статический. Первый определяется динамической составляющей раздражения, а второй — статической (Katz, 1950b). Для первичных окончаний динамическая фаза РП намного выше, чем для вторичных (Hunt, Ottoson, 1973). При снятии раздражения деполяризация рецептора сменяется кратковременной гиперполяризацией — возникает гиперполяризационный off-ответ (Katz 1950b; Lippold et al., 1960a; Hunt, Ottoson, 1973). У изолированных рецепторов эта гиперполяризация обычно наблюдается лишь после прекращения сильного раздражения (Ottoson, Shepherd, 1971а). Деполяризационный off-ответ, аналогичный описанному в тельцах Пачини, в мышечных веретенах не наблюдается. При нанесении на мышечное веретено стимулов большой продолжительности можно отметить некоторое снижение величины РП в ходе действия раздражителя. Этот факт, а также существование двух компонентов РП позволяет поставить вопрос о явлениях адаптации, возникающих в мышечном веретене при его раздражении (см. главу 4).
Как и в случае телец Пачини, предшествующие РП мышечных веретен могут оказывать угнетающее действие на последующие локальные ответы (Ottoson et al., 1969). Явление депрессии было тем больше и продолжительнее, чем больше была величина и длительность первого ответа. Этот эффект, по-видимому, определяется, с одной стороны, электрическими характеристиками рецепторной мембраны, а с другой — механическими свойствами веретена.
Временное течение разрушения РП зависит от скорости снятия растяжения, а также от длительности и силы воздействия (Ottoson, Shepherd, 1970а). Можно показать, что оно^ определялось как электрическими свойствами рецепторной мембраны и мембраны нервного волокна, расположенного между * областью генерации РП и местом его регистрации, так и механическими особенностями вспомогательного аппарата рецептора (скоростью возвращения его к своему исходному состоянию).
Увеличение интенсивности раздражения ведет к прогрессивному возрастанию амплитуды динамической и статической фаз РП (Katz, 1950b). При этом динамическая фаза ответа растет быстрее. Она достигает максимума (при достаточно высокой скорос1и раздражения) уже при удлинении рецептора на 30% относительно его длины в покое (рис. 30). В области больших амлитуд воздействия линейная зависимость между РП и раздражением нарушается. Общий ход изменения динамического компонента РП при варьировании величины раздражения напоминает кривую зависимости амплитуды РП телец Пачини от силы раздражения, что, возможно, говорит в пользу существования сходных механизмов генерации динамических ответов в этих механорецепторах.
Статический компонент РП во всем диапазоне растяжений линейно связан с амплитудой воздействия, монотонно возрастая при усилении раздражения (рис. 30), и может достигнуть тех же значений, что и динамическая реакция. Однако при этом потре
буется удлинение веретена более нем на 100%, которое приводит к необратимому повреждению рецептора (Ottoson, Shepherd, 1965). При перераздражении возможно появление платообразного динамического ответа, который начинает уменьшаться до конца стимула. Это находит отражение в ослаблении импульсного разряда, который в этот период прерывается (Husmark, Ottoson, 1970). Такого рода тормозная пауза может иметь место и в активности других рецепторов, например электрорецепторов, что говорит о достаточной универсальности рассматриваемого явления.
 При
оценке деятельности мышечных веретен
существенно знать влияние такого
параметра раздражения, как напряжение.
Из ра-
При
оценке деятельности мышечных веретен
существенно знать влияние такого
параметра раздражения, как напряжение.
Из ра-
Рис. 30. Зависимость между силой раздражения и амплитудой динамической и статической фаз РП мышечного веретена лягушки (по: Ottoson, Shepherd, 1971а).
По оси абсцисс — амплитуда растяжения (мкм); по оси ординат — амплитуда РП (мв). Черные кружки — динамический, белые — статический компонент ответа. Исходная длина покоящегося веретена — 0.9 мм.
бот старых авторов (Weber, 1846; Fechner, 1862) было известно, что самооценка испытуемым силы своего ощущения пропорциональна логарифму амплитуды раздражающего воздействия. Это хорошо было продемонстрировано Вебером у человека на примере механорецепции (опыты с использованием дозированных грузов, а также точно задаваемых звуковых стимулов). Именно эти данные заставили предположить существование такого рода зависимости между раздражителем и ответами сенсорных приборов. Электрофизиологические методы позволили изучить это явление в опытах на одиночных сенсорных приборах. На примере одиночного механорецептора — мышечного веретена лягушки — такого рода исследование было проведено Мэтьюзом (Matthews, 1931а), который действительно обнаружил, что частота разряда рецептора пропорциональна логарифму величины нагрузки (рис. 31, Б). В дальнейшем было показано (Katz, 1950b), что частота разряда линейно отражает стационарную фазу деполяризационного РП (рис. 31, Л). Таким образом, стало возможным утверждать существование линейной зависимости между логарифмом нагрузки, прилагаемой к мышечному веретену, и величиной стационарной рецепторной деполяризации мышечного веретена. Это удалось потом подтвердить и прямыми экспериментами (Hus- inark, Ottoson, 1970). В этой же работе была показана и экспоненциальная зависимость напряжения от длины веретена, т. е. такое же соотношение, какое существует й в целой мышце 1949; Matthews, 1959). Таким образом, интрафузальные мышечные волокна в данном отношении по своим свойствам существенно не отличаются от экстр афуз ал ьных.
Динамическая фаза рецепторного ответа (рис. 32) в значительной степени зависш от скорости нарастания раздражения (Katz, 1950b). При одной и той же амплитуде воздействующего стимула ее величина линейно связана с логарифмом скорости растяжения (Ottoson, Shepherd, 1965; Husmark, Ottoson, 1970). Скорость нарастания РП в ходе одиночного ответа прямо пропорциональна скорости растяжения в диапазоне менее 200 мкм/сек. (Katz, 1950b),

Рис. 31. Зависимость частоты разрядов мышечного веретена лягушки от величин рецепторного потенциала (4) и нагрузки (Б) (А — по: Katz, 1950b; Б — по: Matthews, 1931а).
По оси абсцисс': на А — амплитуда стационарной фазы РП (мв), на Б — логарифм величины нагрузки в граммах; по оси ординат — частота разрядов (ими./сек.).
Вопрос о происхождении различных фаз РП до настоящего времени не выяснен. Впервые Мэтьюзом (Matthews, 1931а) было высказано предположение, что различные области веретена обладают разными вязко-упругими свойствами, что может быть причиной появления динамического и статического компонентов ответа. Позднейшие исследования с помощью электронного микроскопа (Katz, 1961; Karlsson et al., 1966) действительно показали существование в веретене лягушки двух областей: ретикулярной и компактной. Наличие в ретикулярной зоне большого количества соединительнотканных элементов говорит об иных механических свойствах этой зоны по сравнению с компактной. Отсюда возникла гипотеза, что ретикулярная область веретена ответственна за появление динамической фазы РП, а компактная — за появление статической (Katz, 1961). Имеются факты, говорящие, что данное
объяснение приложимо и к веретенам млекопитающих (R. S. Smith, 1966; ср.: Hunt, Ottoson, 1973). Обзор данных в пользу указанной гипотезы дал Мэтьюз (Matthews, 1964, 1972). Тем не менее объяснить возникновение динамической составляющей ответа исключительно свойствами ретикулярной области, а статической — свойствами компактной области, по-видимому, нельзя (Ottoson, Shepherd, 1968, 1970а). Это следует, в частности, из экспериментов с избирательным разрушением отдельных частей веретена (Ottoson, Shepherd, 1971).

Рис. 32. Сопоставление динамической фазы рецепторного потенциала мышечного веретена лягушки (Л) со скоростью растяжения рецептора (/>) (по: Ottoson, Shepherd, 1970с).
По оси абсцисс — время (мсек.); по оси ординат: на А — амплитуда РП (мв), на Б — величина удлинения рецептора (мкм),
При обсуждении происхождения различных фаз РП мышечного веретена следует учесть и еще одно обстоятельство общего характера. Дело в том, что наличие динамической и статической фаз РП обнаружено в самых разных. сенсорных структурах, на' пример в механорецепторах разных типов: рецепторах растяжения у ракообразных (Eyzaguirre, Kuffler, 1955а; Loewenstein et al., 1963; Nakajima, Onodera, 1969b), волосковых рецепторах у насекомых (Thurm, 1965b; Smola, 1970b), фоторецепторах (Fuortes, Poggio, 1963; Stieve, 1965; Baumann, 1968; Fuortes, 1971), хеморецепторах (Tucker, Shibuya, 1965). Фазный и тонический компоненты импульсного разряда описаны для многих рецепторов, воспринимающих различные виды стимулов, в том числе для терморецепторов (Hensel, 1953, 1973), электрорецепторов (Murray, 1962, 1965а; Акоев, Ильинский, 1973; Акоев, Жадан и др., 1974), хеморецепторов (Beidler, 1961), рецепторов, воспринимающих болевые раздражители (Dodt, 1954), и т. д. Частота же импульсной активности в определенном диапазоне линейно связана с величиной РП.
Таким образом, можно считать, что динамическая и статическая фазы локального ответа рецептора являются весьма универсальным свойством РП вообще и вспомогательные структуры не могут быть единственным фактором, определяющим их возникновение. Можно предполагать, что за возникновение различных компонентов РП ответственны и какие-то фундаментальные свойства рецепторных мембран. Кроме того, фазно-тонический характер ответа может наблюдаться даже в некоторых нервных волокнах при действии на них постоянного электрического тока (Hodgkin, 1948). Следовательно, в формировании динамического и статического компонентов «узора» разряда могут определенным образом участвовать и элементы регенеративной системы генерации.
Характеристики РП различных типов мышечных веретен, а также первичных и вторичных нервных окончаний млекопитающих в настоящее время только начинают исследоваться (Hunt, Ottoson, 1973). Об особенностях РП пока обычно судят лишь по свойствам импульсных разрядов, которые будут рассмотрены ниже.
♦ ♦ *
Таким образом видно, что рецепторные потенциалы различных первичных механорецепторов имеют много сходных свойств. Основное же различие между ними заключается в том, что некоторые (фазные) механорецепторы (типа телец Пачини) могут генерировать РП лишь в период действия динамического компонента раздражения; другие же (тонические) рецепторы (типа мышечных веретен) могут генерировать РП также и в период действия статической фазы воздействия.
Деполяризационные рецепторные потенциалы механорецепторов органов чувств позвоночных
Хотя важность изучения локальных ответов волосковых рецепторов органов чувств несомненна, однако трудности методического характера являются причиной того, что в настоящее время они исследованы еще совершенно недостаточно. При этом, если в отношении суммарной активности рецептирующих клеток, выраженной в так называемых микрофонных потенциалах, в литературе имеются довольно многочисленные данные, то ответы одиночных волосковых рецепторов только начинают изучаться.
Микрофонные потенциалы впервые были обнаружены в кор- тиевом органе кошки (Wever, Bray, 1930), а затем в органах слуха других млекопитающих, в том числе и у человека (Гершуни и др., 1937). В дальнейшем они были описаны в различных структурах
лабиринта (Bleeker, Vries, 1949; Trincker, 1957; Furukawa, Ishii, 1967a, 1967b) и в органах боковой линии (Bleeker, Vries, 1949; Jielof et al., 1952; Kuijper, 1956 — цит. no: Flock, 1965). Они регистрируются при внеклеточном расположении отводящих электродов и в ряде случаев повторяют форму механического воздействия. Именно поэтому они в органе слуха вначале и получили название микрофонного эффекта улитки.
В отношении происхождения микрофонного потенциала в литературе существовали длительные споры. Некоторые авторы склонны были считать его даже чисто физическим явлением, т. е. не связанным со свойствами рецепторов сенсорных структур. Действительно, наличие раздела между двумя средами может при механическом воздействии привести к появлению феноменов, сходных с микрофонными потенциалами (Bekesy, 1960). Однако было показано, что такого рода причины могли бы быть ответственны только за часть ответа, наблюдаемого в органах чувств (в органе слуха примерно за 15—20% амплитуды ответа). Основная же часть микрофонных потенциалов несомненно отражает нормальную деятельность рецепторных элементов и именно те процессы, развитие которых приводит в конечном счете к возникновению возбуждения в афферентных нервных волокнах (подробнее о микрофонных потенциалах см: Bekesy, 1960; Davis, 1960, 1962, 1965; Tasaki, 1960; Hawkins, 1964; Flock, 1965a, 1965b, 1971a; Kat- suki, 1965; Wever, 1966; Молчанов, Радионова, 1972; Dallos, 1973). Полагают, что микрофонные потенциалы генерируются в области основания волосков, на апикальной поверхности рецептирующих клеток (Davis, 1960). Так как микрофонные потенциалы, как полагают сейчас (см.: Flock et al., 1973b), в основной своей части образованы за счет локальных электрических процессов, которые протекают в рецепторах при действии на них механических стимулов, то некоторые характеристики микрофонных ответов могут быть полезными для оценки свойств РП сенсорных структур органов чувств.
Микрофонные потенциалы возникают с небольшим латентным периодом (около 0.1 мсек.), что сходно с величиной скрытого периода РП других механорецепторов, например телец Пачини при регистрации локального ответа вблизи места возникновения (см. стр. 79). Микрофонные потенциалы развиваются без видимого порога, и величина их градуально возрастает по мере усиления раздражения, что также характерно для обычных РП. Частота микрофонных потенциалов может повторять частоту раздражающего воздействия, но может и отличаться от нее. Так, они следуют за частотой синусоидального стимула, наносимого на область ампулярной кристы (Bleeker, Vries, 1949), но отличаются от частоты раздражающих стимулов, удваиваясь при воздействии на структуры боковой линии (Flock, Wersall, 1962). Эта особенность микрофонных потенциалов обусловлена тем обстоятельством, что волосковые рецепторы являются асимметрично организован-
ними структурами: их киноцилия (или ее эквивалент — базальное тельце) расположена сбоку, асимметрично по отношению к стере- оцилиям (см. стр. 47). Оказалось, что волосковая клетка способна возбуждаться лишь при раздражении, направленном от стерео- цилий к киноцилии, и угнетаться при обратном направлении стимула (Lowenstein, Wersall, 1959). Важно, что знак генерируемого ею РП менялся при изменении направления воздействия: деполяризация сменялась гиперполяризацией. Амплитуда в последнем случае была, возможно, меньше, чем при деполяризации (Flock, 1965b).
В разных сенсорных образованиях ориентация клеток может" быть различной. Поэтому если в пределах данной сенсорной структуры все волосковые клетки однонаправлены (например, в crista ampullaris все киноцилии ориентированы одинаково), то микрофонные потенциалы воспроизводят частоту синусоидального воздействия. Напротив, если волосковые клетки ориентированы в разных направлениях, то микрофонные потенциалы, отражающие их суммарную активность, могли иметь иную частоту, чем раздражающий стимул. В органах боковой линии, где волосковые клетки распадаются на две группы, ориентированные в противоположные стороны, в ответной реакции наблюдается удвоение частоты раздражения как следствие суммации де- и гиперполяризационных сдвигов потенциала. Причем деполяризационные ответы преобладали над гиперполяризационными. Подробнее явление чустви- тельности механорецепторов к направлению действия раздражения, в основе которого могут лежать разные причины,, в том числе изменение знака РП, будет рассмотрено далее в главе 10.
Микрофонные потенциалы очень чувствительны к недостатку кислорода, и значительная их часть в органе слуха быстро исчезает после смерти животного (Bekesy, 1960). В органе слуха, где существует значительная постоянная разность потенциалов между отдельными частями сенсорного прибора (так называемый эндолимфатический, или эндокохлеарный, потенциал — подробнее о нем см. стр. 344), микрофонные колебания накладываются на этот постоянный потенциал, вызывая его модуляцию.
В самые последние годы начали появляться первые сообщения, касающиеся прямого микроэлектродного изучения свойств волосковых механорецепторов органов чуств (Harris et al., 1970; Flock, 1971a, 1971b; Flock, Russell, 1973; Flock et al., 1973a, 1973b; Mulroy et al., 1974; Weiss et al., 1974). В экспериментах Флока с соавторами на волосковых клетках боковой линии было установлено, что в покое мембранный потенциал варьирует от 10 до 65 мв. Сопротивление мембраны равно 10—100 мом (в среднем 40 мом). Имеется прямая зависимость между величиной мембрац- ного потенциала, обнаруженного в клетке, и сопротивлением мем-: браны. Волосковые рецепторы и опорные клетки имеют примернд одинаковые электрические параметры. При величине поверхности мембраны водорцрроц клетки примерно 1000 мкм2 удельцод gg-
н
противление мембраны равняется около 1000 ом/см2. Великина удельной емкости мембраны менее 0.3 мкф/см2 (Flock et al., 1973а, 1973b). Постоянная времени мембраны не превышает 0.2 мсек. Таким образом, волосковый рецептор обладает способностью отвечать на стимулы килогерцевого диапазона и средней интенсивности без амплитудных искажений. Вместе с тем механоре- цепторный орган боковой линии как целое имеет резонансную частоту в естественных условиях в районе 100 гц. Таким образом, частотное ограничение системы осуществляется не на уровне рецепторов, а за счет окружающих их элементов, взаимодействующих с водой.

![]() Волосковые
механорецепторы не способны отвечать
импульсной активностью. Они генерируют
только градуальные ответы. Волосковые
механорецепторы являются электрически
чевозбу-
Волосковые
механорецепторы не способны отвечать
импульсной активностью. Они генерируют
только градуальные ответы. Волосковые
механорецепторы являются электрически
чевозбу-
Рис. 33. Изменение рецепторного потенциала волосковой клетки боковой линии протея (Necturus macu- losus) в зависимости от направления смещения купулы (по: Flock, 1971а). Сверху вниз'. РП (внутриклеточная запись); отметка перемещения купулы относительно начального (н) положения. Отметка времени — 5 сек.; калибровка — 5 мв.
димыми (Flock et al., 1973b). Величина РП волосковых клеток сравнительно невелика. У рецепторов боковой линии она достигала лишь 1—2 мв (Flock et al., 1973а), а во внутреннем ухе — 3 мв (Weiss et al., 1974).
В работе Вейса и др. был зарегистрирован многокомпонентный рецепторный потенциал сенсорных волосковых клеток. Основной колебательный компонент ответа возникал с латентным временем 0.5 мсек., и его полярность изменялась при изменении полярности стимула. В опорных вспомогательных клетках были зарегистрированы ответы, аналогичные некоторым компонентам реакции волосковых клеток. Наличие таких ответов в опорных клетках позволяет предположить существование электрической связи между волосковыми и опорными клетками и дает возможность славить вопрос о латеральном электрическом взаимодействии между этими клетками. (У рецепторов же боковой линии если эта связь и существует, то в небольшой степени: Flock et al., 1973а). I (олярность основного начального компонента ответа рецепторных структур может быть различной у разных клеток, что, по-видимому, обусловлено различной ориентацией волосковых рецепторов по отношению к направлению действия стимула.
Зависимость полярности ответа от направления раздражения особенно хорошо была видна в опытах на волосковых клетках боковой линии протея (рис. 33). При этом изменение положения рецепторов относительно истопника раздражения на 1806 меняло полярность ответа данной клетки на противоположную (Harris et aL, 1970; Flock et al., 1973b). Таким образом, гипотеза о зависимости характера ответов волосковых механорецепторов органов чувств от их ориентации по отношению к направлению действия раздражения получила в настоящее время прямое подтверждение в эксперименте.
Как следует из морфологических данных, между волосковой клеткой и окончаниями сенсорного нейрона имеется синаптическая связь, причем пресинаптическим элементом является волосковая клетка. С помощью микроэлектродов удалось зарегистрировать возбуждающие постсинаптические потенциалы (ВПСП) немиелинизированных окончаний сенсорного нейрона (см., напр., в главе 5 рис. 81) (Flock, 1973; Flock et al., 1973b). В покое наблюдается спонтанное возникновение ВПСП, что, возможно, обусловлено спонтанным выделением медиатора из волосковой клетки. При нанесении раздражения на рецептирующую клетку отмечается синхронизация ВПСП афферентных волокон со стимулами (см. в главе 3 рис. 59).
Природа медиатора, переносящего возбуждение с волосковых клеток на афферентные волокна, еще неясна. В последнее время показано, что медиатор в синапсах волосковых клеток с афферентными окончаниями иной, чем в синапсах этих же клеток с эфферентными волокнами (Flock, 1973). Так как эфферентными волокнами выделяется, по-видимому, ацетилхолин (см. об этом более подробно в главах 5 и 8), то следует заключить, что волосковые клетки выделяют другое вещество.
Деполяризационные рецепторные потенциалы механорецепторов у беспозвоночных
Рецепторные потенциалы в рецепторах растяжения у ракообразных
С момента открытия этих механорецепторов (А1е- xandrowicz, 1951) они и по сей день являются одним из основных объектов, на которых проводится изучение свойств рецепторов и нервных клеток вообще и механорецепторов в частности. Крупный размер этих нейронов и их локализация позволяют вводить в них микроэлектроды под контролем микроскопа. Фактически до самого последнего времени это был единственный вид механорецепторов, у которого свойства РП исследовались не только с помощью внеклеточных записей, но и при внутриклеточных отведениях.
Изучение свойств РП рецепторов растяжения было осуществлено в классических работах 50-х годов (Eyzaguirre, Kuffler, 1955а, 1955b), за которыми последовали исследования и других авторов (Krnjevic, Gelder, 1961; Terzuolo, Washizu, 1962; Edwards ot al., 1963; Wendler, 1963; Nakajima, 1964; Obara, Grundfest, 1968; Nakajima, Onodera, 1969b).
При растяжении мышечного рецепторного органа (МРО) возникает деполяризация сенсорного нейрона (рис. 21, Л), которая является РП данного механорецепторного прибора (Eyzaguirre, Kuffler, 1955а). РП генерируется в дендритных разветвлениях под влиянием деформации, оказываемой мышечными волокнами. От места возникновения РП электротонически распространяется на тело сенсорного нейрона и отходящий от него аксон — нервное волокно, где и возникают ПД (Edwards, Ottoson, 1958). Величина

Рис. 34. Характеристика рецепторных потенциалов и импульсной активности медленно адаптирующегося рецептора растяжения у ракообразных (по: Terzuolo, Washizu, 1962).
А — зависимость РП от величины растяжения мышечного пучка у рака. По оси абсцисс — длина мышечного пучка (мм); по оси ординат — амплитуда РП (мв). Б, В — зависимость частоты импульсной активности от величины нагрузки и растяжения мышечного пучка у лангуста. По оси абсцисс: на Б — логарифм величины нагрузки в граммах; на В — длина мышечного пучка (мм); по оси ординат — частота импульсной активности (имп./сек.).
РП градуально возрастает при увеличении раздражения (Eyzaguirre, Kuffler, 1955а). Она связана линейной зависимостью с длиной мышечных волокон рецептора, а также с логарифмом приложенной нагрузки, так как между величиной растяжения рецепторной мышцы и логарифмом напряжения существует прямая зависимость (рис. 34) (Terzuolo, Washizu, 1962). По другим данным (Nakajima, Onodera, 1969b), между растяжением и амплитудой РП линейная зависимость существует лишь в небольшом диапазоне величин РП (10—20 мв), в целом же кривая имеет S-образный вид.
Амплитуда РП оказалась прямо пропорциональной величине мембранного потенциала клетки (Terzuolo, Washizu, 1962). Определение величины равновесного потенциала для РП не может считаться завершенным. По одним данным (Terzuolo, Washizu, 1962), он лежит в пределах от —10 до 0 мв; по другим (Obara, 1968) — в пределах от —20 до —30 мв. Согласно же данным экспериментов с фиксацией потенциала (Klie, Wellhdner, 1973), он имеет положительное значение (порядка +25 мв), хотя точную его величину авторы определить не смогли.
Измерение электрических характеристик рецепторных нейронов в покое показало, что средний мембранный потенциал МРОХ равнялся —50 мв (Terzuolo, Washizu, 1962); по данным других авторов, он колебался от —70 до —80 мв (Eyzaguirre, Kuffier, 1955а; ОЬага, 1968; Nakajima, Onodera, 1969а; Klie, Wellhdner, 1973). Постоянная времени мембраны MPOi составляет около 14— 15 мсек. (Terzuolo, Washizu, 1962); по другим данным — 13.3 мсек, для МРОХ и 10.4 мсек, для МРО2 (Nakajima, Onodera, 1969а). Сопротивление мембраны варьирует от 3.1 до 4.0 мом (Terzuolo, Washizu, 1962; Nakajima, Onodera, 1969a). Общая емкость мембраны равняется (3.4—3.2)-10"9 ф (Nakajima, Onodera, 1969а).
При растяжении рецептора электрические параметры изменялись параллельно величине воздействия. При максимальном раздражении (перерастяжении) мембранный потенциал снижался в среднем до 24 мв, постоянная времени — до 7—8 мсек., сопротивление— до 1.5 мом (Terzuolo, Washizu, 1962). Соответственно при уменьшении растяжения все величины изменялись в меньшей степени. Таким образом, можно считать, что при генерации РП проницаемость мембраны возрастает.
Как показали опыты, прежде всего увеличивается проницаемость для ионов натрия, которые в первую очередь и являются ответственными за генерацию деполяризационного РП (Edwards et al., 1963).
При достаточно быстром нанесении раздражения форма РП практически не отличается от РП, описанного у мышечного веретена: вначале возникает динамическая фаза ответа, за которой следует статическая фаза. При снятии раздражения может иметь место гиперполяризационный off-ответ, который более выражен у предварительно растянутых рецепторов (Nakajima, Onodera, 1969b).
Уже самые первые электрофизиологические исследования (Wiersma et al., 1953; Kuffier, 1954; Eyzaguirre, Kuffier, 1955a, 1955b) показали, что рецепторы растяжения ракообразных могут быть разделены на два типа: медленно и быстро адаптирующиеся (МРОХ и МРО2). Первые отвечали в течение всего периода действия растяжения, а также часто обладали фоновой ритмической активностью. Вторые же, напротив, отвечали всплеском импульсной активности в момент включения раздражения и затем обычно прекращали генерацию спайков. Однако после блокирования ПД и исследования одних лишь РП было установлено, что амплитудные характеристики РП обоих типов рецепторов практически не отличались друг от друга и их временное течение было сходным. Следовательно, различия между двумя типами рецепторов опре
деляются спаикогенерирующими структурами, а не свойствами нерегенеративных систем, определяющих возникновение РП.
При одной и той же величине растяжения РП у МРОХ был больше, чем РП, регистрируемый в МРО2 (примерно в 2 раза). Однако если оценивать величину растяжения по отношению к исходной длине (а размер покоящейся РМ2 больше, чем PMJ, то следует признать, что чувствительность обоих МРО была сходной (Naka- jima, Onodera, 1969b). Аналогичный результат был получен и при измерении чувствительности по стимулирующему напряжению. 11 |)и оценке, основанной на учете лишь величины усилия, действующего на РМ, показатель чувствительности MPOj оказывается в 5 раз больше, чем аналогичный показатель для МРО2. Однако поперечное сечение РМХ значительно меньше, чем РМ2. Удельная же величина напряжения (усилия на единицу поперечного сечения РМ) была в обоих случаях примерно одинаковой. Именно это дало основание утверждать, что одно и то же воздействие на дендриты PKt и РК2 приводит к генерации примерно одинакового РП (Na- kajima, Onodera, 1969b).
Рецепторные потенциалы
в кутикулярных волосковых рецепторах
насекомых
(Wolbarsht, 1960; Thurm, 1964а, 1965b; Smola, 1970b). В этих
работах было установлено, что РП
чувствительных

Рис. 35. Деполяризационный рецепторный потенциал чувствительной волосковой клетки насекомых (А — по: Thurm, 1965b; Б — по: Smola, 1970b).
 Л
— сенсилла у пчелы (1,2
—
два примера регистрации при разной
скорости развертки). Сверху
вниз: PH
(записи с наложением); угол наклона
волоска. Ступенчатое изменение, на
восходящей фазе РП — отражение
возникновения первого НД. Отметка
времени: на 1
—
10, на
2
—
100 мсек.; калибровка — Ю мв. В
—
ветрочувствительные рецепторы у
азиатской саранчи. Сверху
вниз:
РП; период стимуляции (черная
линия).
Угол отклонении волоска — 17.3°. Отметка
времени — 1 сек.; калибровка — 10
МВ.
Л
— сенсилла у пчелы (1,2
—
два примера регистрации при разной
скорости развертки). Сверху
вниз: PH
(записи с наложением); угол наклона
волоска. Ступенчатое изменение, на
восходящей фазе РП — отражение
возникновения первого НД. Отметка
времени: на 1
—
10, на
2
—
100 мсек.; калибровка — Ю мв. В
—
ветрочувствительные рецепторы у
азиатской саранчи. Сверху
вниз:
РП; период стимуляции (черная
линия).
Угол отклонении волоска — 17.3°. Отметка
времени — 1 сек.; калибровка — 10
МВ.
волосковых клеток насекомых имеет хорошо выраженную динамическую и статическую фазы (рис» 35). Амплитуда ответа градуально возрастает по мере увеличения раздражения (угла наклона волоска). Для динамического компонента ответа эта зависимость имеет S-образный вид, в то время как статическая составляющая РП меняется при изменении стимула более линейно.
Латентный период РП равняется примерно 0.1 мсек. В части случаев он может быть больше, но не превышает 0.2 мсек. При выключении раздражения могут возникать гиперполяризационные сдвиги потенциала. Возникающий РП приводит к уменьшению ответа на второй стимул (Thurm, 1965b). Снижение амплитуды динамической фазы второго РП наблюдается как в ходе генерации первого РП, так и после него.
Таким образом, можно заключить, что РП волосковых кутикулярных механорецепторов беспозвоночных по своим основным характеристикам практически не отличается от РП других механорецепторных приборов.
Рецепторные потенциалы в волосковых механорецепторах статоциста
В последнее время появились данные о РП волосковых чувствительных клеток статоциста моллюска Hermissenda crassicornis (Alkon, Bak, 1973). При внутриклеточном отведении был зарегистрирован деполяризационный РП, возникавший в ответ на механическое раздражение со скрытым периодом, менее 2 мсек. Величина деполяризационного РП могла достигать 40 мв. В определенном диапазоне раздражений величина РП была линейно связана с логарифмом амцлитуды смещений. Она зависела от величины мембранного потенциала клетки. Потенциал реверсии равнялся 30—35 мв. В период возникновения деполяризационного РП (генераторного потенциала) проводимость клеточной мембраны для ионов возрастала. (В покое сопротивление мембраны варьировало от 30 до 100 мом). Генераторный потенциал вызывал появление импульсной активности, частота которой линейно связана с величиной локальной деполяризации.
При ритмическом раздражении статоциста в ответ на каждый краткий стимул возникал РП. При увеличении частоты воздействия до 10 в 1 сек. отдельные РП могли суммироваться. При более высоких частотах раздражения ответы вначале суммировались, а затем прогрессивно уменьшались по величине. Авторы полагают, что волосковая клетка может отвечать на смещение независимо от его скорости. Адекватным стимулом они считают сгибание сенсорного волоска вне зависимости от скорости этого процесса.
Таким образом, можно заключить, что деполяризационный РП и этого механорецепторного прибора в целом сходен с аналогичными ответами других рассмотренных ранее рецепторов.
* * *
Существующие в настоящее время данные позволяют говорить о принципиальном сходстве основных особенностей рецепторных потенциалов самых разных механорецепторов позвоночных и беспозвоночных животных вне зависимости от того, являются ли эти механорецепторные приборы первичными или вторичными. Этими основными чертами РП являются прежде всего градуальный характер возникновения и способность к суммации. РП возникают с кратким латентным периодом, позволяющим говорить об отсутствии промежуточного медиаторного механизма между механическим стимулом и процессом генерации РП. РП тонических рецепторов имеет динамические и статические фазы, а у фазных рецепторов наблюдается только динамический компонент ответа.
Как у первичных, так и у вторичных механорецепторов отмечается наличие гиперполяризационных рецепторных потенциалов. Имеется связь этих потенциалов с направлением действия раздражителя. Этот вопрос более подробно будет рассматриваться в главе 10.