

Глава 3
ИМПУЛЬСНАЯ АКТИВНОСТЬ И ЕЕ ОСОБЕННОСТИ
У РАЗЛИЧНЫХ МЕХАНОРЕЦЕПТОРОВ
Систематическое изучение деятельности рецепторов с помощью электрофизиологических методов началось с регистрации импульсной активности афферентных нервов. Этот методический прием оказался столь плодотворным, что и по сей день является одним из важнейших в исследовании работы сенсорных приборов. Естественно, что со временем его возможности значительно расширились. Так, сейчас исследователи работают на уровне одиночных элементов. При этом импульсная активность изучается не только у животных в острых и хронических экспериментах, но и у человека (Hensel, Boman, 1960; Vallbo, Hagbarth, 1967, 1968; Hagbarth, Vallbo, 1968, 1969; Hagbarth et al., 1970; Knibestol, Vallbo, 1970; Vallbo, 1970a, 1970b, и др.). Это позволяет сопоставить регистрируемую электрическую активность с субъективными ощущениями. Важным шагом вперед является также широкое применение вычислительной техники при анализе импульсной активности.
Интенсивное изучение импульсной активности рецепторов определяется в настоящее время двумя основными причинами. Во-первых, оказалось, что именно импульсная последовательность является тем основным «языком», с помощью которого различные части нервной системы общаются между собой (Adrian, 1928, 1932). Поэтому очень важно знать, каким образом те или иные признаки раздражителя, падающего на организм, кодируются в последовательность импульсов, направляющихся от рецепторов в центральную нервную систему. Во-вторых, в очень многих случаях по чисто техническим причинам не представляется возможным регистрировать рецепторные потенциалы, и импульсная активность является основным источником информации о процессах, протекающих в сенсорных приборах при действии на них раздражителей. В силу этих причин импульсная активность изучалась значительно больше, чем локальные электрические явления.
В этой главе представлены данные об импульсной активности многих видов механорецепторов. Импульсная активность других (специализированных) механорецепторных приборов (органа слуха и вестибулярной системы) была подробно рассмотрена в специальных детальных обзорах, уже включенных в другие тома данного руководства. Материал главы расположен ц соответствии с использующимися принципами классификации механорецепторов. Изложение начинается с анализа некоторых общих характеристик импульсной активности и обсуждения вопроса о функциональной роли отдельных параметров импульсации. Далее приводятся данные об импульсации от механорецепторов кожи, об активности механорецепторов внутренних органов и сердечно-сосудистой системы, опорно-двигательного аппарата; кратко резюмируются материалы по импульсной активности механорецепторов органов чувств позвоночных. И, наконец, рассматриваются данные об активности некоторых видов механорецепторов у беспозвоночных.
Некоторые особенности изложения в существенной мере зависели от двух дополнительных обстоятельств. Первое из них касается телец Пдчини. Детальная характеристика импульсной активности пачиниевых телец приводится здесь в начале изложения, в разделе, посвященном кожным рецепторам, хотя основной экспериментальный материал по этому вопросу был получен-на тельцах Пачини, расположенных в брыжейке, т. е. на интероцецторах. Как известно, тельца Пачини широко распространены в организме — в коже, внутренних органах, опорно-двигательном аппарате. Поэтому их можно было бы рассматривать в любом из соответствующих разделов. В этой главе все важнейшие данные об импульсации телец Пачини рассмотрены с самого начала и сразу в одном месте.
Второе обстоятельство касается того очевидного факта, что особенно важно было изложить наиболее подробно данные об импульсной активности тех механорецепторов, в которых был детально исследован и рецепторный потенциал, так как именно это условие дает возможность оценить зависимость характеристик импульсного разряда сенсорного прибора от свойств его локальной электрической активности.
ОБЩАЯ ХАРАКТЕРИСТИКА ИМПУЛЬСНОЙ АКТИВНОСТИ
На основании многочисленных исследований сейчас можно утверждать, что возникновению импульсной активности в рецепторах самых разных типов всегда предшествует развитие в них локальной деполяризации (генераторного потенциала — ГП), которая появляется в результате действия адекватного раздражения (см. обзоры: Granit, 1955; I. А. В. Gray, 1959b; Davis, 1961; Grundfest, 1961b, 1964, 1971; Ильинский, 1966д, 1967, 1972a; Mellon, 1968; Fuortes, 1971; Tamar, 1972, и др.). Достигнув определенного критического уровня (firing level), ГП (точнее лежащие в его основе ионные токи) обусловливает возникновение ПД. В первичных рецепторах функцию генераторного потенциала выполняет деполяризационный РП, а во вторичных — возбуждающий постсинаптический потенциал (ВПСП). Простая констатация существования этих явлений приводит к естественному заключению, что возникновение импульсной активности в рецепторах должно происходить по тем же законам, по каким возникают потенциалы действия в нервном волокне (или клетке) под влиянием электрического тока. При этом, конечно, могут отмечаться и известные особенности, определяемые рядом причин: наличием вблизи места генерации ПД электрически невозбудимых или маловозбудимых мембран, существованием многочисленных разветвлений нервных волокон, образующих сенсорные терминали и т. д. Однако в целом возникновение импульсной активности в рецепторах не может быть хорошо понято без учета известных положений общей физиологии, относящихся к механизмам генерации потенциалов действия в нервных элементах. Заметим далее, что генерация импульсной активности во вторичных рецепторах определяется теми же правилами, что и в других постсинаптических образованиях.
Рассмотрение механизмов возникновения потенциалов действия в миелинизированных и немиелинизированных нервных волокнах и в нервных клетках под влиянием электрических стимулов, равно как и механизмов синаптической передачи, далеко выходит за рамки данной книги. Эти вопросы подробно освещены в ряде специальных сводок (Hodgkin, 1951, 1958, 1964; Eccles, 1957, 1964; Tasaki, 1959а; Костюк, Шаповалов, 1964; Шаповалов, 1966; Katz, 1966; Cole, 1968; Ходоров, 1969), в том числе в томах данного руководства (Аршавский и др., 1969; Либерман, Чайлахян, 1969; Шаповалов, 1969).VII Следует, однако, заметить, что если механизм генерации единичного потенциала действия сейчас изучен весьма детально, то о природе возникновения ритмической импульсной активности известно еще недостаточно (Fitzhugh, 1961; Hodgkin, 1964; Frankenhaeuser, Vallbo, 1965; Tomita, Wright, 1965; Cooley, Dodge, 1966; Ходоров, 1969; Shapiro, Lenherr, 1972). А именно этот тип активности представляет наибольший интерес для физиологии рецепторов.
Для понимания механизма осуществления ритмической активности большое значение придается такому феномену, как аккомодация. Исследователи давно отметили важную роль крутизны нарастания электрического тока при его действии на возбудимые элементы. Ниже некоторого критического значения скорости нарастания стимулирующего тока (минимального градиента) возбуждение вообще не наблюдается. Явление увеличения порогов при снижении скорости постепенного нарастания тока получило наименование аккомодации (Nernst, 1908) (см. обзор литературы: Аршавский и др., 1969; Ходоров, 1969).
Возбудимые структуры приспосабливаются к стимулу в ходе его длительного нарастания, происходит своего рода «вкрадыва- ние» раздражителя. Сходный эффект наблюдается и при действии на нервные проводники постоянного тока. В этом случае возбудимость проводников по мере действия длительного стимула постепенно падает; они (с различной скоростью) перестают реагировать на действие тока. Таким образом, возникает адаптация к постоянно действующему раздражителю.
Следует заметить, что термин «адаптация» в биофизике возбудимых мембран и в физиологии рецепторов употребляется в разных смыслах. В последнем случае этот термин охватывает большую группу явлений. Обычно адаптацией сенсорного прибора называют ослабление его импульсного ответа в ходе действия стационарного адекватного воздействия. Это означает, что явления адаптации рецепторов могут определяться: 1) процессами, протекающими в их вспомогательных аппаратах (и окружающих тканях); 2) процессами, лежащими в основе первичных трансформационных явлений, а также связанных с ними явлений генерации ГП; 3) процессами, определяющими реакцию регенеративных элементов рецептора (часто в этом случае речь идет о его нервном волокне) на действие деполяризационных токов, обусловливающих развитие ГП (подробнее явления адаптации в рецепторах будут рассмотрены в главе 4). Очевидно, что лишь последняя группа явлений, развивающихся в рецепторах, совпадает по смыслу с термином «адаптация», используемом в литературе, посвященной биофизике возбудимых мембран. Известно, что между аккомодацией возбудимых образований и их способностью уменьшать частоту генерируемых повторных разрядов (т. е. адаптационной способностью в биофизическом смысле слова) существует тесная связь (Katz, 1936, 1939; Granit, Skoglund, 1943; Sato, 1961, и др.).
Чем выше аккомодационная способность нервных проводников, тем в меньшей степени способны они генерировать повторные импульсные ответы. У сенсорных волокон, дающих весьма длительные последовательности импульсов в ответ на действие постоянного тока, скорость аккомодации была существенно ниже, чем у не обладавших такой особенностью моторных волокон (Skoglund, 1942; Granit, Skoglund, 1943). Величина минимального градиента для сенсорных волокон лягушки была ниже, чем для моторных (соответственно 89 + 13 и 244 + 14 реобаз/сек.) (Bergman, Stampfli, 1966). Па мякотных волокнах лягушки было показано существование линейной зависимости между логарифмом минимального градиента и числом ПД в разряде (Vallbo, 1964а). Отсюда понятно, почему влияния, снижающие аккомодацию (в случае рецепторов таким фактором может быть, например, гиперполяризационный РП), увеличивают способность нервных волокон генерировать ритмические ответы, а противоположные влияния — ее уменьшают.
В связи с изложенным выше мы используем в книге термин «адаптация» только в смысле, принятом в физиологии сенсорных систем, т. е. прежде всего для характеристики процессов, протекающих в целом рецепторном приборе. Способность же собственно регенеративной системы генерации сенсорного прибора реагировать меньшей частотой импульсов на действие длительного деполяризующего тока (в период существования статической фазы ГП) будет определяться в понятиях аккомодации (об адаптации и аккомодации см. также: Granit, 1955).
Обычно после возникновения ПД сопротивление возбудимой мембраны некоторое время остается пониженным (задержанное выпрямление, обусловленное повышенной калиевой проводимостью), в то время как критический уровень деполяризации является повышенным (за счет частичной инактивации натриевой системы). Оба этих фактора, по-видимому, могут иметь значение при ослаблении реакции нервных волокон на действие постоянно деполяризующего тока (Frankenhaeuser, Vallbo, 1965; Shapiro, Lenherr, 1972).
Реакция разных нервных волокон на действие постоянного тока представлена на рис. 36. Именно такого характера рисунки разрядов типичны
также для медленно и быстро адаптирующихся рецепторов (например, МРО и МРО2 у ракообразных).
При изучении деятельности рецепторов было установлено, что чем больше амплитуда генераторного потенциала, тем выше ча-

Рис. 36. Действие постоянного тока на немиелинизированные (Л) и миелинизированные (Б) нервные волокна (А ~ по: Tomita, Wright, 1965; Б — по: Vallbo, 1964b).
А — ритмические разряды нервного волокна краба. Сверху вниз', отметка раздражения; импульсная активность. На 2 и 3 пунктирные линии — нулевой уровень потенциала и изменение критического уровня деполяризации в ходе ритмического разряда. Длительность постоянного деполяризующего тока — 300 мсек.; сила тока увеличивается в каждом кадре от 1 до 12 на 0.5-10-8 а (от едва надпороговой — 1• 10~8 — до 6.5-10-8 а). Б — ответы одиночного перехвдта Ранвье нервного волокна лягушки. Сверху вниз: мембранный потенциал; амплитуда деполяризующего тока (от 1 до 6 нарастает).
стота импульсных разрядов сенсорного прибора (см. рис. 31) (Katz, 1950b; Eyzaguirre, Kuffler, 1955a; Fuortes, 1959; Rushton, 1959; Terzuolo, Washizu, 1962, и др.). Однако это справедливо только до определенного предела ГП, выше которого наступает угнетение и даже полное выключение импульсной активности (сравн. рис. 36). В зависимости от аккомодационных особенностей регенеративной системы генерации блокирование импульсной активности может возникнуть как при небольших величинах и длительностях действия локальной деполяризации (например, в тельцах Пачини), так и при весьма значительных амплитудах (см. подробнее главу 4) и продолжительности стационарной деполяризации (например, в рецепторах растяжения у ракообразных).
Если рецептор в течение продолжительного времени генерирует импульсную активность, особенно при достаточно высокой частоте импульсации в ответе, то часто после выключения раздражения наблюдается посттетаническое угнетение (например, в рецепторах растяжения у ракообразных,,.— Nakajima, Takahashi, 1966). При ритмическом прерывистом раздражении рецепторов изменение импульсной активности происходит по тем же правилам, как и в других возбудимых образованиях, т. е. так же существуют оптимальные и пессимальные для возбудимого субстрата частоты раздражения, наблюдается явление трансформации ритма и т. д. (Loewenstein, 1958; Ильинский, 1966а, и др.).
За исключением некоторых чрезвычайно быстро адаптирующихся нервных окончаний, для рецепторных приборов является очень характерной генерация ритмического разряда импульсов в ответ на действие раздражителя. Только варьированием показателей ритмического разряда большинство рецепторов может информировать нервную систему о различных параметрах внешнего воздействия. Поэтому ритмическую активность можно считать одной из фундаментальных особенностей деятельности рецепторных приборов, в том числе и механорецепторов.
Изучение природы ритмической импульсации в рецепторах было начато в классических работах Эдриана (Adrian, 1928, 1932) и в дальнейшем продолжалось в двух основных направлениях, посвященных исследованию механизмов аккомодации нерва и механизмов, определяющих интервал между спайками. В физиологии сенсорных систем для объяснения механизмов, определяющих величину межспайковых промежутков, т. е. частоту разряда, первоначально было выдвинуто представление о ведущей роли изменений рефрактерности субстрата после возникновения импульса (Adrian, 1928). Однако с позиций этой гипотезы было трудно объяснить низкочастотную активность рецепторов. В этом случае меж- спайковый интервал был много длительнее времени полного восстановления возбудимости.
Эксперименты на нервных волокнах ракообразных позволили выдвинуть представление, согласно которому интервал между спайками в импульсной активности определяется теми же факторами, что и латентное время, необходимое для возникновения первого импульса в ответ на включение раздражения (Hodgkin, 1948). Оба эти промежутка времени определялись динамикой локального ответа: в момент, когда последний достигал критического уровня, возникал спайк. Эта гипотеза хорошо объясняла характер изменения частоты при низких силах раздражения. При больших же интенсивностях раздражающих воздействий частота разряда зависела скорее от рефрактерного периода, так как латентный период реакции мог быть в этом случае очень малым, а межспайковый интервал никогда не был менее нескольких миллисекунд.
Эксперименты на мышечном веретене (Katz, 1950а; Ottoson, Shepherd, 1971а) и на рецепторе растяжения ракообразных (Еу- zaguirre, Kuffler, 1955а) в целом подтвердили изложенные выше представления о механизмах генерации импульсной активности в рецепторах. В настоящее время при анализе рассматриваемых явлений очень широко используются представления общей физиологии, о которых было упомянуто выше.
Важное значение при обсуждении механизмов генерации импульсной активности в рецепторах придается месту возникновения ПД. Этот вопрос применительно к механорецепторам неоднократно поднимался в литературе (см. обзоры: J. А. В. Gray, 1959а; Edwards, 1960; Loewenstein, 1971). Как было видно из главы 2, свойства локальных электрических ответов рецепторов (РП) существенно отличаются от импульсной активности (ПД). Именно это обстоятельство и привело к употреблению в физиологии рецепторов терминов «нерегенеративная» и «регенеративная системы генерации» (см. стр. 76). Было важно выяснить, являются ли эти системы пространственно разделенными или же они обеспечиваются структурами одной и той же территории. Хотя в целом данный вопрос в настоящий момент и не может считаться окончательно выясненным, все же можно, по-видимому, заключить, что в каждом рецепторном приборе имеется область, наиболее благоприятная для возникновения распространяющейся импульсной активности, где и происходит генерация ПД в обычных условиях. Такова, например, область сужения аксона вблизи от сомы рецепторного нейрона растяжения у ракообразных (Edwards, Ottoson, 1958; Nakajima, Onodera, 1969a), область начала мякотного волокна в тельцах Пачини (Diamond et al., 1956; J. А. В. Gray, 1959а; Loewenstein, 1971; Ильинский, Волкова и др., 1974), основание дендрита в механорецепторных сенсиллах у насекомых (Wolbarsht, 1960; Thurm, 1967, 1970) и т. д. С другой стороны, говорить о наличии какого-то абсолютного резкого пространственного разграничения нерегенеративной и регенеративной систем генерации, по-видимому, нельзя.
Для многих рецепторных структур, например таких, как волосковые механорецепторы полукружных каналов и боковой линии, ряда рецепторов растяжения в легких и в других тканях, некоторых хемо- и электрорецепторов и т. д., характерно наличие фоновой спонтанной активности. Она имеет важное значение в деятельности сенсорных систем, например, позволяет центрам с помощью одних и тех же рецепторов оценивать действие как возбуждающих, так и тормозных раздражителей (см. главу 10). Фоновая импульсация поддерживает также высокую возбудимость центральных образований.
Спонтанная импульсная активность отражает ряд важных особенностей в деятельности рецепторных приборов. Она, по-видимому, связана с флуктуационными изменениями мембранного потенциала, спонтанно происходящими в рецепторных клетках, что находит свое отражение в колебаниях амплитуды РП. Аналогичные изменения потенциала хорошо известны и в различных других нервных структурах (Verveen, 1961; Derksen, 1965; Firth, 1966; Verveen, DeFelice, 1974). Для того чтобы спонтанные изменения ГП могли вызвать появление импульсных разрядов, порог возбуждения регенеративной системы рецепторов должен быть достаточно низким, что и наблюдается в действительности. Чрезвычайно низкие пороги возникновения возбуждения, характерные практически для всех высокоспециализированных рецепторов (например, в органах чувств), а также наличие во многих из них спонтанной активности заставляет предполагать постоянное неустойчивое состояние всех возбудимых систем этих сенсорных приборов (Ильинский, 1967).
Необходимо отметить, что чувствительность сенсорных образований к действию адекватного раздражения, порог их возбуждения является одной из важнейших характеристик рецепторных приборов. Порогом чувствительности рецептора на основании электрофизиологических измерений следует считать ту минимальную силу воздействия, которая необходима для того, чтобы появился первый ПД. Как показывает опыт исследований, определить абсолютный порог чувствительности механорецепторов к действию адекватного стимула в прямых электрофизиологических исследованиях часто бывает сложно. Поэтому в ряде случаев величина порога оценивается на основе психофизических измерений или поведенческих реакций, а в некоторых случаях и по характеру реагирования висцеральных и других систем. Очевидно, что полученные в такого рода экспериментах величины не являются подлинными порогами чувствительности рецепторов к действию адекватного раздражения, а всегда превышают истинное значение, необходимое для возникновения импульсного ответа, на какую-то, возможно, и существенную величину.
В механизмах первичного преобразования адекватного стимула, как это будет ясно из дальнейшего изложения (см. подробнее главу 11), наиболее существенным параметром воздействия па механорецептор является амплитуда смещения. Осуществить непосредственное измерение величины этого смещения на уровне собственно рецептирующих элементов весьма трудно, особенно если принять во внимание, что обычно механический стимул должен распространиться через вспомогательные структуры рецептора, а часто и через окружающую его ткань. Кроме того, следует учесть, что расчетная чувствительность ряда механорецепторов чрезвычайно высока. Она достигает уровня, достаточного для ответа на смещение, соизмеримое с размерами молекул и даже атомов. Поэтому технически в ряде случаев задача прямого измерения пороговых смещений пока еще едва ли выполнима, хотя для ее реализации начинают применять новейшие способы измерений, такие как методы, основанные на мессбауэровском эффекте (Johnstone, Boyle, 1967; Johnstone et al., 1970; Johnstone, Taylor, 1970; Rhode, 1971, 1973), методы когерентной оптики (Tonndorf, Khanna, 1968; Khanna, Tonndorf, 1971; Michelsen, 1973; Drag- sten et al., 1974).
Наиболее чувствительными механорецепторами являются сенсорные элементы в органах чувств и прежде всего в органе слуха как у позвоночных, так и у беспозвоночных животных. Механорецепторы в тканях в целом менее чувствительны, хотя среди них имеются весьма низкопороговые рецепторные образования (например, тельца Пачини у млекопитающих или проприоцепторные хордотональные органы у насекомых и ракообразных).
Связь особенностей импульсной активности с характеристиками механического стимула. Рецептивное поле
В зависимости от особенностей РП и регенеративной системы генерации возникает характерный рисунок разряда импульсов рецепторного прибора. Этот рисунок (узор, pattern) разряда одиночного рецептора, по-видимому, может иметь большое значение для кодирования некоторых признаков раздражающего воздействия (например, скорости нарастания стимула). Вместе с тем, несомненно, что значительная часть признаков раздражения может оцениваться организмом на основе деятельности популяций рецепторов.
Как было видно из описания структуры механорецепторов (глава 1), одиночное нервное волокно иногда образует на своем конце одиночный рецептор (например, тельце Пачини), чаще же, разветвляясь, оказывается связанным с многими рецепторами, распределенными иногда на весьма большой площади (например, в коже). Зона иннервации данного афферентного волокна является его рецептивным полем. Понятие рецептивное поле имеет большое значение в физиологии сенсорных систем.
С морфологической точки зрения, рецептивное поле — это тот участок рецепторной поверхности (например, кожи, сетчатки глаза и т. д.), с которым данная нервная структура (волокно, нейрон 1-го, 2-го. . .п-го порядка) связана анатомически, т. е. жестко. При функциональном же подходе, рецептивное поле — понятие динамическое. Это означает, что в различные отрезки времени в зависимости, скажем, от характеристик падающего раздражения, один и тот же нейрон может воспринимать импульсы от различного числа рецепторов. Максимальная величина рецептивного поля нейрона в этом случае определяется его анатомическими связями, а минимальная может ограничиваться всего лишь одним рецептором. Изменение величины рецептивного поля, играющее большую роль в деятельности сенсорных систем, хорошо видно на примере сетчатки глаза, где по ходу текущей деятельности одна и та же ганглиозная клетка может суммировать работу изменяющегося числа рецепторов (Глезер, 1966; Levick, 1972). К формированию величины функционального рецептивного поля могут быть причастны явления латерального торможения, а также эфферентные влияния. Среди факторов, обеспечивающих надежность в работе периферических сенсорных структур, важную роль играет явление перекрытия рецептивных полей. Так, например, в коже зоны иннервации двух и даже более соседних афферентных волокон обычно в значительной мере перекрывают друг друга. Перекрытие рецептивных полей имеет существенное значение среди механизмов оценки центральным нервным аппаратом характеристик раздражающего воздействия (например, в коже, — J. А. В. Gray, 1962).
Как уже подчеркивалось, оценка характеристик раздражения, падающего на рецепторы из внешней или внутренней среды организма, может быть осуществлена нервной системой только на основе той последовательности импульсов (импульсного кода), которая отмечается в афферентных проводниках. Поэтому вопрос о связи различных параметров раздражающего воздействия с параметрами импульсной активности, генерируемой рецепторами, является одним из наиболее важных при изучении деятельности сенсорных аппаратов.
Начиная с самых первых электрофизиологических работ, эта сторона деятельности рецепторных приборов исследуется весьма интенсивно. Среди изученных к настоящему времени рецепторных аппаратов наиболее разнообразные способы периферического кодирования обнаружены в электрорецепторах (обзор: Акоев и др., 1971).
Наиболее распространенным для различных рецепторов способом кодирования интенсивности адекватного стимула является изменение частоты импульсации (Adrian, 1928; Granit, 1955). Этот способ кодирования является, по-видимому, основным для механорецепторов, причем существует большое число разновидностей указанного частотного кодирования. Это определяется разнообразием функциональных особенностей рецепторов. Одни из них, например, обладают фоновой активностью, а другие нет. Первые могут оценивать не только возбуждающий раздражитель (увеличение частоты импульсации), но и тормозное воздействие (уменьшение частоты разрядов). Вторые же способны оценивать только возбуждающие стимулы. В зависимости от того, является ли рецептор быстро или медленно адаптирующейся структурой, реагирует ли он на включение или выключение раздражения ит. д., рисунок ответной реакции может быть в разных сенсорных приборах различным.
Другим довольно распространенным способом кодирования в рецепторах является числовое кодирование. В этом случае сила стимула оценивается числом возникающих в рецепторе импульсов. Чем больше максимальное число генерируемых разрядов, тем более тонко рецептор может оценивать внешнее воздействие. Наряду с числом импульсов (или длительностью вспышки), по-видимому, может иметь значение и кодирование с помощью временной микроструктуры импульсной последовательности (интер- спайковые вариации). Отмеченные способы кодирования относятся к деятельности одиночных рецепторных структур. Однако существуют рецепторы, которые индивидуально, по-видимому, не могут информировать о параметрах воздействия, хотя и сообщают о самом факте воздействия. Так, например, некоторые быстро адаптирующиеся механорецепторы (типа телец Пачини) могут генерировать всего лишь один или несколько импульсов в ответ на раздражение.
Информация об интенсивности стимула в этом случае может осуществляться кодированием числом одновременно активированных рецепторов. При нарастании силы воздействия увеличивается как число возбужденных рецепторов в точке приложения стимула, так и площадь раздраженной поверхности. Изменение числа активированных рецепторов может, по-видимому, происходить очень плавно, так как оно определяется не только свойствами кожи, вызывающими постепенное затухание механической волны по мере ее распространения от точки приложения стимула (см.: Hensel, 1966), но и существованием разнопороговых нервных окончаний. Если еще учесть наличие взаимоперекрывающихся рецептивных полей, различно возбуждающихся при разных воздействиях, то легко представить, как даже с помощью одних лишь быстро адаптирующихся рецепторов животное может тонко оценивать как силу раздражения, так и его развитие во времени и пространстве. Следует, конечно, помнить, что в естественных условиях раздражение обычно падает на поверхность, содержащую большое количество различных рецепторов, каждый из которых в состоянии охарактеризовать отнюдь не все, а лишь некоторые стороны воздействия. Функция же интеграции принадлежит центральной нервной системе.
Афферентные волокна,
образующие механорецепторные окончания
Известно, что между характеристиками афферентных нервных волокон и типом рецепторных приборов, образуемых ими на периферии, существует определенная корреляция. Так, первичные нервные окончания мышечных веретен образуются наиболее толстыми афферентными проводниками, свободные же нервные
окончания образуются прежде всего немиелинизированными волокнами. Связь между типом рецептора и диаметром волокна описана и для суставов (Novotny, 1973). Следует учитывать и тот факт, что во многих случаях исследователи могут регистрировать только импульсную активность рецепторов. По характеристикам этой активности судят об особенностях самих рецепторов. Роль же регенеративной системы (а следовательно, и нервных волокон) в формировании узора ответа сенсорного прибора уже отмечалась. Нужно еще отметить, что, по-видимому, в ряде случаев имеется определенная корреляция между адаптационными способностями рецепторного прибора и аккомодационными характеристиками его нервного волокна (например, у телец Пачини).
Все это объясняет, почему при изучении импульсной активности, генерируемой рецепторами, исследователи очень большое внимание уделяют свойствам афферентных нервных проводников (обзоры: Paintai, 1963а, 1972; Дуринян, 1965; Matthews, 1972; Burgess, Perl, 1973, и др.). Характеристики этих волокон (их диаметры, скорости проведения, миелинизация) часто используются и при классификациях рецепторов, особенно в коже и внутренних органах. Иногда, зная только скорость проведения импульса по нервному волокну, исследователи пытаются сделать заключение и о диаметре волокон (и наоборот), пользуясь различными формулами, связывающими эти параметры. Обычно при работе с соматическими миелинизированными проводниками исследователи (Gasser, Grundfest, 1939; Hursh, 1939) пользуются простым эмпирическим соотношением |V|=6|Z> | (скорости выражаются в м/сек., а диаметр — в мкм), которое является достаточно приемлемым для более толстых волокон. Для тонких волокон может быть использовано соотношение |V|=4-4-5 \D | (обзоры: Boyd, Davey, 1968; Matthews, 1972). Кроме этих, используют и иные соотношения (см. обзоры: Grundfest, 1940; Аршавский и др., 1969). Для немиелинизированных волокон скорость, по-видимому, пропорциональна \jD (см. обзор: Douglas, Ritchie, 1962), что подтверждается и опытами на нервных волокнах беспозвоночных (Pumphrey, Young, 1938; Burrows et al., 1965). Однако здесь приводятся и другие коэффициенты, например |7| = 1.7 |р | (Gasser, 1955).
Использование тех или иных формул, связывающих скорости проведения ПД и диаметры волокон, затрудняется еще тем обстоятельством, что скорости проведения в тонких миелинизированных и толстых немиелинизированных волокнах могут совпадать (Шапошников, 1974). Частичное перекрытие диаметров этих групп волокон (соответственно до 1 мкм в первом случае и до 3 мкм во втором) отмечают и при морфологическом исследовании (Gasser, Grundfest, 1939; Hursh, 1939; Gasser, 1955; Гетлинг, 1969; Козлова и др., 1969, и др.). Очевидно, что нужно соблюдать осторожность при оценке размеров нервных волокон (прежде всего
g Физиология механорецепторов ЦЗ тонких) на основании электрофизиологических данных. Несомненно, что вопрос о составе волокон в различных нервах и свойствах генерируемых ими ответов требует дальнейшего исследования.
ИМПУЛЬСНАЯ АКТИВНОСТЬ ТКАНЕВЫХ МЕХАНОРЕЦЕПТОРОВ ПОЗВОНОЧНЫХ
Импульсная активность рецепторных образований кожи
Изучению кожной чувствительности уделялось всегда большое внимание. Только за последние 10—15 лет этой проблеме были посвящены многие обобщающие публикации. Эта проблема представлена в обзорах (Rose, Mountcastle, 1959; Sweet, 1959; Zotterman, 1959; Weddell, Miller, 1962; Iggo, 1965, 1966a, 1968, 1974; Hensel, 1966, 1973; Sinclair, 1967; Catton, 1970; Есаков, Дмитриева, 1971; Munger, 1971; Ильинский, 19726; Andres, During, 1973; Burgess, Perl, 1973), а также в сборнике: Advances in biology of skin, vol. 1, Cutaneous innervation, Oxford, 1960, и в трудах ряда международных симпозиумов (см. сборники: Pain and itch. Nervous mechanisms. London, 1959; Touch, heat and pain. London, 1966; Skin senses. Springfield, Ill., 1968; Тканевая рецепция. Л., 1974).
Несмотря на успехи, достигнутые в изучении кожной чувствительности, особенно благодаря широкому применению морфофизиологических подходов, использованию электронномикроскопических и тонких электрофизиологических методов, проведению эспериментов не только на животных, но и на людях, многие стороны данной проблемы остаются еще открытыми. Прежде всего до сих пор еще остается дискуссионным старый вопрос о соотношении структуры рецепторов и их функции (см. о теориях кожной чувствительности: Sinclair, 1967; Ильинский, 19726). Оставляя сейчас в стороне вопрос о сильных и слабых сторонах той или иной концепции, укажем, что теория специфичности кожной чувствительности в ее современной форме, говорящая о приспособленности отдельных видов кожных рецепторов к выполнению специфической функции, результатом которой является проявление специфического ощущения, обладает в настоящее время весьма вескими аргументами в свою пользу. И можно, например, с уверенностью говорить о механорецепторной функции ряда кожных сенсорных приборов, таких как тельца Пачини, Мейсснера, тактильные корпускулы. К механорецепторам, по-видимому, следует отнести также тельца Руффини (Chambers et al., 1972), длительное время рассматривавшиеся как тепловые рецепторы, и, конечно, ряд нервных окончаний вокруг волос. Некоторые авторы приписывают механорецепторную функцию и таким рецепторам, как колбы Краузе (Winkelmann, 1960с), которые раньше обычно
относились к холодовым? рецепторам [современная тонна арония по этому вопросу изложена Минут-Сорохтиной (1972)].
Благодаря свойствам перечисленных, а также ряда других еще не столь точно идентифицированных механорецепторов формируются ощущения прикосновения, давления, смещения кожной поверхности и волосяного покрова. Сигнализация от механорецепторов участвует и в формировании более сложных ощущений (Sinclair, 1967). Некоторые авторы (Rose, Mountcastle, 1959) говорят о тактильных стимулах как о воздействиях, к которым можно отнести все виды раздражений, вызывающих смещение волос или деформацию кожи без ее повреждения. Они же выделяют и кинестетические стимулы, к числу которых причисляют механические воздействия, достигающие через неповрежденную кожу рецепторных образований подкожной соединительной ткани, надкостницы, костей, оболочек сухожилий, мышечных фасций, суставных сумок. Часто в качестве самостоятельного вида выделяют вибрационное ощущение, возникающее при действии на кожную поверхность ритмических колебаний (Hensel, 1966).
Вопрос о том, какие точно структуры кожи связаны с восприятием тех или иных видов механических воздействий, остается еще недостаточно выясненным. Полагают, что прикосновение и давление воспринимаются нервными сплетениями, окружающими волосяные фолликулы, тельцами Мейсснера и Пачини, тактильными корпускулами, свободными нервными окончаниями. Вибрация воспринимается фактически всей кожной поверхностью, хотя чувствительность разных ее участков существенно разнится (стр. 123). Вместе с тем отмечается, что у губ, обладающих высокой чувствительностью к тактильным раздражениям, нет специальной чувствительности к вибрационным воздействиям (Caine, Pallis, 1966). Морфологическим субстратом, обеспечивающим вибрационную чувствительность, являются преимущественно механорецепторы кожи, прежде всего фазного типа, локализованные как в поверхностных, так и в глубоких ее слоях. Рецепторы, расположенные в более глубоких слоях кожи, в большей степени участвуют в восприятии вибраций. Вибрационную чувствительность могут обеспечивать также механорецепторы, заложенные в фасциях, надкостнице, тканях суставов. Мышечные же веретена, по-видимому, не участвуют в обеспечении вибрационной чувствительности (см. обзоры: Keidel, 1956; Bekesy, 1959b; Hensel, 1966; Caine, Pallis, 1966).
Основные исследования свойств кожных рецепторов проводятся по сей день в экспериментах с регистрацией импульсной активности одиночных нервных волокон. Все афферентные нервные волокна кожных нервов млекопитающих (табл. 1) можно подразделить на три основные категории (Gasser, 1955, 1960): 1) миелинизированные альфа-волокна (группа Аа; диаметр 6—17 мкм, включая миелин; скорость проведения 30—100 м/сек.); 2) миелинизированные дельта-волокна (группа AS; диаметр 1—5 мкм,
А
«
сб
3
сб
3
3
3
ч
о
я=я
Ф о ЯГ д
я
я
3
Л
I
Он с
3
Он
3
о ф
ч
3
и
и
со сб е
я
я
сб
.
я
■3
§
я
ф . 3 Сб я
Ч
«
aS
o' Я § »о g я
й
«
В*
2
S ,й
Я
I
3
12 R,
vo g 3 ф
я
й^з 2-2 я tf g я Ф Я Ч
о
Я
о
о
з
5
А
2
Й
Я g Я ф
ф
Я’оя Я" ф о Ф Они Ом д |
ф ф с?
з я “ я ч
Л
3
® ф я
Я"
в tд
Ф яг
я
§ i'g
Й
ф Э о я
я
я
Таблица
1 Кожные афферентные волокна и типы
рецепторов (по: Burgess, Perl, 1973)
3
3 ЙО О о
к
Я
& я
3
.
3"
з
о
Я
Он
Ф
Примечание.
1. Самая ранняя классификация волокон
(Gasser, Erlanger, 1927; Erlanger, Gasser, 1937) включала
бета-и гамма- волокна в дополнение к
Аа-, АЗ- и С-группам, показанным в таблице,
и относилась как к моторным, так и к
сенсорным волокнам. Далее была предложена
классификация специально для сенсорных
волокон в мышечных нервах, разделяющая
волокна на группы I, II, III и IV (Lloyd, 1943).
Позже Гассер (Gasser, 1960) вновь проверил
классификацию кожных нервов; гамма-волна
сложного потенциала n. saphenous кошки
была признана в основном артефактом,
а бета-волна была недостаточно устойчивой.
включая миелин; скорость проведения 4—30 м/сек.); 3) немиели- низированные волокна (группа С; диаметр 0.3—1.6 мкм; скорость проведения 0.4—2 м/сек.). По сравнению с афферентными проприоцептивными проводниками (которые подразделяются на группы I—IV, Lloyd, 1943) в кожных нервах отсутствуют очень толстые миелинизированные волокна (сравн. с табл. 3). Волокна всех групп кожных нервов могут быть связаны с механорецепторами, однако основная часть механорецепторных приборов связана с окончаниями волокон группы Аа.
По своим функциональным характеристикам механорецепторы в коже могут быть подразделены на фазные и тонические. Различные механорецепторы по-разному могут относиться к таким параметрам стимула, как скорость и направление воздействия. У них выявлена также и разная скорость восстановления характеристик после действия раздражения. Среди кожных механорецепторов есть структуры, отвечающие исключительно на механические стимулы (например, механорецепторы волос и тельца Пачини), и неспецифические структуры, реагирующие как на механические, так и на термические воздействия (нервные окончания мякотных волокон Аа и AS групп и немиелинизированных волокон С группы). Все эти свойства учитываются при классификации кожных механорецепторов, однако в настоящее время в литературе нет установившейся единой точки зрения. Отсутствие единой классификации, естественно, затрудняет единообразное изложение данных, полученных в этой области.
До настоящего времени методические причины, как и в случае многих других сенсорных приборов, не позволили исследователям провести изучение локальных электрических процессов, протекающих в рецепторах кожи. Единственным исключением являются тельца Пачини, которые в большом количестве располагаются в подкожных структурах и которые широко вовлекаются в процесс возбуждения при различного рода механических воздействиях на кожу. Нет никаких оснований полагать, что эти рецепторы в чем-либо существенно отличаются от аналогичных телец Пачини в брыжейке кишечника кошки, свойства которых к настоящему времени подверглись детальному изучению.
Активность телец Пачини
(окончания волокон Аа группы)
Общая характеристика импульсного ответа. Волокна этих механорецепторов относятся в основном к группе Аа (Sheehan, 1933; Gernandt, Zotterman, 1946; Quilliam, Sato, 1955; McLeod, 1958; Волкова, Черепнов, 1971, и др.) и проводят импульсы со скоростью от 13 до 70 м/сек. (Downman, Evans, 1957; Ильинский, 1963; Leitner, Perl, 1964, и др.). При этом рецепторы больших размеров связаны с более толстыми и быстропроводящими волокнами.
Продолжительность измененной возбудимости регенеративной системы телец Пачини невелика: обычно она не превышает 5— 6 мсек. (J. А. В. Gray, Malcolm, 1950; Loewenstein, Altamirano- Orrego, 1958; Armett, Hunsperger, 1961; Ильинский, 1962; Ozeki, Sato, 1964). Поэтому при достаточно сильных стимулах межспай- ковый интервал может быть весьма кратким.
На примере телец Пачини хорошо видно, как свойства РП отражаются в характеристиках импульсного ответа, хотя и не определяют его полностью. Крайне небольшая продолжительность


Рис. 37. Импульсная активность телец Пачини (по: Ильинский, 1966в).
А — характер ответов телец Пачини в зависимости от характера раздражения. 1—3 — зависимость возникновения ПД от изменения скорости раздражения; 4, 5 — on- и off-ответы телец Пачини; 6 — множественные разряды рецепторов (записи с наложением). Отметка времени: на 1—3 — к, на 4—6 — 5 мсек.; калибровка: на 1—3 — 200, на 4—6 — 50* мкв. В — схема колебательных процессов, происходящих в капсуле рецептора при деформации, а, д — рецептор в состоянии покоя; б, в — нанесение и снятие раздражения (стрелки); г —- период свободного колебания слоев капсулы рецептора.
локального ответа в тельцах Пачини приводит к тому, что один деполяризационный РП вызывает появление только одного (рис. 37), реже — двух ПД (J. А. В. Gray, Sato, 1953; Loewenstein, Altamirano-Orrego, 1958; Ильинский, 1962). Критический уровень деполяризации, при котором возникает ПД, составляет примерно 10—16% от его амплитуды (J. А. В. Gray, Sato, 1953; Ильинский, 1962). Указанное значение критической деполяризации относится к пороговым амплитудам воздействия. При увеличении силы раздражения, когда скорость нарастания РП возрастает, величина критического уровня несколько снижается^ На возникновении ПД сказывается скорость нарастания стимула (рис. 37, 1—3), так как величина РП, как об этом говорилось ранее (стр. 79), существенно зависит от этого параметра раздражения. При достаточной длительности раздражающего стимула наблюдается появление он- и off-ответов (рис. 37, 4—5}. При сильных механических ударах рецептор может генерировать множественные разряды, возникающие из ряда последовательных РП.
При ритмической стимуляции телец Пачини возникает ритмический разряд импульсов (рис. 38). Однако при достаточно большой частоте воздействия отмечается явление трансформации
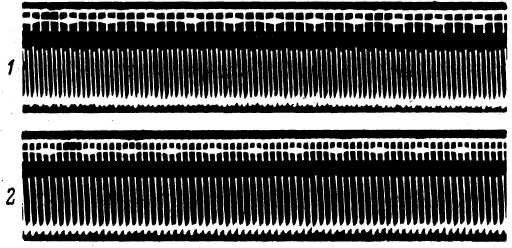


Рис. 38. Действие ритмического раздражения на тельце Пачини (Ильинский, 1966а).
Активность рецептора на первой секунде после включения ритмического механического раздражения с частотой 200 (1), 300 (2), 400 (3) и 650 гц (4). Сверху вниз: отметка раздражения; отметка времени (20 мсек.); биоэлектрическая активность.
ритма: количество ПД становится меньше числа стимулов (рис. 38, 4). На протекании указанного явления сказывались сила и длительность раздражения. Когда же в ходе высокочастотного раздражения рецептора наблюдается суммация отдельных РП в единый стационарный деполяризационный ответ, импульсная активность блокируется полностью благодаря быстрой аккомодации регенеративной системы генерации. Таким образом, при определенных условиях раздражения между локальной и импульсной активное!ью рецептора не наблюдается соответствия.
При пороговых интенсивностях раздражения ПД возникают нерегулярно. При этом их амплитуда и латентный период весьма варьируют. Флуктуация амплитуды ,ПД отмечается только вблизи рецептора. На некотором удалении от него величина спайков стационарна (Ильинский, 1963, 1966г). Латентный период возникновения ПД закономерно снижается по мере увеличения силы раздражения. При больших силах раздражения его величина приближается к величине скрытого периода РП. Между силой раздражения и латентным периодом ПД имеется экспоненциальная зависимость (J. А. В. Gray, Malcolm, 1950; Ильинский, Фикс, 1963).
Место возникновения ПД в тельцах Пачини. По мнению одних исследователей (Diamond et al., 1956; Loewenstein, Rathkamp, 1958), местом возникновения ПД является область первого перехвата Ранвье. Другие авторы считают, что регенеративные структуры локализованы и на мембране немиелинизированной части окончания (Hunt, Takeuchi, 1962; Sato, Ozeki, 1963; Ozeki, Sato, 1964), т. e., с их точки зрения, участки мембраны, обладающие нерегенеративными и регенеративными свойствами, располагаются мозаично. Хотя доводы последних авторов и кажутся достаточно убедительными, тем не менее не все исследователи с ними согласны (Loewenstein, 1971). Вне зависимости от окончательного решения этого вопроса, никак нельзя исключить возможность первичного возникновения распространяющегося потенциала действия не в безмякотной части окончания, а в другом месте, в области своего рода «наибольшего благоприятствования», как это имеет место в мотонейронах (область аксонального холмика — Eccles, 1957) или в рецепторах растяжения (начальная часть аксона — Edwards, Ottoson, 1958). Такой областью может быть не только район первого перехвата Ранвье, как предполагалось ранее (J. А. В. Gray, Sato, 1953), но, по-видимому, и самое начало немиелинизированной части нервного волокна, где имеется сужение осевого цилиндра и где плотность генераторного тока на единицу поверхностной мембраны должна быть наибольшей (Ильинский, Волкова и др., 1974).
Ответы на включение и выключение раздражения. Множественные разряды. Известно, что многие сенсорные приборы способны отвечать возбуждением при выключении раздражения (Granit, 1955). Важную роль в возникновении этих ответов играют свойства регенеративной системы генерации, которые могут быть сходными у рецепторов, воспринимающих самые разные раздражения. Однако у механорецепторов в формировании off-ответов большое значение могут иметь также и вспомогательные структуры. Особенно четко это удалось выяснить на примере телец Пачини (Ильинский, 1965, 1966b, 1971; Loewenstein, Mendelson, 1965; Loewenstein, 1966, 1971; Loewenstein, Skalak, 1966; Nishi, Sato, 1968). Анализ происхождения off-ответов в тельцах Пачини выявил ряд фактов, очень существенных для понимания многих аспектов деятельности данных рецепторов. Эти аспекты будут рассмотрены в дальнейшем.
Первоначальные наблюдения над on- и off-ответами дали противоречивые результаты. Согласно одним авторам (J. А. В. Gray, Malcolm, 1950; J. А. В. Gray, Sato, 1953), off-ответ в тельцах Пачини наблюдался не всегда и обычно имел более высокий порог возникновения, чем оп-ответ. По другим данным (Ильинский, 1963), off-ответ наблюдался не реже on-реакции и порог его возникновения часто был ниже, чем для оп-ответа. Дальнейшее изучение показало (Ильинский, 1966в), что on- и off-ответы могут наблюдаться практически во всех рецепторах, хотя пороги их возникновения и латентное время могут существенно варьировать. Более того, оказалось, что на одном и том же рецепторе можно получить различное соотношение между on- и off-ответами, меняя направление и место нанесения толчка, а также величину исходного давления на капсулу тельца. Это заставило предположить (Ильинский, 1965, 1966в), что в возникновении off-ответа большую роль играют механические свойства капсулы тельца, которая обладает хорошей упругостью. Согласно этой гипотезе, в момент включения раздражения происходит деформация нервного окончания в одном направлении (в направлении действия стимула), а в момент выключения за счет сил упругости капсулы возникает деформация в направлении, перпендикулярном первому. Таким образом, тельца, а соответственно и нервные окончания, деформируются дважды, но в разном направлении. Результатом первой деформации нервного окончания является возникновение on-, а второй — off-реакции (схема на рис. 37).
Эксперименты на декапсулированных тельцах показали (Ильинский, 1966в; Loewenstein, Skalak, 1966; Nishi, Sato, 1968), что поведение сильно декапсулированных телец действительно существенно отличалось от реакции интактных рецепторов. Как правило, off-ответ у них отсутствовал, и при этом пороги для оп-ответа понижались. Длительность РП, возникавшего при раздражении декапсулированных рецепторов стимулами большой продолжительности, была больше, чем у интактных телец (см. в главе 4, рис. 65). Специальное рассмотрение этого вопроса показало, что свойства капсулы телец Пачини являются основным фактором быстрой адаптации этих образований (Ильинский, 1965; Loewenstein, Mendelson, 1965), а off-ответы являются реакциями на саму деформацию, а не на ее прекращение.
Если off-реакция по существу не отличается от оп-реакции, то изменения ее под влиянием любого воздействия должно идти параллельно с изменениями оп-ответа. Опыты показали, что под влиянием температурных воздействий on- и off-деполяризационные РП изменяются сходным образом. При охлаждении пороги реакции повышались, а амплитуда ответов и скорость нарастания уменьшались. Все изменения шли параллельно для обоих видов ответов (Ильинский, 1966в).
На основании полученных результатов принято считать, что ответ телец Пачини на выключение раздражения фактически является разновидностью ответа на включение воздействия и зависит от характеристик раздражения и свойств капсулы рецептора. Возникновение его происходит при снятии раздражения благодаря существованию обратно направленного смещения в капсуле рецептора. В дальнейшем мы вернемся еще раз к off-ответам

Рис. 39. Изменения ответов на краткий тестирующий стимул пороговой силы в зависимости от времени его приложения относительно длительного обусловливающего толчка^(по: Ильинский, 1966а).
1 — тестирующий стимул не подан; 2 — тестирующий стимул подан в период рефрактер- ности после off-ответа; 3—8 — тестирующий стимул подан на фоне постоянной деформации рецептора'; 9 — тестирующий стимул подан непосредственно перед началом регистрации (не попал в кадр). Сверху вниз: отметка раздражения; биоэлектрическая активность (запись с наложением). Отметка времени — 4 мсек.; калибровка — 200 мкв.'
телец Пачини при рассмотрении гиперполяризационных ответов этих рецепторов (стр. 440).
Единство происхождения on- и off-ответов телец Пачини делает вероятным предположение об их тождественности и в некоторых других чувствительных образованиях. Скорее всего такого рода положение может иметь место в различных механо- рецецторных структурах, обладающих довольно сложным вспомогательным аппаратом, и в частности в тех пластинчатых рецепторах, которые имеют строение, сходное с тельцами Пачини (см., напр., Quilliam, 1966). Капсула телец Пачини участвует и в возникновении множественных ответов при больших амплитудах
 воздействия
(Ильинский, 1966в). При раздражении
рецепторов с интактной капсулой стимулами
даже большой амплитуды, но с плавно
изменяющимися фронтами нарастания и
спадения (в этом случае возникновение
колебательного процесса в капсуле
затруднено) наблюдается обычно появление
лишь одного ПД. В декапсулированных
рецепторах также отмечалось появление
только одного ПД в момент включения
стимула. Напротив, в случае интактных
рецепторов даже тогда, когда возникает
лишь один on- и один off-ПД, можно на фоне
действия продолжительного толчка с
крутыми фронтами отметить волнообразное
изменение возбудимости рецептора,
обусловленное колебательными процессами
в капсуле (рис. 39). Возникновению
множественных ответов в тельцах Пачини
способствует также и явление
постпгиперполяри-
зационного возбуждения
(Ильинский, 1965, 1966г, 1969). В этом случае
отмечается появление осциллирования
локального потенциала рецептора и
снижение критического уровня генерации
воздействия
(Ильинский, 1966в). При раздражении
рецепторов с интактной капсулой стимулами
даже большой амплитуды, но с плавно
изменяющимися фронтами нарастания и
спадения (в этом случае возникновение
колебательного процесса в капсуле
затруднено) наблюдается обычно появление
лишь одного ПД. В декапсулированных
рецепторах также отмечалось появление
только одного ПД в момент включения
стимула. Напротив, в случае интактных
рецепторов даже тогда, когда возникает
лишь один on- и один off-ПД, можно на фоне
действия продолжительного толчка с
крутыми фронтами отметить волнообразное
изменение возбудимости рецептора,
обусловленное колебательными процессами
в капсуле (рис. 39). Возникновению
множественных ответов в тельцах Пачини
способствует также и явление
постпгиперполяри-
зационного возбуждения
(Ильинский, 1965, 1966г, 1969). В этом случае
отмечается появление осциллирования
локального потенциала рецептора и
снижение критического уровня генерации
Рис. 40. Амплитудно-частотная характеристика вибрационного раздражения кожи при пороговой силе ощущения (по: Bekesy, 1939).
По оси абсцисс — частота стимуляции (гц); по оси ординат — амплитуда раздражения (мм).
ПД. Повышение возбудимости рецептора также способствует появлению множественных разрядов: например, при повышении температуры (Inman, Peruzzi, 1961; Sato, 1961; Ильинский, 1966а), в результате действия симпатомиметических средств (Loewenstein, Altamirano-Orrego, 1956; Черепнов, 1969; Акоев и др., 1974а, 19746), при сохранении нормального кровоснабжения рецепторов (J. А. В. Gray, Malcolm, 1950; J. А. В. Gray, Matthews, 1951b; Ильинский, 1966a).
Тельца Пачини кожи в силу своих свойств обусловливают обнаружение быстро протекающих воздействий. В частности, полагают, что данные механорецепторы могут рассматриваться как структуры, в значительной мере обеспечивающие вибрационную чувствительность организма в целом (Caine, Pallis, 1966).
Вибрационная чувствительность у человека. Вибрационной чувствительностью обладает кожа, как покрытая волосами, так и лишенная их. В последнем случае диапазон воспринимаемых частот шире, а пороги восприятия ниже. Наибольшей вибрационной чувствительностью у человека обладает кожа в области пальцев рук, а наименьшей — в брюшной и ягодичной областях (Wil- ska, 1954). Такое различие в порогах зависит прежде всего от механических свойств тканей, расположенных в данной области (развитие жирового слоя, мышц, костей и т. д.), и от различной плотности распределения рецепторов. Последнее следует из за
висимости порогов от площади поверхности вибрационного воздействия (Haller, 1935; Verrillo, 1966).
у
человека в ^диапазоне оптимальных
частот могут разниться.
 В
среднем, для кончика пальца при частоте
200 гц пороговое смещение равняется
10-4
мм. Эта величина при частоте 400 гц
В
среднем, для кончика пальца при частоте
200 гц пороговое смещение равняется
10-4
мм. Эта величина при частоте 400 гц
Рис. 41. Чувствительность изолированного тельца Пачини к синусоидальной вибрации различной частоты (по: Sato, 1961).
По оси абсцисс — частота стимуляции (гц); по оси ординат — абсолютные амплитудные пороги (усл. ед.). Температура среды — 24 (Z) и 37° (•?).
может равняться 5*10-5 мм (Wilska, 1954), а в других экспериментах порог был равен даже 1.3-10~5 мм (Keidel, 1952).
Данные, полученные при исследовании вибрационного ощуще^ ния, следует сопоставить с результатами экспериментов, выполненных на одиночных рецепторах. Линдблом и Лунд (Lindblom, Lund, 1966) считают, что порог тактильного ощущения определяется порогом возбуждения наиболее чувствительных рецепторов кожи — телец Пачини. Можно отметить, что количество их на пальцах и ладонях резко возрастает у слепых, тактильная чувствительность которых чрезвычайно высока и превышает таковую у обычных людей (Отелин, 1961). Исследование способности телец Пачини кошки воспринимать вибрационные стимулы показало (Sato, 1961), что наименьшими порогами данные рецепторы обладали в районе 200—300 гц (рис. 41), т. е. именно в области оптимальных частот для вибрационных ощущений. По другим данным (Миркин, 1966), максимальная чувствительность рецепторов у кошки отмечалась в диапазоне 100—120 гц, а у обезьяны — в диапазоне 125—150 гц (Lindblom, Lund, 1966). Величины пороговых смещений (10“4—10“5 мм), полученные для телец Пачини разными исследователями (J. А. В. Gray, 1959b; Loewenstein, 1965; Ильинский, 1966а), также хорошо соответствуют величине минимальных вибрационных порогов. По последним данным (Гаврилов и др., 1974), порог возбуждения одиночных изолированных телец Пачини при использовании ультразвуковых стимулов равнялся 3.0—5.0 • 10-5 мм, а порог тактильного ощущения у человека, полученный при том же способе раздражения с помощью фокусированного ультразвука, равнялся 1.0-1.2-10-4 мм.
Таким образом, имеются основания считать, что за пороги вибрационной чувствительности действительно могут отвечать именно тельца Пачини и сходные с ними образования. Возбуждение этих рецепторов при различного рода вибрациях, наносимых на кожу, была продемонстрирована многими исследователями (Hunt, McIntyre, 1960а; Skoglund, 1960; Hunt, 1961; Janig et al., 1968; Merzenich, Harrington, 1969; Nilsson, 1969b; Lynn, 1971, и др.).
Следует заметить, что исследование действия вибраций представляет специальный интерес, связанный с их широким распространением в среде, окружающей человека, а также в связи с возможным использованием такого рода раздражителя в прикладных целях. Установлено, например, что можно различать высоту звуков посредством вибраций. Правда, способность различения частот в этом случае по сравнению со слухом чрезвычайно грубая (см. обзор: Caine, Pallis, 1966).
Активность рецепторов I и II типа
(окончания волокон Аа группы)
Другие механорецепторы, связанные с кожными покровами, изучены значительно меньше. Сведения о них получены только на основании регистрации импульсной активности. На базе исследования этого рода, а также с учетом некоторых морфологических особенностей среди механорецепторов кожи можно выделить несколько типов (Iggo, 1966а, 1968; Brown, Iggo, 1967; Burgess et al., 1968; Perl, 1968). В коже, покрытой волосами, отмечают два типа медленно адаптирующихся рецепторов (тип I й тип II), которые являются окончаниями волокон А а и которые не связаны с волосяными фолликулами (Iggo, 1966а).VIII Афферентные волокна рецепторов I типа, согласно Игго, относятся к быстропроводящим волокнам: у кошек большинство из них имеет скорость проведения 48—72 м/сек.; волокна рецепторов II типа являются несколько более медленными проводниками: у кошек максимум волокон имеет скорость 36—66 м/сек. Рецепторами I типа, которые встречаются чаще, являются тактильные корпускулы (рис. 2 — touch corpuscles) (Iggo, 1963; Iggo, Muir, 1969). Рецепторами II типа, по-видимому, следует считать тельца Руффини (рис. 6 — Chambers et al., 1972). Рецепторы I типа в силу особенностей своего строения нечувствительны к тангенциальному растяжению кожи, в то время как рецепторы II типа, напротив, весьма чувствительны к такого рода воздействию. Тактильное тельце (корпускула) занимает достаточно большую поверхность кожи (несколько сот микрон), и можно показать, что наибольшей чувствительное! ью (порог — 1—5 мкм) обладает именно средняя область структуры (Iggo, 1966а; Iggo, Muir, 1969). Это, по-видимому, связано с особенностями распределения механических усилий в корпускуле. Так как одиночное нервное волокно может быть связано с несколькими (2—3) тактильными корпускулами, то участков с максимальной чувствительностью для волокон рецепторов I типа может быть несколько. Каждый же рецептор II типа связан с самостоятельным волокном, и поэтому максимум чувствительности всего лишь один. Рецепторы II типа обладают дирекционной чувствительностью: растяжение кожи в одном направлении дает максимальный эффект, а растяжение под углом в 90° к этому направлению вызывает снижение активности (Knibestol, Vallbo, 1970; Chambers et al., 1972).
В ответ на плавное, мягкое механическое воздействие (смещение кожи на 5—15 мкм) рецепторы I типа отвечают резким всплеском импульсации. Частота разряда рецепторов I типа может превышать 1000 в 1 сек. При сходных же воздействиях на рецепторы II типа мгновенная частота не превышает 800 имп./сек. (Chambers et al., 1972). Оба типа рецепторов обладают способностью генерировать активность в течение длительного (2—4 мин.) действия статического раздражителя. Однако скорость адаптации у рецепторов I типа больше и стационарная частота его разрядов меньше (10 имп./сек. по сравнению с 20—25 имп./сек.) и менее регулярна (стандартное отклонение в гистограмме распределения интервалов достигает 50% от среднего значения при частотах 15—50 имп./сек.), чем у рецепторов II типа (стандартное отклонение составляет 5—10%) (Iggo, 1966а, 1968; Burgess et al., 1968; Iggo, Muir, 1969; Chambers et al., 1972). Рецепторы I типа обладают большей чувствительностью к влиянию скорости раздражения. Таким образом, рецепторы I типа могут рассматриваться как преимущественно детекторы скорости воздействия, а рецепторы II типа — как преимущественно детекторы положения.
В состоянии покоя рецепторы I типа либо не обладают активностью, либо частота их импульсации не превышает 1—2 имп./сек., в то время как рецепторы II типа могут иметь фоновую активность с частотой порядка 20 имп./сек. После нанесения раздражения восстановление возбудимости у рецепторов II типа происходит медленнее (более чем через 100 мсек.), чем у рецепторов I типа (менее 40—50 мсек.). Рецепторы II типа обладают в целом свойствами, сходными с другими нервными окончаниями типа телец Руффини, локализованными в самых различных тканевых структурах.
Активность рецепторов Г, G, D (окончания волокон Аа и Л В групп)
Изучение механорецепторов, связанных с волосяной сумкой, было начато на заре электрофизиологических исследований (Adrian, 1931). В дальнейшем они неоднократно исследовались многими авторами (см. обзоры: Есаков, Дмитриева, 1971; Burgess, Perl, 1973). В зависимости от свойств и локализации их можно подразделить на несколько видов. Согласно Игго (Iggo, 1966а, 1968; A. G. Brown, Iggo, 1967), вокруг различных волосяных фолликулов наблюдаются три типа рецепторов: Т, G и D. Все они относятся к разряду фазных и могут отвечать только одним импульсом на небольшое смещение. При непрерывном смещении волоса они могут генерировать более длительный разряд.
Рецепторы типа Т образованы альфа-волокнами (диаметр 9—16 мкм), разветвляющимися вокруг специализированных tylotrich-фолликулов (Straile, 1960,1961). Эти фолликулы окружены большим венозным синусом, и их волос более прямой и длинный, чем обычный сторожевой. Они встречаются у различных млекопитающих в разном количестве (у кроликов чаще, чем у кошек). Каждое афферентное волокно дает разветвления вокруг 3—10 волосяных фолликулов.
Рецепторы типа G образованы альфа-волокнами (диаметр 4—16 мкм), разветвляющимися вокруг сторожевых волосков (guard hair). Каждое волокно иннервирует 10 и более фолликулов. Величина рецептивного поля не превышает 2 см2. В зависимости от скорости раздражения, которая необходима для возбуждения рецепторов этого типа, они подразделяются на рецепторы Gx (скорость механического воздействия больше 20 мм/сек.), G2 (скорость меньше 0.5 мм/сек.) и рецепторы промежуточного характера. Рецепторы Gx связаны с наиболее быстро проводящими волокнами, хорошо реагируют на вибрационные воздействия высокой частоты (выше 60—80 гц) и большой амплитуды (свыше 50 мкм). Обладая высокими адаптационными свойствами, они редко генерируют больше, чем 2—3 импульса, быстро восстанавливают свою возбудимость (в пределах 10 мсек.)5 имеют низкую дирекционную чувствительность. Наряду с тельцами Пачини они сигнализируют о динамических параметрах механического раздражения кожи. Рецепторы G2 в отличие от рецепторов двух других типов образуются наиболее медленно проводящими волокнами; цикл восстановления возбудимости у них протекает также наиболее замедленно. Они обладают тоническим компонентом ответа, который угасает довольно быстро (в пределах 1—2 сек. после достижения нового положения волоса).
Наконец, рецепторы типа D образуются вокруг простых (down hairs) и иногда сторожевых волосков дельта-волокнами (диаметр 2—4 мкм). Рецептивное поле таких волокон может достигать 2 см2. Рецепторы типа D характеризуются очень высокой
чувствительностью к перемещению волос. При этом их реакция на медленные смещения больше, чем у рецейторов типа G2. Частота их разряда возрастает по мере увеличения скорости нанесения раздражения.
Активность рецепторов вибрисс
Многие млекопитающие обладают специальными чувствительными волосками — вибриссами. Вибриссы имеют весьма сложную систему нервных окончаний, которые различаются по своим свойствам (Fitzgerald, 1940; Kerr, Lysak, 1964; Nilsson, Skoglund, 1965; Iggo, 1968; Nilsson, 1969a, 1969b; Zucker, Welker, 1969;. Gottschaldt et al., 1973).
Прежде всего имеются рецепторы, демонстрирующие чувствительность к статическим смещениям, что отличает их от нервных окончаний вокруг обычных волос. Они также чувствительны к скорости нанесения раздражения и к его направлению. Дирек- ционная чувствительность оказалась более выраженной у рецепторов с менее регулярной активностью. Предполагают, что данным типом рецепторов являются тельца Меркеля, располагающиеся среди нервных окончаний вибриссы (Gottschaldt et al., 1973). Напротив, у рецепторов с более ритмичной активностью и наличием фоновой импульсации (порядка 20 имп./сек.) чувствительность к направлению раздражения оказалась ниже.
Среди нервных окончаний вибриссы имеются рецепторы, весьма сильно реагирующие на динамическую составляющую воздействия. Они обладали незначительной дирекционной чувствительностью (Nilsson, 1969а). Наконец, были обнаружены фазные рецепторы с хорошо выраженной реакцией на вибрационное воздействие и отсутствием дирекционной чувствительности (Nilsson, 1969b; Zucker, Welker, 1969; Gottschaldt et al., 1973). Предполагают, что ими могут являться скопления телец Пачини или сходных с ними образований, встречающихся вблизи вибрисс.
Активность F-рецепторов
(окончания волокон Аа группы)
В коже, покрытой волосами, имеются еще рецепторы фазного типа, образованные быстропроводящими альфа-волокнами (тип F). Они не возбуждаются (в отличие от рецепторов волосяных сумок) при движении одиночных волосков, но могут генерировать разряд при одновременном смещении группы волос, особенно если оно осуществляется воздействием, приложенным вблизи кожной поверхности. Это говорит о том, что данные рецепторы возбуждаются не движениями волос как таковыми, а теми смещениями кожи, которые возникают при одновременном движении большой группы волос. Действительно, прямое раздражение кожи в зоне поля данного афферентного волокна дает значительную ответную реакцию. Рецепторы этого типа получили название рецепторов области, или участка (field receptors — Burgess et al., 1968; Perl, 1968; Burgess, Perl, 1973). У кошек они наиболее многочисленны в волосистой коже вокруг подушечек ног и пальцев. Часть F-рецепторов обладает незначительной позиционной чувствительностью (F-l), другие же в большей степени могут сигнализировать и о новом постоянном состоянии (F2). Кроме того, рецепторы F2 обладают большей чувствительностью к тому, удаляет ли раздражение сенсорную структуру от положения покоя или же, напротив, приближает к нему, т. е. обладают линейной дирекционностью. Позиционная реакция рецепторов F2 выражена больше, чем у рецепторов G2, и они могут обладать низкочастотной активностью даже через 10—15 сек. после нанесения резкого раздражения.
F-рецепторы также весьма обычны и в коже, лишенной волос, например на подошвах и пальцах лап у кошки, ладонях обезьян. (Janig et al., 1968; Lynn, 1969). Имеются основания считать тельца Мейсснера и подобные им образования ответственными за возникновение ответов F-гипа (Janig, 1971).
Активность С-механорецепторов (окончания волокон группы С)
Механорецепторы кожи, являющиеся окончаниями немиелинизированных волокон, были впервые описаны Соттер- маном (Zotterman, 1939) и в дальнейшем изучались рядом авторов (Douglas, Ritchie, 1957; Douglas et al., 1960; Iggo, 1960; Iggo, Kornhuber, 1968; Bessou et al., 1971). Одним из примеров С-механорецепторов являются рецепторы волосяного мешочка. С-во- локна, заканчивающиеся механорецепторами, составляют 50% немиелинизированных волокон в проксимальных отделах афферентных нервов задней конечности кошки. При этом у кошек и обезьян эти рецепторы не обнаружены в коже, лишенной волос (Bessou et al., 1971).
С-механорецепторы обладают рядом особенностей, отличающих их от рецепторов, связанных с миелиновыми волокнами. Прежде всего, пороги их возбуждения выше, а минимальное время, необходимое для их активации, значительно больше, чем у механорецепторов мякотных волокон (150—200 мсек, при 6-пороговом воздействии в первом случае и не более, чем 3 мсек., во втором). Это время может значительно возрасти (до 500 мсек.), если на предыдущей минуте рецептор уже возбуждался (Bessou et al., 1971). Поэтому при синусоидальных воздействиях с частотой выше 1—2 гц такие рецепторы редко возбуждаются. Медленное вовлечение в процесс возбуждения и наличие очень длительных следовых явлений весьма характерны для С-механорецепторов.
При различных стационарных воздействиях в С-механорецеп- торах обнаруживаются явления адаптации. Первоначальный пик активности в течение 3—4 сек. быстро уменьшается, и отдельные импульсы сохраняются в протяжении последующих 15—20 сек. Адаптация С-механорецепторов протекает так же, как аналогичный процесс у Р2-рецепторов.
С-механорецепторы волос не отвечают на движение изолированных волос, но могут генерировать умеренный разряд при одновременном движении нескольких волос вблизи их основания (Iggo, 1960).
При резком снятии стационарного раздражения С-механорецепторы часто генерируют разряд, что существенно отличает их от рецепторов А-волокон (Zotterman, 1939; Douglas, Ritchie, 1957; Iggo, 1960). Наиболее выражен такой следовой разряд (after discharge) в С-волокнах при движении раздражителя с достаточно медленной скоростью через рецептивное поле. Некоторые механорецепторы С-волокон могут возбуждаться при резких перепадах температуры (при значительном охлаждении до 5—15° у кошек и при нагревании до 55—70° у приматов).
В целом С-механорецепторы наиболее эффективны в обнаружении медленно протекающих механических раздражений.
Импульсная активность рецепторов внутренних органов и сердечно-сосудистой системы
Первоначально основные сведения о наличии механорецепции в афферентных системах желудочно-кишечного тракта, органов дыхания, моче-половой системы, сердечно-сосудистой системы, органов кроветворения были получены в экспериментах, в которых показателем деятельности интероцепторов являлись рефлекторные изменения кровяного давления, дыхания, мочеотделения, активности скелетной и гладкой мускулатуры и т. д. Именно таким способом удалось установить существование самого факта чувствительности интероцепторов к действию механических раздражений (интеромеханорецепции) и проанализировать некоторые ее закономерности применительно к системам внутренних органов и отдельным органам (см. обзоры: Schweitzer, 1937; Быков, 1942; Черниговский, 1943в, 1960; Гамбашидзе, 1951; Айрапетьянц, 1952; Курцин, 1952; Черниговский, Ярошевский, 1953; Aviado, Schmidt, 1955; Могендович, 1957; Heymans, Neil, 1958; Булыгин, 1959, 1966; Меркулова, 1959; Фролькис, 1959; Хаютин, 1964; Кулаев, 1972, и мн. др.). Следует признать, что указанный подход, исчерпывающе разработанный В. Н. Черниговским и его сотрудниками, несомненно является одним из наиболее адекватных при оценке функциональной значимости афферентной сигнализации, поступающей от сенсорных аппаратов различных интероцептивных областей.
Применение электрофизиологических методов для изучения свойств интероцепторов и связанных с ними нервных провод
ников, начатое еще в работах Эдриана’ (Adrian, Umrath, 1929; Adrian, 1933), было широко использовано многими авторами (см. обзоры: Черниговский, 1960, 1967; Paintai, 1963а, 1963Ь. 1972, 1973; Iggo, 1966b; Sharma, 1967; Iggo, Leek, 1970; Булыгин, 1971; Бакунц, 1972; Leek, 1972; Глебовский, 1973; Булыгин, Ка- люнов, 1974; Ноздрачев, 1974, и др.). Разработка встретила большие методические трудности: основная часть нервных проводников, связанных с интероцепторами, относится к тонким миелинизированным и немиелинизированным волокнам. Поэтому вначале многие работы были выполнены методикой регистрации суммарной импульсной активности нервных стволов (Tower, 1933; Evans, 1936; Talaat, 1937; Gernandt, Zotterman, 1946; Адамович, 1953; Делов и др., 1953, 1960; Замятина^ 1954, 1957; Аникина, 1956, 1960а, 19606; Лагутина, 1957, 1958, 1959; Пинес, 1960; Сиротин, 1960, и мн. др.), и только в отдельных исследованиях удалось охарактеризовать свойства одиночных рецепторов, которые, как правило, были связаны с относительно толстыми нервными волокнами (Bronk, Stella, 1932, 1935; Gammon, Bronk, 1935; Gernandt, Zotterman, 1946; Knowlton, Larrabee, 1946; J. A. B. Gray, Malcolm, 1950; Neil, 1954, и др.). Лишь постепенно, благодаря усовершенствованию методов экспериментирования с тонкими нервными волокнами (Paintai, 1954а; Iggo, 1955, 1957b) , в практику исследований импульсной активности интероцепторов широко вошло изучение импульсации, передаваемой по одиночным нервным проводникам. Начинает использоваться и микроэлектродная техника (детали этой техники см., например: Лев, 1964; Скок, 1970; Crowcroft et al., 1971), позволяющая регистрировать афферентную импульсную активность от интероцепторов в структурах спинальных и вегетативных ганглиев (Mei, 1970а, 1970b, 1970с; Качалов, Ноздрачев, 1972; Arlhac 1972; Ranieri et al., 1973, и др.). Все эти исследования позволяют оценить природу адекватного стимула, пороги чувствительности, величину рецептивного поля, измерить скорость проведения в связанных с механорецепторами нервных волокнах и т. д.
Несомненным шагом вперед в изучении свойств интероцепторов явились также работы, в которых импульсная активность хотя и отводилась от целых нервных стволов, однако эксперименты эти проводились в хронических условиях, т. е. в условиях, значительно более приближенных к реальной жизни животных (Ноздрачев, 1969; Черниговский и др., 1972; Климов и др., 1973).
Как уже неоднократно упоминалось, многие важные факты, касающиеся общих механизмов деятельности механорецепторов, получены в экспериментах на одиночных мезентериальных тельцах Пачини (см. обзоры: J. А. В. Gray, 1959а; Loewenstein, 1966, 1971; Sato, Ozeki, 1966; Ильинский, 1967, 1971). Эти данные были рассмотрены отдельно (см. стр. 75, 117).
Виды механорецепторов. Рецепторы, чувствительные к действию механических стимулов, обычно подразделяют в зависимости от места их локализации, т. е. на механорецепторы желудочно-кишечного тракта, органов дыхания и т. д. При этом естественно, что характерный рисунок импульсной активности механорецепторов существенно зависит от специфических особенностей деятельности различных органов и систем.
Кроме того, вне зависимости от места расположения рецепторы делят на медленно и быстро адаптирующиеся (соответственно тонические и фазные). Первые отвечают стационарной активностью в течение всего времени действия раздражителя. Эти ответы могут иметь хорошо выраженную динамическую фазу и обязательную статическую фазу. Величина динамической фазы определяется амплитудой и скоростью нарастания сигнала, а величина статической фазы зависит от интенсивности статической составляющей воздействия. Импульсная активность от рецепторов второго типа регистрируется лишь при включении и выключении раздражения, но не наблюдается в период действия его статической части.
К быстро адаптирующимся (фазным) механорецепторам внутренних органов относятся мезентериальные тельца Пачини (Adrian, Umrath, 1929; Sheehan, 1932; Gammon, Bronk, 1935; Gernandt, Zotterman, 1946; J. A. B. Gray, Malcolm, 1950; Ильинский, 1963, и др.), рецепторы движения (movement receptors), локализованные под серозной оболочкой тонкой кишки (Besson, Perl, 1966), рецепторы слизистой кишечника (Paintai, , рецепторы мукозного слоя желудка (IggQ, 1957b; Niijima, 1962), часть из которых может рассматриваться и как хеморецепторы, так как они реагируют на изменения pH (Iggo, 1957b), рецепторы течения (flow receptors) в уретре (Iggo, 1956; Todd, 1964), рецепторы напряжения в*уретре (Todd, 1964), ирритантные рецепторы легких (Mills et al., 1970) ит. д. Медленно адаптирующиеся рецепторы есть также во всех внутренних органах. Это, например, рецепторы растяжения легких (Adrian, 1933; Knowlton, Larrabee, 1946; Widdicombe, 1954a, 1954b, 1964), каротидные барорецепторы (Bronk, Stella, 1932, 1935; Landgren, 1952a), рецепторы напряжения или растяжения в желудке (Paintai, 1954а, 1954b,,1963а; Iggo, 1957а) и в мочевом пузыре (Iggo, 1955) и т. д. Следует подчеркнуть еще раз, что описанные типы рецепторов были выделены лишь на основании рисунка их импульсной активности, а не по каким-либо другим признакам.
Явления адаптации (оцениваемые по изменениям кровяного давления и дыхания), наступающие при раздражении механорецепторов внутренних органов, были предметом специальных исследований В. Н. Черниговского и его сотрудников (Черниговский, 1949, 1960; Уголев и др., 1950; Хаютин, 1952; Хаютин, Черниговский, 1952). В этих экспериментах удалось показать, что в изменениях системных реакций, наблюдаемых в ходе механического раздражения интероцепторов, ведущую роль играют процессы, протекающие в центрах, а не на периферии [см, также
Медиальный лемниск

Рис. 42. Схема распределения висцеральных чувствительных нервов и их связь с сегментами спинного мозга кошки (по: Rasmussen, 1932).
Слева — нервы, отходящие от внутренних органов; справа — сегменты спинного мозга и входящие в них нервы. 1 — шейный, II — грудной, III — поясничный, IV — сакральный отделы спинного мозга. 1 — язычок; 2 — нёбо; 3 — глотка; 4 — язык; 5 — трахея; в — легкие; 7 — сердце; 8 — диафрагма; 9 — печень; ю — желчный пузырь; н — желудок; 12 — селезенка; 13 — поджелудочная железа; 14 — кишечник; 15 — почка; 16 — яичник; 17 — матка; 18 — мочевой пузырь; 19 — мочеиспускательный канал; 20 — прямая кищка,
электрофизиологические исследования Лагутиной (1958, 1959)]. Очевидно, что эти данные выходят за рамки настоящей книги и поэтому специально здесь не рассматриваются. Ряд важных фактов, касающихся механизмов работы самих механорецепторов, полученных в этих работах, будет рассмотрен далее, в главе 11.
Характеристика афферентных висцеральных нервных волокон. Рассмотрению свойств афферентных висцеральных проводников в литературе посвящено много работ (см. обзоры: Черниговский, 1960; Ruch et al., 1961; Paintai, 1963a, 1972, 1973; Дуринян,
1965;
Булыгин, 1966, 1971; Leek, 1972; Глебовский, 1973;
Муся- щикова, Черниговский, 1973;
7000-М
27000=
НМ


Рис. 43. Количество миелинизированных (М) и немиелинизи- рованных (НМ) волокон в блуждающем нерве (по: Paintai. 1963а).
— корешки блуждающего нерва:
— языкоглоточный нерв; з — предсердная ветвь; 4 — яремный ганглий;
5 — глоточный нерв; в — узловатый ганглий; 7'— верхнегортанный 'нерв;
8 — блуждающий "нерв (шейный отдел); 9"—^возвратная ветвь нижнегортанного нерва; 10 — сердечные ветви;
11 — нервные ветви к плечевому сплетению; 12 — блуждающий нерв (поддиафрагмальный отдел).
ются блуждающие (nn. vagi), чревные (nn. splanchnici) и тазовые нервы (nn. pelvici). Конечно, имеются и иные афферентные пути (например, языкоглоточные нервы и др.), однако, они в значительно меньшей степени участвуют в передаче сигналов от внутренних органов. Схема иннервации висцеральными нервами внутренних органов, а также распределения их в сегментах спинного мозга кошки представлена на рис. 42.
Как показывают данные многих авторов, висцеральные нервы содержат большое количество афферентных волокон. Так, в блуждающем нерве млекопитающих число афферентных проводников может составлять 80—90% от общего количества волокон, равного примерно 30 тысячам в шейном отделе блуждающего нерва у кошки. При этом из них лишь примерно 20% составляют миелинизированные волокна, остальные волокна безмякотные (рис. 43) с диаметром менее 1—2 мкм (Jones, 1932; Foley, 1934; DuBois, Foley, 1937; Foley, DuBois, 1937; Daly, Evans, 1953; Agostoni et al., 1957; Стовичек, 1966). В чревном нерве количество афферентных проводников превышает 50% (Foley, 1948), а в тазовом нерве достигает порядка 30% (Ranson, 1921). Весьма возможно, что и это число является заниженным, так как основную массу волокон составляют немиелинизированные проводники, подсчет которых подчас бывает затруднительным. Таким образом, следует констатировать, что висцеральные нервы являются в, значительной степени сенсорными.
Афферентные миелинизированные волокна в висцеральных нервах представлены в основном группой АВ и в меньшей степени волокнами Ар (рис. 44). В блуждающем нерве афферентные волокна Ар связаны в основном с мышцами гортани и глотки. Следовательно, можно считать, что они выполняют афферентные

Рис. 44. Распределение миелинизированных афферентных волокон по их диаметрaMj(no: Ruch et al., 1961).
а — блуждающий нерв (количество волокон — 1674); б — чревный нерв (1379); в — тазовый нерв (6658). По оси абсцисс — диаметр волокон (мкм); по оси ординат — отношение числа волокон данного диаметра к общему числу волокон, принятому за 100 (%).
функции соматических проводников. В легких же наиболее толстые волокна связаны с медленно адаптирующимися рецепторами растяжения. Скорость их проведения составляет около 59 м/сек. (Paintai, 1963а). В чревном нерве наиболее толстые волокна связаны прежде всего с тельцами Пачини, однако и другие механорецепторы в кишечнике могут быть связаны с такого рода проводниками (Gernandt, Zotterman, 1946; Amassian, 1951; Downman, Evans, 1957; McLeod, 1958). Толстые волокна тазового нерва, возможно, связаны также с образованиями типа телец Пачини, локализованными, например, в стенке мочевого пузыря (см.: Дуринян, 1965).
Произвести расчет скоростей проведения в висцеральных афферентных проводниках на основе знания диаметра, равно как и обратная задача — оценить диаметр волокна на основе измерения скорости проведения — не всегда просто. Это объясняется, помимо трудностей общего характера (см. стр. ИЗ), тем, что ряд висцеральных нервных: волокон по своему ходу могут существенно изменять диаметр и степень миелинизации (стр. 26).
Так, например, в интраторакальной части блуждающего нерва для большинства кардио-васкулярных волокон отмечается снижение скорости проведения на 10—40% сравнительно со скоростью проведения в шейном отделе блуждающего нерва. Для 10% волокон это снижение достигает даже 50% (Paintai, 1962). Данное заключение справедливо для вагусных волокон со скоростями проведения свыше 10 м/сек.; нервные проводники с меньшими скоростями проведения в этом плане не исследовались (Paintai, 1972).3
Активность механорецепторов сердца и сосудов
Исследователи проявляют большой интерес к изучению механорецепторов в системе кровообращения (см. обзоры: Черниговский, 1943в, 1960; Heymans, Neil, 1958; Фролькис, 1959; Удельнов, 1961; Paintai, 1963а, 1972; Хаютин, 1964; Кулаев, 1972, и др.).
К настоящему времени выполнено большое количество электрофизиологических работ, посвященных исследованию этих механорецепторов, а также изучению свойств их волокон (Bronk, Stella, 1932, 1935; Анохин, Шумилина, 1947; Буйя, 1948; Paintai, 1953а, 1953b, 1953с, 1963b; Irisawa, Ninomiya, 1967; Angell James, 1968; Koushanpour, McGee, 1969; Angell James, Daly, 1971; Анюховский и др., 1974; Arndt et al., 1974; Gilmore, Zucker, 1974, ими. др.). В последнее время стали появляться работы, в которых наряду с импульсной активностью сосудистых механорецепторов начинают регистрировать и РП (Matsuura, 1973).
Афферентные проводники от механорецепторов сердечно-сосудистой системы исследовались весьма подробно. Как правило, скорость проведения по афферентным кардиоваскулярным мякот-
Интересно, что увеличение в проксимальном направлении диаметра мякотных афферентных волокон блуждающего нерва можно отметить и у низших позвоночных (Ильинский и др., 1971). Такого рода факты говорят о том, что при изучении висцеральных проводников (особенно, небольшого диаметра), по-видимому, следует соблюдать известную осторожность в оценках скорости проведения (И) или диаметра (/)) на основе одного лишь из этих показателей. В этой связи можно указать, что, например, измерение скорости проведения в волокнах от фазных механорецепторов (рецепторов движения), связанных с брыжеечными нервами, показало, что она варьирует от 2 до 21 м/сек. (в среднем 5—10 м/сек.), тогда как, по данным гистологических исследований, в изучаемых нервах были выявлены тонкие мякотные волокна, диаметры которых варьировали от 2 до 8 мкм (в большинстве случаев — порядка 3 мкм) (Bessou, Perl, 1966). Следовательно, в этом случае V равняется 14-3 D, но не 6 и даже не 4 D.
Таблица 2
Скорость проведения возбуждения по афферентным волокнам рецепторов сердечно-сосудистой системы (сводные данные, по: Paintai, 1972)
Тип рецепторов |
Афферецтные волокна |
Скорость проведения (м/сек.) |
Каротидные барорецепторы | Аортальные барорецепторы Рецепторы правого предсердия (тип А) Рецепторы левого предсердия (тип А) Рецепторы правого предсердия (тип В) Рецепторы левого предсердия (тип В) Вентрикулярные рецепторы давления Эпикардиальные рецепторы Перикардиальные рецепторы |
Миелинизированные Немиелинизирован- ные (группа С) Миелинизированные » » » » » Немиелинизирован- ные (группа С) Миелинизированные |
Неизвестна 0.5—2.0
12—19 8-29 11-26 8—19 0.4—5.0 2.5—7.0 |
ным нервным волокнам превышает 10 м/сек. (табл. 2). В волокнах сердечных барорецепторов она варьируется от 8 до 29 м/сек. (Paintai, 1972; см. также: Mei, 1970а, 1970Ь), аортальных барорецепторов — от 10 до 55 м/сек., с максимумом 25—30 м/сек. (Paintai, Riley, 1966) или 22—32 м/сек. (Mei, 1970а). Волокна рецепторов перикарда имеют скорость проведения 2.5—7.0 м/сек. (Sleight, Widdicombe, 1965b) или 18—23 м/сек. (Mei, 1970а).
Изучение состава аортального нерва кошки (Devanandan, 1964) показало, что основная часть волокон имеет диаметр менее 4— 6 мкм, и лишь небольшое количество волокон достигает диаметра 10 мкм (максимальная скорость проведения 64 м/сек.). Сходные данные были получены при исследовании аортального нерва свиньи (Schmidt, Stromberg, 1967). По-вйдимому, наиболее толстые и средние волокна в нервах сердечно-сосудистой системы связаны с механорецепторами, а тонкие волокна — прежде всего с хеморецепторами (De Castro, 1951; Paintai, 1972). Тем не менее в каротидном нерве примерно 30% немиёлинизированных волокон со скоростями проведения 0.5—2 м/сек. принадлежат также барорецепторным элементам (A. Sato et al., 1968; Fidone, Sato, 1969). Однако в аортальном и блуждающем нервах барорецепторных немиелинизированных волокон либо нет вообще, либо количество их незначительно (Paintai, 1972).
Механорецепторы сердечно-сосудистой системы часто называют в литературе барорецепторами. Этот термин отражает старую точку зрения, согласно которой именно давление крови является фактором, возбуждающим сосудистые механорецепторы. Однако в настоящее время стало очевидным (Heymans, Neil, 1958; Черниговский, 1960), что возбуждение механорецепторов сосудов вызывается не давлением как таковым, а обусловленным им растяжением сосудистых стенок. Следует указать, что вообще возбуждение самых разных тканевых механорецепторов вызывается именно растяжением, которое приводит к увеличению площади поверхности чувствительной мембраны нервных окончаний (Ильинский, 1967; см. также главу 11).
Синокаротидные барорецепторы. Систематическое изучение деятельности механорецепторов сосудов с помощью электрофизиологических методов было начато уже в 30-х годах (Bronk, 1931; Bronk, Stella, 1932, 1935) на механорецепторах каротидного синуса. Эти рецепторы являются медленно адаптирующимися, и их стационарная активность возрастает линейно по мере увеличения уровня постоянного давления. Возрастание импульсации происходит до некоторой величины — уровня насыщения, который варьирует у разных рецепторов; обычно он достигается при давлении не более 200 мм рт. ст. (чаще всего при 140— 180 мм рт. ст.). Однако для возбуждения рецепторов наиболее эффективным оказалось не постоянное давление, а его нарастание, обусловленное сердечным циклом. Каждая систола вызывала появление пачки импульсов, а в период диастолы импульсация пропадала. Пороги возбуждения различных рецепторов варьировали.
Дальнейшие исследования полностью подтвердили приведенные выше факты. Так, было показано (Ead et al., 1952), что частота импульсной активности барорецепторов при действии постоянного давления ниже, чем при действии давления, вызванного пульсирующим раздражением. При этом пульсирующее воздействие вовлекало в процесс возбуждения большее количество рецепторов.
Было проведено детальное исследование свойств одиночных механорецепторов каротидного синуса кошки (Landgren, 1952а, 1952Ъ). Согласно полученным результатам, активность рецепторов изменяется в определенном диапазоне исследовавшегося постоянного давления. При давлениях 50—70 мм рт. ст. импульсная активность рецепторов обычно отсутствовала. Пороговое давление, необходимое для получения устойчивой импульсации в рецепторах, генерировавших большие спайки, равнялось 80— 120 мм рт. ст., а в рецепторах, отвечающих спайками малой амплитуды, — 120—150 мм рт. ст. (соответственно, волокна групп А и С — Fidone, Sato, 1969). При одной и той же величине воздействия рецепторы первого типа имели более высокую частоту разрядов, чем рецепторы второго типа (250—350 и 50—150 имп./сек. соответственно). В ответ на резкое повышение давления рецепторы отвечали всплеском активности, которая постепенно уменьшалась до некоторого фонового значения, характерного для данного уровня постоянной составляющей давления. Чем выше была скорость изменения давления, тем значительнее было первоначальное изменение частоты импульсации. Активность рецепторов после снятия раздражения угнеталась, как это было описано еще в ранних исследованиях (Bronk, Stella, 1932).
Было отмечено существование так называемых парадоксальных ответов некоторых барорецепторов (Landgren, 1952а). Эти ответы возникали при снижении давления в каротидном синусе от исходного сравнительно небольшого уровня (60—20 мм рт. ст.) до нуля. Подобного типа разряды сосудистых механорецепторов в дальнейшем наблюдали и другие авторы (Wiemer, Ott, 1963). Причиной такого рода «отрицательных» разрядов 4 исследователи считают деформацию механорецепторов, обусловленную спадением сосудистых стенок.
Аналогично тому, как это в свое время сделали первые исследователи (Bronk, Stella, 1932, 1935), объяснявшие полученные ими результаты на основе данных, установленных в опытах с мышечными веретенами, Ландгрен также привлек для трактовки своих фактов результаты исследований, проведенных на других рецепторных образованиях, а кроме того, и концепцию Гранита и его школы о генераторном потенциале. В частности, Ландгрен считает возможным объяснить полученные им результаты на основе тех данных, которые были обнаружены Катцем (Katz, 1950b) в опытах на мышечных веретенах лягушки. Именно на этом объекте впервые были описаны РП механорецепторов и показано, что чем больше величина раздражения, тем значительнее амплитуда локальной деполяризации, величина стационарной фазы которой в свою очередь линейно связана с частотой импульсации. Кроме того, было показано существование гиперполяризационного off-РП, а также отмечена роль фактора скорости нарастания раздражения. Постулирование существования такого же рода явлений и в барорецепторах позволило Ландгрену (Landgren, 1952а) полностью объяснить все полученные им результаты. Следует указать, что этот автор обратил внимание и на возможную роль механических факторов (вязких свойств рецепторных приборов и окружающей их эластической ткани).
Таким образом, уже первые исследования, посвященные изучению свойств барорецепторов, показали, что все особенности импульсной активности этих структур могут быть объяснены на основе общих закономерностей деятельности механорецепторов. Только некоторые детали характеристик реакций барорецепторов, по-видимому, были связаны с механическими свойствами вспомогательных структур и окружающих рецепторы тканей. Последующие работы подтвердили этот вывод (см., например, обзор: Paintai, 1972).
Аортальные и брахиоцефальные барорецепторы. Барорецепторы других артерий. Эти сенсорные приборы обладают в целом свойствами, сходными с наблюдаемыми в каротидных барорецепторах (Whitteridge, 1948; Neil, 1954; Bloor, 1964).
Правильнее было бы говорить «разряды на отрицательные изменения давления».
Основной особенностью аортальных барорецепторов является генерация систолической вспышки импульсов (рис. 45) (примерно через 80 мсек, после зубца Q ЭКГ). В экспериментах на изолированной дуге аорты было показано (Angell James, 1971а, 1971b, 1971с), что рецепторы обладали способностью генерировать стационарную импульсацию, величина которой линейно возрастала по мере увеличения постоянного давления до некоторого максимального значения (давление колебалось для разных рецепторов
Ь
I 1
Рис.
45. Импульсная активность одиночного
волокна левого нерва аорты (верхняя
запись)
и кровяное давление в левой общей сонной
артерии (нижняя
запись)
(по: Neil, 1954).![]()
![]()
![]()
![]()
![]()



Среднее значение кровяного давления: А — 125, Б — 80, В — 62, Г — 55, Д — 42 мм рт. ст. Отметка времени — 1 сек.
от 60 до 160 мм рт. ст.). При переходе к пульсирующему воздействию непрерывная импульсация разбивалась на пачки.
Наличие артериальных барорецепторов отмечено в легочной артерии (Coleridge, Kidd, 1960, 1961; Nishi et al., 1974), а также во многих внутренних органах (Черниговский, 1943в, 1960; Niijima, Winter, 1968; Niijima, 1971, и др.).
В зависимости от различных условий (например, гипертонии) механорецепторы сосудов, а также и сердца могут изменять свою деятельность (Анюховский и др., 1974). В перенастройке работы механорецепторов, по-видимому, участвуют различные факторы, например, изменение механических свойств стенки сосудов (Abraham, 1967; Hilgenberg, 1967) и регулирующее влияние симпатической иннервации (Sampson, Mills, 1970; Bagshaw, Peterson, 1972).
Механорецепторы сердца. В настоящее время описано два основных типа рецепторов в предсердиях — А и В (Paintai, 1953а, 1963а, 1963Ь, 1972). Рецепторы типа А генерируют только одну пачку импульсов в ходе сердечного цикла, совпадающую с первой волной пульсового изменения предсердного давления. Рецепторы типа В возбуждаются синхронно со второй волной пульсового изменения предсердного давления. Латентное время вспышки импульсов относительно волны Q ЭКГ равняется у кошки 70—210 (обычно 140) мсек. Разряд рецепторов типа В нарастает по частоте во времени, чем он отличается, например, от большинства аортальных барорецепторов, разряды которых обладают начальным максимумом активности. По своей адаптационной способности оба типа рецепторов предсердий могут рассматриваться как медленно адаптирующиеся. Показано, что активность рецепторов типа В линейно связана с объемом жидкости в предсердии (Paintai, 1953а).
Рецепторы типа В у кошки локализуются преимущественно в задней части стенки предсердия. Возможно, что рецепторы типа А и В отличаются лишь своим различным расположением в предсердии.
Рецепторы предсердия подобно другим^типичным висцеральным механорецепторам, таким как рецепторы растяжения в легких (Adrian, 1933), каротидные барорецепторы (Bronk, Stella, 1935), рецепторы растяжения желудка (Paintai, 1954b) и другие механорецепторы внутренних органов (Черниговский, I960), мало чувствительны к аноксии.
В стенке желудочка описаны ^прессорецепторы^ образованные миелиновыми волокнами (Paintai, 1955). Описаны также и эпикардиальные рецепторы, иннервируемые немиелинизированными волокнами и чувствительные к ряду химических агентов (Coleridge et al., 1964; Sleight, Widdicombe, 1965a). Такого же рода образования отмечены и в перикарде (Черниговский, 1941, 1943а, 19436; Sleight, Widdicombe, 1965b).
Активность механорецепторов желудочно-кишечного тракта
Анализу деятельности механорецепторов желудка, различных отделов кишечника, брыжейки посвящено большое количество исследований (см. обзоры: Курцин, 1952; Черниговский, 1960, 1967; Булыгин, 1966, 1971; Leek, 1972; Итина, 1972; Ноздрачев, 1974; Newman, 1974). Во многих работах исследовалась суммарная активность большого числа рецепторов, оцениваемая к тому же часто по реакциям кровяного давления, дыхания, изменению мочеотделения и т. д. И лишь сравнительно небольшое число исследований (если не считать опытов на мезентериальных тельцах Пачини) было выполнено с помощью методики регистрации активности от одиночных нервных волокон (Paintai, 1954b, 1957; Iggo, 1955, 1957а, 1966b; Niijima, 1960, 1961, 1962; Besson, Perl, 1966; Iggo, Leek, 1967a, 1967b, 1970; Andrews et al., 1972; Davison, 1972; Morrison, 1973, и др.)£илиь одиночных нейронов в ганглиях (Mei, 1970а, 1970с; Crowcroft et al., 1971; Качалов, Ноздрачев, 1972; Ranieri et al., 1973).
Афферентные волокна. Ряд авторов изучали характеристики афферентных^волокон, связанных£с органами пищеварительного канала. Так, было показано, что волокна блуждающего нерва от механорецепторов пищевода проводят возбуждение со скоростью 1—14 м/сек. (Mei, 1970а). Волокна механорецепторов желудка и кишечника кошки, входящие в состав блуждающего нерва, характеризуются скоростью проведения 6.5—13 м/сек. (в среднем 9 м/сек.) (Paintai, 1954b). По другим данным (Iggo, 1958), волокна, связанные с рецепторами желудка и кишечника, обладают меньшей скоростью проведения — порядка 1 м/сек. (от 0.5 до 2.5 м/сек.) или 4.7 м/сек. (Mei, 1970а).
В дистальных отделах тонких мякотных волокон скорость проведения меньше, чем в более проксимальных участках. Это, по-видимому, связано с потерей мякотной оболочки некоторыми нервными проводниками (возможно, в грудном отделе — см. выше). Величины скоростей проведения по волокнам, связанным с механорецепторами желудка, в опытах Игго были меньше, чем в опытах Пайнтала, что может быть обусловлено различиями в технике измерений. Учитывая небольшие диаметрьг волокон блуждающего нерва, можно думать, что данные Игго, по-видимому, являются более правильными.
У жвачных животных было установлено, что механорецепторы рубца (одной из камер преджелудка) связаны с более толстыми миелиновыми волокнами. Их скорость проведения была равна 2—12 м/сек. (Iggo, 1955), а по другими данным — в среднем 12.4 м/сек. (Leek, 1969).
По данным, полученным на жабе, диаметр волокон, иннервирующих^ желудок, равняется 3—14 мкм (Niijima, 1960, 1962). Эти величины сходны с аналогичными величинами, полученными для бета- и дельта-волокон в кишечнике у кошки (сравн.: Ger- nandt, Zotterman, 1946). Были также описаны медленно и быстро адаптирующиеся механорецепторы желудка у амфибий (Niijima, 1960, 1962).
Наиболее толстые мякотные волокна в мезентериальных нервах берут начало от телец Пачини. Диаметры этих волокон обычно колеблются от 3 до 13 мкм (Edgeworth, 1892; Sheehan, 1933; Gernandt, Zotterman, 1946; Glees et al., 1949; Quilliam, Sato, 1955; McLeod, 1958; Черепнов, 1968a). Скорости проведения по волокнам указанного типа достаточно велики и варьируют от 13 до 70 м/сек. (Downman, Evans, 1957; Ильинский, 1963; Leitner, Perl, 1964; Ranieri et al., 1973, и др.). О других фазных механорецепторах (рецепторах движения) и их волокнах говорилось выше (стр. 136) (данные из работы: Besson, Perl, 1966).
В опытах с растяжением' и сдавливанием кишечника было установлено, что возникновение импульсной активности наблюдается в мезентериальных волокнах групп AS и С (Gernandt, Zotterman, 1946). Электронномикроскопическое изучение аналогичных тонких веточек мезентериального нерва, иннервирующих тонкий кишечник (исследовались как афферентные, так и эфферентные проводники), показало, что они состоят из большого количества немиелинизированных волокон, диаметр которых варьировал от 0.2 до 2.2 мкм (с преобладанием волокон диаметром 0.2—0.8 мкм), а также тонких миелинизированных волокон диаметром от 0.6 до 6.2 мкм (максимум распределения в области 1—3.6 мкм). Скорость проведения по немиелинизированным волокнам (группа С) составляла 0.4—2.5 м/сек., а по мякотным волокнам (группа AS) — 5.3—11.9 м/сек. (Козлова и др., 1969; Лебедева Л Козлова, 1971; Лебедева, Филиппова, 1972). Сходные данные были получены и в других работах (Булыгин, Солтанов, 1969; Mei, 1970а). Можно сравнить эти данные также с данными о скоростях проведения по немиелинизированным волокнам других‘ висцеральных проводников, например, в сердечных и почечных нервах, где скорость проведения составляла 0.7 и 0.6 м/сек. (Катунский, Хаютин, 1968), нервах от каротидных барорецепторов (0.5—2.0 м/сек. — Fidone, Sato, 1969), и др.
Медленно адаптирующиеся рецепторы. Исследование импульсации, передаваемой по чувствительным волокнам от отдельных структур желудочно-кишечного тракта, позволило описать большое количество различных механорецепторов. В мышечных слоях ряда органов: в пищеводе (Andrew, 1957), желудке кошки (Paintai, 1954а, 1954b; Iggo, 1957а; Ranieri et al., 1973), кишечнике кошки (Iggo, 1957а; Ranieri et al., 1973), преджелудках коз (Iggo, 1955) и овец (Leek, 1969) были описаны медленно адаптирующиеся ме- ханорецепторные системы (stretch receptors у Пайнтала и tension receptors у Игго), включенные последовательно с мышечными структурами.
В тех случаях, когда моторная активность указанных выше органов отсутствует, в этих рецепторах наблюдается фоновая, обычно нерегулярная активность, частота которой пропорциональна исходному напряжению стенок. Наличие разнопороговых рецепторов приводит к тому, что только часть из них в покоящемся органе имеет исходную^активность. Изменение частоты разряда рецептора при пассивном растяжении стенок органа зависит от скорости растяжения, возрастая при ее увеличении (Paintai, 1954b; Iggo, 1955). При резком воздействии обнаруживается выраженная динамическая составляющая ответа, которая довольно медленно (например, за 1 мин.) переходит в статический компонент. Величина статического компонента (частота импульсации) соответствует новому стационарному уровню растяжения.
При сокращении гладкой мускулатуры органа в изотонических условиях, когда нет изменения напряжения стенки органа, но уменьшается его^объем, частота импульсации может практически не изменяться. Напротив, в изометрическом режиме, когда при сокращении мышц объем полого органа не изменяется, но напряжение стенки растет, наблюдается большая частота импульсации (Iggo, 1955; Leek, 1969).
Были также описаны характеристики импульсации, свидетельствующие о наличии медленно адаптирующихся рецепторов сальника (Iggo, 1957b). Медленно адаптирующиеся механорецепторы являются окончаниями А8- и С-волокон, ’входящих в состав чревного нерва. Они были обнаружены в малом сальнике, брыжейке желчного пузыря, в области портальной вены, поджелудочной железы, селезенки, в различных участках тонкого и толстого кишечника и их брыжейках (Morrison, 1973). Рецептивные поля одиночных волокон могли весьма варьировать. Некоторые из них, содержащие 1—2 чувствительные точки, были невелики (область корня брыжейки, портальной вены). Другие же (в области брыжейки тонкого кишечника) могли содержать до 7 чувствительных участков, расположенных друг от друга на расстоянии до 40 мм. В ответе рецепторов на раздражение хорошо были видны динамическая и статическая составляющие. Тоническая составляющая ответа могла продолжаться более 1 мин. Рецепторы были мало чувствительны к действию кислоты, гипоксии и гиперкапнии.
Быстро адаптирующиеся рецепторы. К ним относятся тельца Пачини, активность которых исследовалась многими авторами не только в опытах in vitro, но и in vivo (Gammon, Bronk, 1935; Gernandt, Zotterman, 1946; J. A. B. Gray, Malcolm, 1950; Замятина, 1954, 1957; Аникина, 1956, 1960a; Ильинский 1963, 1966a; Ильинский, Лебедева, 1963; Leitner, Perl, 1964). Тельца Пачини часто формируют фоновую активность в ритме пульсовых толчков артериального давления.
К быстро адаптирующимся механорецепторам относятся рецепторы движения (movement receptors), обнаруженные под серозной оболочкой тонкого кишечника и среди мезентериальных сосудов кошки (Bessou, Perl, 1966). Эти рецепторы не реагировали на статическую составляющую воздействия, но хорошо отвечали в моменты включения и выключения раздражения. Частота их разряда при раздувании кишки хорошо коррелировала со скоростью изменения растяжения и величиной давления в конце динамической фазы стимула. Рецепторы движения не могли отвечать на синусоидальную вибрацию уже при частотах 100 гц, и их реакция на низкочастотное ритмическое воздействие (30 колебаний в 1 сек.) исчезала очень быстро — в течение первых секунд раздражения. Большинство исследованных волокон обладали скоростью проведения 5—10 м/сек., т. е. являлись тонкими мякотными волокнами (основной диаметр у большинства — порядка 3 мкм). Величина рецептивного поля волокон часто не превышала 1 мм2, но могла распространяться вдоль сосуда на расстояние порядка 1 см. Авторы полагают, что эти рецепторы активируююя в результате слабых механических перемещений
в брюшной полости, вызванных изменением позы, дыханием, деятельностью сердечно-сосудистой системы или движением кишечника.
Существование быстро адаптирующихся механорецепторов было обнаружено в слизистой тонкого кишечника (Paintai, 1957; Niijima, 1962). Активность такого типа рецепторов наблюдается в виде вспышек и увеличивается при движении содержимого кишечника. Считают, что они располагаются в ворсинках слизистой (Paintai, 1957).
Имеются также чувствительные волокна, рецепторы ^которых расположены в слизистой желудочно-кишечного тракта и которые реагируют как на механические, так и на химические воздействия (Iggo, 1957b; Davison, 1972).
Активность механорецепторов легких
С помощью регистрации импульсной активности в чувствительных нервных волокнах было доказано наличие в различных структурах воздухоносных путей и в самой ткани легких большого количества механорецепторов. В зависимости от локализации различают рецепторы растяжения, связанные прежде всего с гладкомышечными структурами стенок воздухоносных путей, рецепторы слизистой трахеи и бронхов, рецепторы интерстициальной ткани легких, лежащие вблизи капилляров альвеол (их еще называют юкстакапиллярные, или J-рецепторы — juxta-pulmonary capillary receptors), и ряд других (Widdicombe, 1954а, 1964, 1974; Виноградова, 1959, 1960; Лашков, 1963; Paintai, 1963а, 1969, 1970; Mills et al., 1970; Fillenz, Widdicombe, 1972; Глебовский, 1973; Boushey et al., 1974, и др.).
145
Рецепторы растяжения легких. Основным типом структур являются разветвления достаточно толстых волокон в гладких мышцах стенок воздухоносных путей (Widdicombe, 1954b, 1964). Их называют даже веретенами гладких мышц (Elftman, 1943), хотя по своему строению рецепторы больше напоминают аналогичные разветвления в каротидном синусе, дуге аорты и камерах сердца (Sunder-Plassmann, 1933; Hayashi, 1937). Ответы именно этих рецепторов легких и были первыми зарегистрированными в виде импульсной активности афферентных нервных волокон блуждающего нерва (Adrian, 1933). Деятельность рецепторов растяжения в легких исследована наиболее подробно. Показано, что они являются медленно адаптирующимися структурами (рис. 46). Активность их возникает или учащается при вдохе. Максимальная частота разряда у кошки может доходить до 100—300 имп./сек. при давлении вдуваемого в легкие воздуха до 30 см вод. ст.
Увеличение импульсной активности отмечается в процессе поступления воздуха в легкие. Затем частота импульсации снижается, и эта фаза ответа может быть весьма выраженной. Снижение активности, возможно, связано не только с адаптационными свойствами самих рецепторов, но и с механическими особенностями ткани, в которой они располагаются. Так, рецепторы, локализованные в более крупных бронхах, отличаются более медленной адаптацией.
Рецепторы растяжения характеризуются весьма низким порогом амплитудной чувствительности. Наиболее чувствительная часть из них, обладающая фоновой активностью, тормозится только после снижения объема воздуха в легких ниже величины функциональной остаточной емкости. Частота импульсации рецепторов при умеренных растяжениях находится в линейной зависимости от величины объема воздуха (Adrian, 1933). Рецепторы растяжения мало чувствительны к асфиксии, действию углекислого газа.
Скорости проведения импульсов от медленно адаптирующихся рецепторов весьма велики: 14—59 м/сек. (Paintai, 1963а), 12— 50 м/сек. (Mei, 1970а).
Быстро адаптирующиеся рецепторы растяжения. К этой группе рецепторов (Knowlton, Larrabee, 1946), по-видимому, относятся ирритантные рецепторы эпителия воздухоносных путей (Fillenz, Widdicombe, 1972). Они связаны с волокнами меньших диаметров. Ряд авторовТуназывают для них следующие скорости проведения импульсов: 9 м/сек. (Knowlton, Larrabee, 1946), 25 м/сек. (Paintai, 1963а), 14—17 м/сек. (Mei, 1970а).
Рецепторы этого типа раздражаются парами едких веществ. Импульсная активность ирритантных рецепторов наблюдается в момент увеличения и уменьшения объема легких.
Ирритантные рецепторы (см. подробнее об’их свойствах: Fillenz, Widdicombe, 1972; Widdicombe, 1974) являются окончаниями




Рис. 46. Импульсная активность рецепторов трахеи (I) и легких (77) (I — схематизировано по: Widdicombe, 1954b; II — по: Paintai, 1969).
1 — ПД быстро адаптирующихся рецепторов трахеи. А — раздувание трахеи; Б — спадение трахеи; В — раздражение эпителия трахеи слабым прикосновением катетера. Сверху вниз: давление внутри трахеи (шкала справа); ПД рецепторов трахеи. Отметка времени — 0.1 сек. II — ответы J-рецепторов легких (маленькие двуфазные спайки) и рецепторов растяжения легких (большие моно- фазные спайки) при открытой грудной клетке на вдувание в легкие примерно 60 (А) или 150 мл (Б, В) воздуха (чувствительность регистрации раздражения на А в 2 раза меньше, чем на Б и Б). Сверху вниз: отметка раздражения; ПД рецепторов легких.
нервных волокон, разветвляющимися в эпителии воздухоносных путей от трахеи до респираторных бронхиол, и описаны многими авторами (Larsell, 1921; Fisher, 1964; Spencer, Leof, 1964). Они возбуждаются как при действии аэрозолей и газов, так и при механическом раздражении. Некоторые из этих. рецепторов, например расположенные в трахее, отвечают как на нанесение раздражения, так и на его устранение (рис. 46). В ряде случаев (при раздражении гортани и трахеи) механическое воздействие может вызывать кашель. Волокна, связанные с ирритантными рецепторами, проводят импульсы со скоростью 3.6—25.8 м/сек. (Mills et al., 1969, 1970).
Юкстакапиллярные рецепторы. Вначале они были обозначены как «специфические рецепторы спадения» (Paintai, 1955). Однако в дальнейшем это наименование было заменено термином «юкстакапиллярные рецепторы» (Paintai, 1969, 1970). Эти рецепторы связаны с волокнами, характеризующимися скоростями проведения от 0.8 до 7.0 м/сек. Возможно, что некоторые из этих волокон являются миелинизированными. Однако большинство из них проводит со скоростью менее 3.0 м/сек., что говорит, по- видимому, о том, что они скорее всего лишены миелиновой оболочки. Рецепторы этого типа быстро адаптируются. Ответы их не очень воспроизводимы при повторной стимуляции (рис. 46).
Выделяют еще медиастинальные рецепторы, возбуждаемые при движениях средостения (Adrian, 1933; Widdicombe, 1954а). Они являются медленно адаптирующимися структурами и связаны с мякотными волокнами.
Основные известные рефлексы с легких вызываются активностью рецепторов растяжения, ирритантных рецепторов и юкста- капиллярных рецепторов (Fillenz, Widdicombe, 1972).
Активность механорецепторов выделительной и половой систем
Как и в других внутренних органах, на основании экспериментов с регистрацией импульсной активности механорецепторы моче-половых органов могут быть подразделены на быстро и медленно адаптирующиеся (Evans 1936; Talaat, 1937; Хаютин, 1952; Адамович, 1954; Iggo, 1955, 1956, 1966b; Бокша, 1957; Лагутина, 1957, 1959; Niijima, 1959; Меркулова, Миркин, 1966, и др.).
Существование обоих типов рецепторов, отражающих различные виды механических усилий, возникающих в мочевом пузыре и в уретре, было продемонстрировано и с помощью регистрации активности одиночных элементов. Так, было показано наличие в мочевом пузыре рецепторов напряжения, которые можно рассматривать как включенные параллельно с мышечными элементами (Iggo, 1955, 1966b; Arlhac, 1972). Пример реакции такого типа рецепторов представлен на рис. 47. Рецепторы напряжения прежде всего реагируют на наполнение органа и его опорожнение. Частота их разрядов не очень высока (до 40 в 1 сек.) и значительно меньше, чем, например, для рецепторов течения в уретре (Iggo, 1955). Рецепторы напряжения, по-видимому, играют важную роль в рефлекторном управлении деятельностью мочевого пузыря (Iggo, 1966b). Колебания внутрипузырного давления хорошо улавливаются быстро адаптирующимися рецепторами (Arlhac, 1972).
В уретре были обнаружены рецепторы, получившие название рецепторов течения (Iggo, 1956; Todd, 1964). Интересной их осо-

Рис. 47. Изменение импульсации медленно адаптирующихся рецепторов растяжения при сокращении мочевого пузыря кошки (по: Iggo, 1966b).
1 — изменение объема мочевого пузыря; 2 — изменение давления внутри мочевого пузыря; з — частота импульсации одиночного волокна, выделенного из тазового нерва. По оси абсцисс — время (сек.); по оси ординат: для 1 — объем (мл), для 2 — давление (мм рт. ст.), для 3 — частота импульсации (имп./сек.).
бенностью является возбуждение лишь в условии протекания жидкости в уретре, и частота импульсов в их разряде (она могла достигать 300 в 1 сек.) была связана со скоростью течения. Постоянное растяжение стенки уретры было неэффективным, импуль- сация не возникала. По-видимому, рецепторы возбуждались при колебаниях стенки, возникающих в уретре при токе жидкости, который, возможно, носит вихревой характер. Помимо рецепторов течения были описаны быстро адаптирующиеся разряды импульсной активности, возникавшие либо в момент растяжения уретры, либо в момент прекращения воздействия (Todd, 1964). Эти ответы, вероятно, могут быть обусловлены мелкими пачиниподобными тельцами, находящимися в уретре. Кроме того, более медленно протекающие деформации уретры могли быть зарегистрированы медленно адаптирующимися рецепторами (Arlhac, 1972).
Медленно адаптирующиеся импульсные разряды были описаны в афферентных нервных волокнах почки и яичка (Niijima, 1959). Соответствующие рецептивные поля составляли примерно 5x5 мм2. В этих же органах отмечена активность быстро адаптирующихся рецепторов (Tower, 1933). Медленно адаптирующиеся механорецепторы были описаны также в матке (Bower, 1959, 1966).
* * *
Рассмотрение характеристик механорецепторов внутренних органов и сердечно-сосудистой системы, основанное на импульсной активности, регистрируемой в одиночных нервных волокнах, приводит к заключению о том, что нет никаких принципиальных особенностей, отличающих механорецепторы внутренних органов от других тканевых механорецепторов, расположенных в коже, мышцах, суставах и т. д. Все характеристики реакции импульсной активности интероцепторов на действие адекватных механических раздражителей могут быть легко поняты в свете хорошо известных фактов, полученных в опытах на тельцах Пачини, мышечных веретенах и других первичных сенсорных приборах. Эти характеристики определяются особенностями органа и ткани, в которой локализуются рецепторы, развитием и свойствами вспомогательного аппарата, характерисаиками нервных волокон, а также, можно думать, и свойствами собственно рецептирующих элементов.
Кодирование интероцепторами параметров стимула, выражающееся в узоре генерируемой ими импульсной активности, также осуществляется обычным способом. Сохраняются обычные соотношения и между параметрами раздражения и частотой импульсного разряда (например, объемом вдыхаемого воздуха и частотой импульсации тонических рецепторов растяжения легких).
Таким образом, как этого и следовало ожидать, в рисунке разряда механорецепторов внутренних органов и сердечно-сосудистой системы прежде всего четко выступают не столько особенности самих рецепторов, сколько особенности функции тех органов^ которых они локализованы. Только это обстоятельство и определяет присущую им специфику ответов. Ни у каких других механорецепторов не встречается та, по-видимому, длящаяся годами, строго закономерная ритмическая активность, как, например, у рецепторов сердца, сосудов, органов дыхания, обусловленная ритмическим характером работы сердечно-сосудистой и дыхательной систем. Очень хорошо можно выявить и функциональное назначение той информации, которая поступает от интероцепторов (например, таких как ирритантные рецепторы органов дыхания, аортальные барорецепторы и т. д.) в центральную нервную систему.
Импульсная активность опорно-двигательного аппарата
Рецепторы этих структур в свое время были выделены Шеррингтоном (Sherrington, 1906) в особую группу рецепторных приборов — проприоцепторов. Этот термин, наряду с понятиями: экстеро- и интероцепторы, широко используется в литературе, хотя с позиций физиологии рецепторов такое подразделение сенсорных приборов сегодня может показаться и не всегда оправданным. Действительно, у позвоночных в деятельности ряда экстероцепторов (тканевых — в коже), интеро- цепторов и проприоцепторов легко подметить больше сходства, чем различия. Вместе с тем несомненно, что выделение рецепторов двигательного аппарата (мышц, сухожилий, суставов и т. д.) в отдельную группу сенсорных приборов наглядно свидетельствует о той функциональной роли, которую они играют в деятельности организма. Это нашло свое отражение и в интенсивности изучения данных рецепторов (некоторые вопросы классификации рецепторов см. на стр. 471).
Импульсная активность различных проприоцепторов исследовалась весьма подробно. Уже в самых первых работах, выполненных с помощью электрофизиологических методов, анализу активности мышечных нервов было уделено самое пристальное внимание (Adrian, Zotterman, 1926; Adrian, Bronk, 1928, 1929; Matthews, 1929, 1931a, 1931b, 1933). В дальнейшем эти исследования были существенно расширены. Особенно много было проведено их на основном виде проприоцепторов — мышечных веретенах. Они осуществлялись в опытах как на изолированных рецепторах, так и на рецепторах, сохранивших все свои связи с организмом, в экспериментах на теплокровных и холоднокровных животных с применением различных способов воздействия и т. д. Поэтому неудивительно, что в настоящее время физиология проприоцепторов (а мышечных веретен в особенности) является одной из наиболее разработанных глав нейрофизиологии сенсорных систем (обзоры: Granit, 1955, 1970; Глебовский и др., 1969; Ottoson, Shepherd, 1971а; Matthews, 1972; Skoglund, 1973).
Идентификация импульсной активности различных проприоцепторов
В афферентном импульсном разряде мышечного нерва млекопитающих можно различить три основных компонента, которые отражают активность трех основных типов рецепторных образований: первичных и вторичных окончаний веретен, а также сухожильных органов Гольджи. Хотя еще с прошлого столетия на основании работ морфологов было известно о существовании этих рецепторных мышечных приборов, однако лишь сравнительно недавно с помощью различных приемов и прежде всего сопоставления скорости проведения импульсов по нервным волокнам, связанным с этими типами рецепторных структур, удалось окончательно идентифицировать рецепторы, а затем исследовать их функциональные характеристики.
Прежде всего следовало разделить ответы мышечных веретен и сухожильных органов. Как ранее отмечалось ^(стр. 32), вере-
тела включены параллельно экстр афу зальным мышечным волокнам, а сухожильные рецепторы — последовательно. На это обстоятельство впервые обратили внимание в конце 20-х годов (Fulton, Pi-Sufier, 1928). Сразу стало ясно, что благодаря такому расположению рецепторов в ткани лз момент мышечного сокращения нагрузка^на веретена должна уменьшаться (они «разгру-

 А
А
^--1IIIIIHIIIIIIIIIIIHIIIIIIIIIIIII
l-WIIII-HUHII
HI
III111
Рис. 48. Ответы рецепторов мышечных веретен (РМВ) и сухожильных рецепторов Гольджи (СР) при кратковременном стимулирующем толчке (Л) и при медленно нарастающем длительно поддерживаемом растяжении (Б) (А — по: Matthews, 1972; Б — по: Matthews, 1933).
Сверху вниз: механический стимул; биоэлектрическая активность РМВ; то же СР. Отметка времени — 200 мсек.
жаются»), а на сухожильные органы — возрастать (они «нагружаются»), т. е. веретена при этом должны уменьшать свою активность, а сухожильные рецепторы — увеличивать. При растяжении же мышцы оба типа рецепторов должны возбуждаться. Указанный принцип в дальнейшем широко использовался в исследованиях.
Впервые существование предсказанных особенностей ответов мышечных рецепторов было продемонстрировано Мэтьюзом (Mat-
thews, 1933). В его опытах одни рецепторы (A-окончания) прекращали импульсацию при изотоническом (без изменения напряжения, но с уменьшением длины) или изометрическом (без изменения длины, но с увеличением напряжения) сокращении мышцы. Другие же рецепторы (В-окончания) в этих условиях вели себя противоположным образом, т. е. возбуждались. И, наконец, оба типа рецепторов генерировали разряд импульсов при растяжении мышцы. Эти особенности ответов мышечных
Рис.
49. Распределение афферентных волокон
по диаметрам (Z) и скоростям проведения
(II)
для сухожильных органов и мышечных
веретен кошки (I
—
по: Adal, Barker, 1962; II
— по: Hunt, 1954).



I — рецепторы прямой мышцы бедра. А — волокна сухожильных органов; Б и В — соответственно волокна первичных и вторичных окончаний веретен. По оси абсцисс — диаметр волокон (мкм); по оси ординат — отношение числа волокон данного диаметра к общему числу волокон, принятому за 100 (%). Цифры справа от оси ординат — абсолютное число волокон. II — рецепторы камбаловидной мышцы. По оси абсцисс — скорость проведения (м/сек.); по оси ординат — отношение числа волокон с данной скоростью проведения к~ общему числу волокон, принятому за 100 (%). Сплошная линия — афферентные волокна веретен, пунктирная — сухожильные органы.
веретен и сухожильных органов хорошо видны на рис. 48. Было ясно, что A-окончания являются мышечными веретенами (сейчас можно думать, что это были первичные окончания), а В-окончания — сухожильными органами.
Другим подходом к идентификации различных сенсорных структур в мышце явилось изучение диаметров афферентных нервных волокон и скоростей проведения импульсов. Обычно мышечные афферентные волокна делят на четыре группы (I—IV) (Lloyd, 1943). Группу I образуют наиболее толстые афферентные волокна с диаметром от 12 до 20 мкм (максимум в области 15—16 мкм). Группу II составляют волокна с диаметром 4—
Таблица 3
Обычная классификация проприоцептивных афферентных волокон (по: Matthews, 1972)
Группа волокон |
Диаметр (мкм) |
Скорость проведения (м/сек.) |
Иннервируемые рецепторы мышц |
Иннервируемые рецепторы суставов |
I (толстые миелинизированные) |
12-20 |
72—120 |
Первичные окончания веретен (подгруппа 1а) Сухожильные органы (подгруппа lb) Отдельные свободные окончания (?) |
Некоторые органы Гольджи |
II (средние миелинизированные) |
4—12 |
24—72 |
Вторичные окончания веретен Пачиниподобные тельца Некоторые свободные окончания (число очень вариабельно для разных нервов) |
Основная часть органов Гольджи Окончания Руффини Некоторые свободные окончания |
III (тонкие миелинизированные) |
1-4 |
6—24 (может быть, всего лишь 2 м/сек.?) |
Свободные окончания (разных видов ?) |
Пачиниподобные тельца Свободные окон чания |
IV (немиелинизированные) |
меньше 1 |
меньше 2 |
Свободные окончания |
Свободные окончания |
Примечание. Приведенные значения относятся только к задней конечности кошки (и даже в этом случае не являются абсолютно точными). Приведенные диаметры и скорости относятся к волокнам, находящимся в основных нервных стволах. Вблизи рецепторов все величины должны уменьшаться, но не строго закономерно. У моторных нервов альфа-волокна характеризуются скоростью проведения 50—120 м/сек., гамма-волокна — около 10—50 м/сек., что соответствует диаметрам волокон 8—20 и 2—8 мкм соответственно.
12 мкм (максимум 8—9 мкм). Группу III образуют волокна с диаметром 1—4 мкм (максимум около 3—4 мкм), а группу IV (С-во- локна) — немиелинизированные тонкие волокна (ниже 1 мкм) (табл. 3).
Как удалось установить в дальнейшем, волокна первичных и вторичных нервных окончаний веретен различаются между собой по диаметру (рис. 49, I). Это и явилось основой для различения ответов данных рецепторов (Merton, 1953; Hunt, 1954).
После предварительной идентификации веретенных и сухожильных рецепторов удалось на основе рассмотренного принципа показать (Hunt, 1954), что нервные волокна, связанные с веретенами мышц задней конечности, по скорости их проведения могут быть подразделены на две группы (рис. 49, II). Так, например, одна группа волокон, идущая к m. soleus кошки, обладала скоростью проведения от 12 до 60—72 м/сек; другая — от, 60—-72 м/сек. до 114 м/сек. Наконец, волокна, связанные с сухожильными органами, имели лишь один максимум (между 80— 90 м/сек.); пределы же колебаний их скоростей проведения находились в основном между 70—100 м/сек. По мнению автора, раздел между I и II группами нервных волокон лежит в районе 72 м/сек., что соответствует диаметру волокон примерно 12 мкм. Автор высказал мнение, что группу I образуют волокна первичных окончаний, а группу II —волокна вторичных окончаний
А

Рис. 50. Различия в ответах одиночных первичного (Пере.) и вторичного (Втор.) окончаний веретена на быстрое растяжение камбаловидной мышцы у кошки (растяжение приблизительно на 14 мм со скоростью 70 мм/сек.) при наличии (А) и в отсутствие (В) фузимоторной активности (по: Matthews, 1972).
Сверху вниз: напряжение мышцы (достигающее при растяжении 300 г); биоэлектрическая активность вторичного окончания (скорость проведения в волокне — 85 м/сек.); то же первичного окончания (скорость проведения в волокне — 44 м/сек.). Отметка времени — 0.1 сек.
веретен. В настоящее время исследователи располагают целым набором критериев, позволяющих им уверенно идентифицировать различные виды рецепторов мышц. Тем не менее скорость проведения, оцениваемая по латентному периоду, до сих пор является основным методом при различении импульсной активности в волокнах первичных и вторичных окончаний (Granit, 1970).
Первичные и вторичные нервные окончания располагаются в различных участках интрафузальных мышечных волокон, причем механические свойства этих участков существенно разнятся. В части случаев разные окончания располагаются к тому же и на разных типах мышечных волокон (ЯС- и ЯЦ-волокна), а отличия в свойствах вспомогательного аппарата неминуемо должны сказываться и на особенностях ответных реакций рецепторов при действии различных стимулов. Должно также иметь значение и различие в толщине афферентных волокон, отходящих от рецепторов, так как волокна большего диаметра имеют более низкий порог возбуждения. Эти различия действительно были выявлены в экспериментальных условиях.
Как показали исследования ряда авторов, пороги возбуждения первичных рецепторов были ниже, чем вторичных (Hunt, 1954; Lundberg, Winsbury, 1960; Cooper, 1961; Harvey, Matthews, 1961; Bessou, Laporte, 1962; Bianconi, van der Meulen, 1963; Tamaki, 1967). Особенно четко эта разница обнаруживалась при нанесении быстрых раздражений, поскольку первичные окончания обладали наибольшей чувствительностью к действию динамической составляющей стимула (рис. 50). Так, при использовании краткого (менее 5 мсек.) механического стимула показано (Lundberg, Winsbury, 1960), что при небольшом исходном натяжении ш. soleus кошки порог возбуждения волокон группы 1а наблюдается при раздражениях амплитудой менее 60 мкм, а для некоторых волокон — даже при воздействии порядка 10 мкм. Половина же волокон II группы возбуждалась лишь стимулами свыше 500 мкм, а из остальных волокон 1/3 — стимулами порядка 150—200 мкм. Только одно волокно имело порог возбуждения, сходный с таковым самого нечувствительного волокна 1а группы.
Заметно больше, чем для первичных окончаний, был и порог возбуждения сухожильных органов Гольджи. Выявлены различия в порогах возбуждения нервных окончаний и сухожильных органов (Stuart et al., 1970). Однако эти данные отличаются от результатов описанной выше работы (Lundberg, Winsbury, 1960) в отношении порогов возбуждения первичных и вторичных окончаний, что, по-видимому, можно объяснить известными различиями в технике эксперимента. По данным другого исследования (Tamaki, 1967), минимальная скорость растяжения, необходимая для возбуждения первичных окончаний m. triceps surae кошки, равняется 3 мм/сек., вторичных — 29 мм/сек., а сухожильных органов Гольджи — 10 мм/сек. Наличие у веретен интрафузаль- ной мускулатуры, обладающей своей моторной иннервацией, позволяет использовать и это их отличие от сухожильных органов для целей идентификации.
Активность пассивных веретен
Представляется целесообразным рассмотреть вначале свойства мышечных веретен, изолированных от влияний центральной нервной системы. Такие веретена, лишенные эфферентной иннервации, получили наименование пассивных. Поведение пассивных веретен в ряде отношений существенно отличается от реакций активных веретен, сохранивших фузимоторную иннервацию, что говорит о большом значении эфферентной регуляции в нормальной деятельности данного типа механорецепторов. Эфферентная регуляция работы мышечных веретен будет рассмотрена особо в главе 5.
Первичные окончания. Как уже отмечалось, изучение свойств первичных окончаний веретен было начато в работе Мэтьюза (Matthews, 1933). Однако в дальнейшем систематическое исследование их у млекопитающих оказалось в известной мере затрудненным в связи с методическими сложностями, в частности трудностями идентификации ответов различных типов мышечных рецепторов. Поэтому ряд важных сведений о поведении первичных окончаний удалось получить в опытах на лягушках, веретена которых содержат только один тип рецепторных структур и которые представляют собой весьма удобный объект для точных аналитических исследований in vitro (Katz, 1950а, 1950b; Otto- son, Shepherd, 1965, 1969, 1970a, 1970b, 1970c, 1971a, 1971b; Shepherd, Ottoson, 1965; Husmark, Ottoson, 1970, 1971a, 1971b, 1971c, 1971d, и др.).
Первичные окончания часто обладают фоновой активностью даже при полностью расслабленной мышце. Эта активность может быть весьма значительной и нерегулярной у интактных веретен (Matthews, 1933; Matthews, Stein, 1969b, и др.). В значительной степени ее можно отнести за счет деятельности фу зимоторных волокон. После перерезки эфферентных путей она может уменьшаться, становясь при этом более регулярной, и даже полностью блокироваться (рис. 50). Однако, как правило, фоновую импульсацию можно наблюдать и в изолированных рецепторах (Katz, 1950а; Ottoson, 1961), что свидетельствует о периферической природе этого явления, о каких-то процессах, протекающих непосредственно в самом рецепторе. При отведении потенциалов вблизи нервного окончания часто можно наблюдать появление спонтанно возникающих потенциалов действия небольшой амплитуды. По-видимому, они представляют собой потенциалы действия терминальных и претерминальных веточек основного афферентного волокна. Возможно, что несколько таких абортивных спайков могут генерировать распространяющийся нервный импульс (Katz, 1950а).
При растяжении первичного окончания^в момент нанесения раздражения возникает вспышка импульсов (рис. 48). Если растяжение сохраняется, то возросшая первоначально частота импульсации постепенно снижается до некоторого стационарного уровня, величина которого превышает исходное фоновое значение, и тем больше, чем значительнее была растянута мышца. Таким образом, в ответе рецептора четко видны динамический и статический компоненты, Хорошо заметно также в начальный период действия раздражения развитие процесса адаптации, хотя сам рецептор, благодаря его чувствительности к положению, т. е. к стационарной длине мышцы, следует отнести к числу тонических, медленно адаптирующихся структур.
Интенсивность первоначального возрастания импульсации, а также узор разряда зависят как от амплитуды растяжения, так и от скорости его нарастания. Частота статического разряда первичных окончаний у кошки приблизительно пропорциональна показателю растяжения (Eldred et al., 1953).
В тех случаях, когда это удалось выяснить, можно убедиться, что характеристики импульсного ответа мышечных веретен тесно связаны с особенностями протекания РП. Исследование связи величины разряда с амплитудой деполяризационного РП (генераторного потенциала) было начато еще Катцем (Katz, 1950b). Дальнейшие исследования, проведенные также на лягушках (Ottoson, Shepherd, 1971а), подтвердили первоначальные данные и показали, что не только статический, но и динамический компонент реакций пропорционален амплитуде ГП и величине растяжения. При этом динамический ответ достигает максимального значения при более низких величинах растяжения, чем статический. Частота разряда прямо пропорциональна скорости растяжения рецептора (Shepherd, Ottoson, 1965).
Латентное время. При быстром растяжении веретена лягушки латентное время появления первого ПД составляет 0.7—0.8 мсек., а при медленном растяжении оно может достигать 200 мсек. (Ottoson, Shepherd, 1971а). Зависимость скрытого периода ПД от скорости растяжения рецептора носит полулогарифмический характер. Появлению спайка предшествует развитие препотенциала (скорость его нарастания растет при увеличении скорости и величины растяжения — Ottoson, Shepherd, 1970b), который при быстром растяжении имеет скрытый период, равный примерно 0.3—0.4 мсек, (у веретена, которое перед нанесением растяжения находилось в нерастянутом состоянии) или менее 0.3 мсек, (у предварительно несколько растянутого рецептора). Это означает, что минимальный скрытый период возникновения РП в мышечном веретене был примерно таким же, как например у телец Пачини (стр. 79) или волосковых рецепторов беспозвоночных (стр. 100). Такое быстрое развитие реакции означает, по мнению исследователей (Ottoson, Shepherd, 1971а), что РП в веретенах является результатом механической деформации сенсорной мембраны окончания, а не следствием какого-либо промежуточного процесса, например обусловленного выделением химического передатчика. Этот вывод полностью совпадает с мнением и других исследователей, работавших с мышечными веретенами (Katz, 1950b; см. также обзор: Granit, 1970).
Рефрактерные периоды. В опытах на лягушке было установлено, что абсолютный рефрактерный период афферентного нервного волокна веретена равнялся примерно 2 мсек.
(Ottoson, Shepherd, 1969). Относительный рефрактерный период длился 4—5 мсек., период субнормальности — 20—30 мсек., а следовавший затем период несколько повышенной возбудимости — 60 мсек, и более. Ясно, что абсолютный рефрактерный период не определяет ответа веретена лягушки, так как межимпульсный интервал разряда практически всегда больше, остальные же периоды измененной возбудимости могут иметь значение в формировании импульсной последовательности. В явлениях угнетения, отмечаемых после нанесения раздражения на веретено, роль явлений рефрактерности, по-видимому, менее значительна, чем роль следовых эффектов в механических структурах рецептора (Ottoson, Shepherd, 1971а).
Паузы в деятельности веретен. Начиная с первых работ на мышечных веретенах лягушки (Matthews, 1931а, 1931Ь) и кошки (Matthews, 1933), многочисленные авторы показали, что рисунок ответа веретен может значительно варьировать в зависимости от характера наносимого на мышцу раздражения, а также в зависимости от особенностей самих веретен (Katz, 1950а, 1950b; Hunt, Kuffler, 1951а; Granit, van der Meulen, 1962; Jansen, Matthews, 1962a; Jansen, Rudjord, 1963; Renkin, Vallbo, 1964; Shepherd, Ottoson, 1965, и др.).
В частности, этими исследователями отмечалось существование пауз в активности веретен. Пауза, обусловленная разгрузкой веретена в момент сокращения мышцы, наблюдается в случае как первичных, так и вторичных нервных окончаний. У интактных веретен имеет место заполнение паузы за счет активности эфферентных нервных волокон, обусловливающих сокращение интрафузальной мускулатуры и, соответственно, возбуждение рецепторов (рис. 74).
Перед возникновением паузы часто] ^отмечается увеличение частоты разряда (у первичных окончаний это наблюдается чаще, а у вторичных — реже) (Hunt, 1954). После этой первой паузы в части случаев появляется вспышка импульсов,, за которой следует новая пауза, и лишь затем импульсация рецептора возвращается к первоначальному уровню. В других же случаях сразу после паузы возникает стационарный разряд, который может иметь весьма регулярный характер (Granit, van der Meulen, 1962). В зависимости от генерируемого веретенами разряда авторы делят обследованные рецепторы на фазные и тонические. Вторая вспышка разрядов и вторая пауза наблюдались в ответах именно фазных веретен. Возможно, что тоническим ответом обладали первичные окончания ЯЦ-волокон. Рисунок разряда, кроме того, существенно зависел от величины и скорости сокращения мышцы, а также от исходного напряжения.
В той же работе (Granit, van der Meulen, 1962) было выявлено существование двух типов рецепторов. Одни обладали короткой паузой (короткопаузные), а вторые — длинной (длиннопаузные). В короткопаузных веретенах m. gastrocnemius наблюдалась пауза
при сокращении мышцы после раздражения ее нерва 2—3 ударами тока, равная в среднем 70 мсек., а в веретенах m. soleus она составляла 150 мсек. У длиннопаузных веретен этих мышц величина паузы равнялась 180 и 310 мсек, соответственно.
Начальная вспышка разрядов могла быть обусловлена рядом причин. Во-первых, наличием у веретена альфа-(бета)-иннерва- ции, благодаря которой интрафузальные волокна возбуждаются одновременно с экстрафузальными. Во-вторых, она могла быть связана с появлением возвратного ответа на основе эфаптиче- ского раздражения веретена высокосинхронными ПД или потенциалом концевых пластинок (Lloyd, 1942; Leksell, 1945; Hunt, Kuffler, 1951a, 1951b; Granit et al., 1959a, 1959b; Brown, Matthews, 1960). В-третьих, она может быть вызвана растяжением веретена экстрафузальными волокнами, расположенными асимметрично ему (Hunt, Kuffler, 1951а, 1951b; Granit et al., 1959a, 1959b; Green, Kellerth, 1967). Возможно, что начальная вспышка могла быть обусловлена одновременным действием нескольких факторов (Granit, 1970).
Возникновение небольшой паузы в активности веретена отмечается и при резком растяжении мышцы вслед за начальным разрядом (Matthews, 1931а). Данное наблюдение в дальнейшем было подтверждено как на лягушках (Katz, 1950b; Shepherd, Ottoson, 1965), так и на млекопитающих (Jansen, Matthews, 1962а; Jansen, Rudjord, 1963). По наблюдениям Катца, эта пауза делит ответ на две части — динамическую и статическую — и является отражением изменений локального, рецепторного потенциала в момент перехода от динамической фазы к статической. В дальнейшем эта краткая пауза, или переходный интервал (transitional interval), детально изучалась на изолированном веретене лягушки (Shepherd, Ottoson, 1965).
Опыты показали, что данная пауза является хорошо воспроизводимым феноменом и ее длительность находится в тесной связи с амплитудой и скоростью нарастания стимула (рис. 51). Пауза наблюдается именно в момент завершения динамической фазы ответа, между динамическим и статическим компонентами реакции, и лишь тогда, когда амплитуда и (или) скорость раздражения достаточно высоки. При скоростях растяжения ниже 20 мм/сек. она не возникает даже при очень больших амплитудах растяжения. При увеличении скорости раздражения величина паузы возрастает, возрастает и частота разряда в момент завершения динамического компонента реакции, а начальная частота статического компонента ответа практически не меняется. Однако такое положение наблюдается до определенного предела. При перерастяжении пауза исчезает, а динамический и статический разряды сливаются. Пауза также отсутствует при слишком малых силах воздействия.
Данная пауза, по-видимому, может иметь определенное информационное значение, так как обостряет импульсационный 160
контраст и позволяет центрам лучше судить о динамических и статических характеристиках стимула (Shepherd, Ottoson, 1965). Аналогичного типа пауза может отмечаться и в реакции некоторых других рецепторов, например фоторецепторов глаза мечехвоста (Dowling, 1968), волосковых механорецепторов насекомых (Thurm, 1965b), что позволяет говорить о существовании ряда общих свойств в деятельности различных медленно адаптирующихся рецепторов, и в частности их РП (Ottoson, Shepherd, 1971а).
Рис.
51. Ответы изолированного мышечного
веретена лягушки на динамическую и
статическую составляющие раздражения
(по: Shepherd, Ottoson, 1965).




Сверху вниз: биоэлектрическая активность рецептора; отметка раздражения (растяжение возрастающей амплитуды и скорости). Скорость растяжения — 130 (1—3), 13 (4—6), 1.3 мм/сек. (7). Амплитуда растяжения одинакова на 1 и 4, 2 и <5, а также на 3, 6 и 7. Видна пауза между динамическими и статическими разрядами при больших скоростях и амплитудах растяжения. Отметка времени — 50 мсек.
Как и любое другое явление, протекающее в рецепторах и оцениваемое по характеру импульсной активности, переходный интервал может теоретически иметь в своей основе по крайней мере три причины: свойства вспомогательных структур (прежде всего вязко-упругие особенности интрафузальных волокон), свойства нерегенеративной системы генерации (особенности РП) и, наконец, свойства регенеративной системы генерации потенциалов действия. Вторая причина, по-видимому, является основной (Ottoson, Shepherd, 1971а).
Уже ранние исследования (Katz, 1950b) показали, что возникновение паузы происходит в момент завершения динамической фазы действия стимула, и именно в этот момент происходят существенные изменения в картине развития РП: величина динамического пика РП, достигнув максимума, начинает уменьшаться,
161
и наблюдается постепенный переход к статической платообразной части РП (рис. 52). Такой переход от динамической к статической части является весьма характерной особенностью РП медленно адаптирующихся рецепторов вне зависимости от их функциональной специализации. Эта особенность отмечалась в РП фоторецепторов (Fourtes, Poggio, 1963; Fulpius, Baumann, 1969; Mil- lecchia, Mauro, 1969a, 1969b), хеморецепторов (Boeckh et al., 1965), волосковых механорецепторов (Smola, 1970b) и др., что


Рис. 52. Соотношение между рецепторным потенциалом и импульсной активностью мышечного веретена у лягушки (по: Ottoson, Shepherd, 1970с).
Динамический разряд в ответ на короткое нарастающее (а—в) и длительное медленное (г) астяжение. ПД зарегистрированы до блокирования 0.2%-м раствором лигнокаина, после чего регистрируются только РП (записи с наложением). Нижний луч — запись растяжения. Отметка времени: на а—в — 25 мсек.; на г — 100 мсек.
позволяет говорить об известной общности природы этого феномена.
Вторичные окончания. Эти рецепторы изучены значительно меньше, нежели первичные окончания. Некоторые их свойства уже отмечались выше. Основная особенность ответов вторичных окончаний состоит в том, что в них почти отсутствует динамический компонент. Это существенно отличает вторичные рецепторы от первичных (рис. 50). После снятия нагрузки в активности вторичцых окончаний не отмечается паузы, характерной для первичных окончаний (рис. 53).
Изучение межспайковых интервалов показало, что импульса- ция от вторичных окончаний обладает большей стабильностью, чем в случае первичных окончаний. Особенно это было заметно у активных (сохранивших эфферентную регуляцию) веретен (см. рис. 73).
Вибрационная чувствительность веретен. Различный характер ответов первичных и вторичных окончаний на действие динамической составляющей раздражения получает свое отражение и в их реакции на вибрационные стимулы. Оказалось, что им-
Рис.
53. Различия в ответах первичных и
вторичных окончаний веретен при
медленном растяжении деэфферентированной
камбаловидной мышцы у кошки (по: Harvey,
Matthews, 1961, отретушировано).
А — первичное окончание со сравнительно тонким афферентным волокном (скорость проведения — 83 м/сек.); Б — вторичное окончание со сравнительно толстым афферентным волокном (скорость проведения — 65 м/сек.). Скорость растяжения — 2.3 мм/сек. На Б исключен участок, соответствующий периоду в 1 сек. Сверху вниз: растяжение мышцы; импульсная активность. Стрелки — начало и конец статической фазы воздействия. Отметка времени — 1 сек.
Пер
6.
Втор.
с:
О
75
мкм

![]()
![]()


Рис. 54. Различное влияние вибрации на первичное (Пере.) и вторичное (Втор.) окончания, расположенные в одной и той же части деэфферентированной камбаловидной мышцы (по: Matthews, 1967).
Прямые линии — продольная вибрация (частота — 300 гц); цифры над ними — амплитуда раздражения. Шкала — частота импульсов в ответах.
пульсная активность первичных окончаний обладает очень высокой чувствительностью к вибрациям (рис. 54) (Kuffler et al., 1951; Granit, Henatsch, 1956; Bianconi, van der Meulen, 1963; Crowe, Matthews, 1964b; Brown et al., 1967; Takano, Homma, 1968; Арутюнян, 1971, и др.). Так, рецепторы могли возбуждаться вибрациями, приложенными к сухожилию, при амплитуде вибраций менее 5 мкм (Brown et al., 1967). Фактическая же величина смещений в области самого веретена была в этом случае менее 1 мкм. Первичные окончания могли генерировать импульсы в ритме раздражения вплоть до частоты в 200—400 гц. Растяжение мышцы повышало чувствительность первичных окончаний к действию вибрации, а также их способность воспроизводить ритм раздражения.
Вторичные окончания обладают невысокой чувствительностью к действию вибрационных раздражителей, приложенных к сухожилию мышцы (рис. 54), и неспособны следовать за ритмом стимуляции, особенно при небольших амплитудах воздействия, даже на низких частотах (Besson, Laporte, 1962; Bianconi, van der Meu- len, 1963; Brown et al., 1967; Matthews, 1967).
Раздражение веретена путем растяжения, приложенного к сухожилию, является, по-видимому, более адекватным воздействием. Если же конец вибратора помещали на мышцу вблизи вторичных окончаний, то можно было в части случаев наблюдать их ответы на вибрационные стимулы. Первичные же рецепторы хорошо возбуждались вибрацией при различных способах ее подведения (Bianconi, van der Meulen, 1963). Такое различие в реакции вторичных нервных окончаний на действие вибрации в зависимости от пути, по которому она их достигает, лишний раз свидетельствует о важной роли механических свойств вспомогательных структур рецепторов, а также и окружающих тканей в формировании ответов механорецепторных приборов на действие раздражителя.
Реакция на скорость и положение. При исследовании импульсации от первичного окончания в ответ на растяжение было установлено, что в самом начале раздражения имеется резкое возрастание частоты, за которым обычно после снижения активности наблюдается новое более медленное усиление импульсации (Jansen, Matthews, 1962а; Matthews, 1963; Lennerstrand, Thoden, 1968a). Чем больше скорость растяжения, тем более выражено первоначальное увеличение частоты разряда. Анализ этого явления привел исследователей к заключению, что начальную величину можно объяснить действием сухого трения в момент начала движения (своего рода эффект «заедания») и, так как оно отражает лишь переходные процессы, его нельзя использовать при изучении основных характеристик мышечных веретен.
Было показано (Lennerstrand, Thoden, 1968а, 1968b), что веретена не реагируют на действие ускорения, а отвечают лишь на скорость растяжения и на смещение. При этом если из ответа веретена вычесть статическую реакцию, обусловленную положением, т. е. получить чисто динамическую часть ответа, то в этой части можно выявить два компонента — быстрый и медленный. Оказалось, что расчленить реакцию рецепторов на эффект сухого трения, статическую часть и динамическую часть с быстрым Щмёд-
Таблица 4
Сравнение двух типов афферентных окончаний веретен кошки (по: Matthews, 1972)
Сравниваемые
признаки
Первичное
окончание
Вторичное
окончание


Локализация
Афферентное
волокно
Эфферентный
контроль
Ответ
на действие линейного растяжения
величиной в несколько миллиметров:
в
начале раздражения
Средне-экваториальная
область ЯС- и ЯЦ-во- локна
Большое,
группа !а (диаметр 12—20 мкм, скорость
проведения 70— 120 м/сек.)
Как
статические, так и динамические гамма-
волокна
Высокая
динамическая чувствительность
Может
наблюдаться быстрое возрастание
частоты импульсации
после
завершения динамического растяжения
при
расслаблении
Ответ
на удар по сухожилию
Ответ
на вибрацию, приложенную к сухожилию
Вариабельность
меж- спайкового интервала
Чувствительность
к малым смещениям
Большой
динамический индекс (40—120 имп./ сек. при
растяжении деэфферентированного
препарата со скоростью 10 мм/сек.)
Резкое
прекращение разряда (при условии
незначительной динамической
фузимоторной активности)
Низкий
порог (50 мкм); возможен множественный
разряд
Низкий
порог (50 мкм) для воспроизведения ритма
при частотах 100—300 гц
Высокая
(средний коэффициент вариации 6% при
деэфферентации, 25% при децеребрации)
Высокая
(типичная —100 имп.-сек."1-мм-1
при децеребрации)
Рядом
с экваториальной областью (преобладают
ЯЦ-волокна)
Среднее,
группа II (диаметр 4—12 мкм, скорость
проведения 20— 70 м/сек.)
Статические
гамма-волокна
Низкая
динамическая чувствительность
Постепенное
увеличение разряда, линейно связанное
с прогрессивно нарастающим растяжением
Малый
динамический индекс (5—21 имп./сек.
при тех же условиях)
Постепенное
снижение импульсации до нуля
Высокий
порог (500 мкм)
Высокий
порог (по крайней мере 250 мкм для 100
гц)
Низкая
(средний коэффициент вариации 2% при
деэфферентации, 6% при децеребрации)
Низкая
(типичная —
7
имп. - сек.-1
• мм-1
при децеребрации)










Примечание. Помимо указанных различий первичные и вторичные окончания мышечных веретен обладают чертами сходства: 1) ультраструктурой собственно нервных окончаний и 2) статическим ответом при действии больших растяжений (чувствительность — около 4 имп.«сек.-1*мм-1 для обоих видов окончаний).
ленным компонентами было легче всего в опытах с применением треугольных стимулов (Lennerstrand, Thoden, 1968а).
Линейное
растяжение Статуя//"
Пер&ичное llllllllllllllllllllllll
Вторичное
шшшш
Синусоидальное
Удар
растяжение Освобождение
![]()

пни hi in iiiii uni iii шш ш
Ш1Ш lllllllllllll llllllil 11 Illi I
Рис. 55. Схема типичных ответов первичного и вторичного окончаний веретен на действие различных стимулов большой амплитуды в отсутствие фузи- моторной активности (по: Matthews, 1964).
В реакции вторичных окончаний можно было выделить, как и в ответе первичных окончаний, два компонента динамического разряда, однако они были выражены значительно слабее. Это является отражением функциональных свойств вторичных окончаний, предназначенных в первую очередь для измерения длины (смещения, положения).
Особенности реакций первичных и вторичных окончаний мышечных веретен на действие различных стимулов суммированы на рис. 55 и в табл. 4.
Активность сухожильных органов Гольджи
Деятельность этих рецепторов изучалась во многих работах (Matthews, 1933; Jansen, Rudjord, 1964; Alnees, 1967; Houk, 1967; Houk, Henneman, 1967; Houk, Simon, 1967; Stuart et aL, 1970; Houk et al., 1971, и др.), хотя и менее интенсивно, чем мышечные веретена. При этом исследовалась только импульсная активность рецепторов, которая на основании ряда критериев могла быть весьма четко отдифференцирована от активности мышечных веретен. Правда, такого рода разделение в некоторых случаях сталкивалось с известными трудностями (например, при существовании альфа-фузимоторной иннервации веретен, атипичной реакции веретен при мышечном сокращении и т, д ).
Выявление признаков, отличающих импульсную реакцию сухожильных органов от импульсного ответа мышечных веретен, было начато еще в первой работе Мэтьюза (Matthews, 1933), в которой удалось различить ответы: A-окончаний (веретен), В-окончаний (сухожильных органов) и С-окончаний (свободных нервных окончаний). Как уже отмечалось, в силу своей локализации сухожильные органы включены последовательно экстра- фузальным мышечным волокнам, и поэтому они возбуждаются в момент мышечного сокращения, т. е. в момент, когда мышечные веретена разгружаются и их импульсация исчезает (период молчания). Такое различное поведение сухожильных органов и мышечных веретен хорошо видно на рис. 48.
Сухожильные органы в своем составе не содержат мышечных структур. Поэтому с помощью веществ, активирующих интра- фузальные мышцы и, соответственно, возбуждающих веретена, но не действующих на сухожильные органы, также можно отличить активность одного типа рецепторов от другого (это было показано, например, в опытах с использованием сукцинилхо- лина — Granit et al., 1953). Естественно также, что раздражение гамма-волокон сказывается лишь на веретенах, но не на сухожильных органах (Hunt, Kuffler, 1951а).
При растяжении мышцы сухожильные органы могут быть возбуждены, как и веретена, однако порог их активации оказался значительно больше (Matthews, 1933). Мэтьюз обнаружил также отличие в реакции веретен и сухожильных органов при действии быстрого растяжения. В момент действия динамической фазы стимула мышечное веретено разряжается сО значительно большей частотой, чем после ее завершения и достижения постоянного уровня растяжения. Сухожильные же органы такой особенностью не обладали. Благодаря своей низкой чувствительности к скорости изменения растяжения, частота их разряда в момент действия динамической фазы раздражения лишь незначительно превышала частоту при стационарной стимуляции (рис. 48). В дальнейшем/ правда, оказалось, что чувствительность к скорости растяжения присуща лишь первичным окончаниям веретен, а вторичные окончания в этом отношении ведут себя аналогично сухожильным органам. В свою очередь чувствительность последних к быстрому растяжению и особенно к действию нарастающего мышечного сокращения зависит от места их залегания (Alnees, 1967). Например, чувствительность к скорости воздействия оказалась у рецепторов передней мышцы голени значительно выше, чем у рецепторов камбаловидной головки икроножной мышцы. Пороговые скорости нарастания растяжения, необходимые для получения возбуждения, были у сухожильных органов выше, чем у первичных окончаний, но ниже, чем у вторичных (Tamaki, 1967).
Уже в ЗО-е годы Мэтьюз (Matthews, 1933) показал, что сухожильные органы являются медленно адаптирующимися рецепторами. По данным этого автора, различий в реакциях рецепторов в зависимости от характера раздражения (пассивного растяжения или активного сокращения) не наблюдалось. В итоге Мэтьюз пришел к заключению, что сухожильные органы являются датчиками, регистрирующими напряжение, различным образом возникающее в мышце. Аналогичные данные получил и Альнес (Alnaes, 1967), который обнаружил, что при одинаковом напряжении в сухожильном органе Гольджи частота его разряда оказывается одной и той же вне зависимости от того, каким способом это напряжение было достигнуто. Вместе с тем следует указать (Jansen, Rudjord, 1964; Houk, Henneman, 1967), что активное мышечное сокращение является более эффективным стимулом для сухожильных органов. При таком воздействии порог раздражения был ниже, чем при пассивном растяжении. Пассивное растяжение, даже превышающее физиологические границы, по данным этих авторов, редко может вызвать появление устойчивого разряда сухожильного органа. Однако отмечается существование стационарной активности у 2/3 сухожильных органов ш. soleus при очень сильной дорзофлексии (Stuart et al., 1970). Очевидно, что большое значение имеет как выбор мышцы для исследования, так и применяемая для ее раздражения методика воздействия. Поэтому, по мнению Гранита (Granit, 1970), нет оснований для отказа от первоначального представления Мэтьюза о сухожильных органах как датчиках напряжения во всех случаях воздействия на мышцу.
Недавно было проведено (Houk et al., 1971) систематическое обследование характеристик сухожильных органов мышц (mm. soleus, gastrocnemius med., tibialis ant.) кошки в условиях, приближающихся к нормальным. Эти авторы установили, что лишь 35% всех рецепторов обладают стационарной активностью при растяжении, вызванном вращением в суставе в физиологических пределах. При этом подавляющее большинство указанных рецепторов активировалось лишь на последних 10° вращения. По данным этих авторов, сухожильные рецепторы возбуждаются в в 1.5—8 раз более эффективно при мышечном сокращении, нежели при пассивном растяжении, если наносимое раздражение находится в пределах физиологической нормы. Наконец, в экспериментах на мезенцефалических животных было показано (Северин и др., 1967), что при ходьбе сухожильные органы генерируют наибольшую активность при сокращении их собственных мышц.
Важные сведения о работе сухожильных органов были получены в экспериментах с раздражением одиночного моторного волокна в вентральном корешке (Houk, Henneman, 1967). Было показано, что сокращение одиночной моторной единицы достаточно для возбуждения сухожильного органа Гольджи (рис. 56). Оказалось, что на один орган Гольджи могут влиять от 4 до 15 различных двигательных волокон, каждое из которых, естественно,
иннервирует ряд мышечных волокон, оканчивающихся на различных сухожильных рецепторах. По гистологическим данным, с одиночными сухожильными рецепторами связано от 3 до 25 мышечных волокон. Это означает, что одного, самое большее — двух мышечных волокон достаточно для приведения сухожиль-
 Рис.
56. Ответ сухожильного органа Гольджи
камбаловидной мышцы на тетаническое
сокращение одиночной моторной
единицы, вызванное раздражением
одиночного альфа- волокна в вентральном
корешке (по: Houk, Henneman, 1967).
Рис.
56. Ответ сухожильного органа Гольджи
камбаловидной мышцы на тетаническое
сокращение одиночной моторной
единицы, вызванное раздражением
одиночного альфа- волокна в вентральном
корешке (по: Houk, Henneman, 1967).
Сверху вниз: напряжение мышцы; афферентный разряд. Сокращение сопровождается напряжением в 18 г и длится около 1 сек.
кого органа в состояние возбуждения. Одновременная активация двух моторных единиц, каждая из которых возбуждала сухожильный орган, приводила к эффекту, большему по величине, чем давала каждая моторная единица в отдельности, но меньше суммы их индивидуальных ответов.
Активность суставных механорецепторов
Рецепторы суставов, по-видимому, сообщают в центральную нервную систему основные сведения о различных параметрах движения (см. обзор: Skoglund, 1973). В последнее время развивается иная точка зрения (Burgess, 1974), согласно которой о перемещениях в суставах сигнализируют рецепторы, расположенные в мышцах. Это разногласие хорошо показывает, что суставные рецепторы исследованы еще сравнительно неполно. Кроме того, из-за технических сложностей, возникающих при их изучении, исследователи до настоящего времени пользовались только одним показателем — импульсной активностью.
Основная масса работ была выполнена на коленном суставе кошки (Gardner, 1948; Andrew, Dodt, 1953; Boyd, Roberts, 1953; Andrew, 1954; Boyd, 1954; Cohen, 1955; Skoglund, 1956; Eklund, Skoglund, 1960; Burgess, Clark, 1969b), и лишь отдельные исследования были осуществлены на других суставах (Andrew, 1954; Godwin-Austen, 1969; Залкинд, 1971, 1972). Основными критериями, на основании которых суставные механорецепторы были подразделены на несколько видов, явились прежде всего адаптационные характеристики их импульсной активности, а также особенности импульсной реакции на амплитуду, скорость и направление движения в суставе.
В суставах, вероятно, преобладают тонические, медленно адаптирующиеся рецепторы. Так, по данным, полученным в детальном исследовании, из 278 обследованных рецепторов 209 обладали именно такого рода особенностями (Burgess, Clark, 1969b). Медленно адаптирующиеся рецепторы могли сохранять частоту своих разрядов практически без изменения в течение 4—5 час. (Skoglund, 1956).
Уже первые исследования суставных механорецепторов (Andrew, Dodt, 1953; Boyd, Roberts, 1953; Skoglund, 1956) показали, что медленно адаптирующиеся рецепторные элементы возбуждаются лишь при определенных й различных для разных рецепторов положениях сустава. Диапазон углов смещений сегментов конечностей, при которых эти рецепторы возбуждались, был весьма невелик (15—30°), а максимум активности наблюдался всего лишь в диапазоне нескольких градусов (рис. 57). Большинство рецепторов активировалось в положениях, близких к полному сгибанию или разгибанию. На основании собственных экспериментов и данных литературы, Скоглунд (Skoglund, 1956, 1973) пришел к заключению, что все суставные рецепторы можно подразделить по крайней мере на три основных типа: два типа медленно и один тип быстро адаптирующихся.
Медленно адаптирующимися рецепторами первого типа являются сухожильные органы Гольджи, лежащие в связках суставов. На их активности не сказывается работа мышц, подход дящих к суставу, и это означает, что они могут сигнализировать о точном положении сустава, т. е. являются позными рецепторами. Рецепторы этого типа могли также сигнализировать о направлении движения (чаще в одном направлении). Однако чувствительность их к движению является низкой, и поэтому они не могут давать информацию о скорости перемещений.
Медленно адаптирующиеся рецепторы второго типа находятся в капсуле сустава, и к ним относят тельца Руффини. Они воспринимают направление движения (встречаются одно- и двунаправленные рецепторы), а их высокая чувствительность к перемещениям позволяет им оценивать и скорость движения (рис. 58). Импульсация этих рецепторов регистрируется в небольшом диапазоне углов. Поскольку на импульсной активности сказывается работа мышц, изменяющих натяжение капсулы сустава, они не могут рассматриваться, как истинные позные рецепторы. Вместе с тем' благодаря своей чувствительности к влиянию мускулатуры эти медленно адаптирующиеся рецепторы могут участвовать в различении активного и пассивного движений.
Быстро адаптирующимися рецепторами в суставах являются пачиниподобные тельца. Они чувствительны ко всем движениям в суставах вне зависимости от начального положения. Частота их разряда зависит от скорости движения. Ответ их состоит из небольшого числа импульсов.
Поведение всех видов суставных рецепторов не выходит за рамки рассмотренных выше свойств различных механорецеп- торных приборов и может быть легко понято на основе общих
имп./сек.
50 г

Рис. 57. «Угловая специфичность» медленно адаптирующихся рецепторов коленного сустава у кошки (по: Skoglund, 1956).
По оси абсцисс — положение голени (градусы сгибания коленного сустава); по оси ординат — частота разрядов в одиночных волокнах (ими./сек.). Соединены точки, характеризующие ответ одного и того же волокна. На графике представлены ответы 5 волокон одного животного (сплошные линии) и 3 волокон другого (прерывистые линии).

А _ ответы на угловое смещение на 5° из одного и того же начального положения в одно и то же конечное при трех различных скоростях смещения. Б — то же в противоположном направлении. 1 — скорость смещения 8°, 2 — 15°, 3 — 30°, 4 — 35°, 5 — 12°, 6 — 5° в 1 сек. По оси абсцисс — время после поворота (сек.); по оси ординат — частота разрядов (имп./сек.).
свойств быстро и медленно адаптирующихся рецепторов. Ясно, что в рисунке ответов медленно адаптирующихся рецепторов большую роль играют свойства окружающих тканей. Наличие же ряда импульсов в быстро адаптирующихся ответах пачиниподоб- ных телец говорит скорее всего о возникновении в тканях при движениях колебательных явлений.
Активность свободных нервных окончаний
Изучение ответов свободных нервных окончаний в мышцах, как отмечалось, было начато Мэтьюзом (Matthews, 1933). Он на основании изучения импульсной активности мышечного нерва описал рецепторы типа С, идентифицированные им как свободные нервные окончания мышечной фасции. Они встречаются относительно редко, генерируют разряд из 5—20 импульсов в период динамической фазы мышечного растяжения, а иногда и при снятии раздражения, но, будучи быстро адаптирующимися структурами, не обладают способностью поддерживать активность в период действия постоянного растяжения.
В дальнейшем свободные нервные окончания в мышцах изучались в отдельных работах (Bessou, Laporte, 1958, 1961; Iggo, 1960; Paintai, 1960). Морфологические исследования показывают, что свободные нервные окончания образуются, как правило, не- миелинизированными, а также тонкими мякотными волокнами. Лишь некоторые свободные окончания являются рецепторами миелинизированных волокон среднего диаметра (табл. 3).
Исследование свободных нервных окончаний, образованных немиелинизированными волокнами (скорость проведения 0.4— 1.6 м/сек.) показало (Bessou, Laporte, 1968; Iggo, 1960), что данные рецепторы, по-видимому, не являются механорецепторами, а скорее служат для восприятия болевых стимулов (они не возбуждались при мышечном сокращении, но активировались при инъекции гипертонического раствора, ишемии, изменениях температуры выше 41° или ниже 25°С). Изучение свойств рецепторов, образованных тонкими мякотными волокнами (скорость проведения ниже 24 м/сек.), показало (Paintai, 1960; Bessou, Laporte, 1961), что некоторые из них могут рассматриваться как вторичные окончания веретен, а отдельные — как сухожильные органы Гольджи. Примерно 15% волокон не возбуждалось механическими стимулами. Большинство же волокон, которые можно было считать оканчивающимися свободными нервными окончаниями, могли возбуждаться при локальном сдавливании поверхности мышцы, но не приходили в состояние значительного возбуждения при мышечном сокращении или растяжении. Пороги таких окончаний к механическим стимулам были значительно выше, чем у кожных рецепторов прикосновения или первичных окончаний веретен. Большинство рецепторов являлось быстро адаптирующимися структурами, лишь некоторые из них адаптировались к действию постоянной нагрузки очень медленно. По одним данным (Bessou, Laporte, 1961), пиковая частота разрядов рецепторов не превышала 30 в 1 сек., а в других исследованиях (Paintai, i960) были обнаружены отдельные волокна с пиковой кратковременной частотой активности, равной 100—150 имп./сек. В отдельных случаях рецепторное поле афферентного волокна могло захватывать всю мышцу; чаще же оно ограничивалось областью всего лишь в несколько квадратных миллиметров.
Если учесть данные Пайнтала (Paintai, 1962), что большинство тонких мякотных волокон не возбуждается при быстром сильном растяжении мышцы или ее максимальном сокращении, при действии вибрации, т. е. при обычных воздействиях, активирующих классические проприоцепторы, можно предположить, что окончания этих волокон вряд ли сигнализируют о механическом состоянии мышцы в ходе ее нормальной работы, но, по-видимому, могут давать информацию о давлении на мышцу.
* * *
Имеющиеся в литературе данные позволяют утверждать, что все рецепторы опорно-двигательного аппарата могут рассматриваться как первичные механорецепторные приборы. Особенности узора генерируемых ими разрядов определяются прежде всего их расположением в тканях, свойствами вспомогательных структур, свойствами их афферентных волокон. В основе импульсного разряда данных рецепторных приборов (как’это показано на примере мышечных веретен) лежат локальные деполяризационные явления (генераторный потенциал), которые развиваются в окончаниях афферентных волокон.
ИМПУЛЬСНАЯ АКТИВНОСТЬ МЕХАНОРЕЦЕПТОРОВ ОРГАНОВ ЧУВСТВ ПОЗВОНОЧНЫХ
Импульсная активность органов боковой линии
Импульсная активность механорецепторов «обычных» органов боковой линии исследуется весьма интенсивно (Hoagland, 1933а, 1933b, 1933с, 1933d; Sand, 1937; Katsuki et al., 1951a, 1951b, 1951c; Katsuki, Yoshino, 1952; Dijkgraaf, 1956; Murray, 1956; Gorner, 1963; Harris, Milne, 1966; Harris, Flock, 1967; Suckling, 1967; Russell, 1968; Roberts, 1972; Alnaes, 1973a, 1973b; Katsuki, 1973; Murray, Capranica, 1973; обзор: Dijkgraaf, 1963; см. также труды симпозиума: Cahn (ed.), 1967).X Рецепторы боковой линии обладают фоновой активностью в состоянии покоя (до 100 ими./сек.), что, по-видимому, обусловлено спонтанным выделением медиатора из волосковых рецептирующих клеток (см. стр. 238). В некоторых опытах более толстые афферентные волокна (максимум диаметров 12 мкм) обладали и более регулярной спонтанной активностью, чем тонкие волокна (максимум диаметров 3 мкм) (Alnees, 1973а). Спонтанная активность механорецепторов боковой линии, говорящая о высокой возбудимости этих приборов, коррелирует с их высокой чувствительностью. Величины пороговых смещений оцениваются порядком 25 А. при стимуляции в диапазоне наиболее хорошо воспринимаемых частот — 75/сек. (Kuiper, 1967). При раздражении рецепторов импульсы группируются в пачки. С усилением воздействия число импульсов в пачке возрастает, а латентное время первого импульса сокращается. Антидромное раздражение угнетает спонтанную активность (Harris, Flock, 1967; Alnaes, 1973b).
Очень характерным в картине импульсного ответа является то, что часть рецепторов возбуждается при одном направлении воздействия, а другие — при изменении направления стимула на 180°. Торможение активности наблюдается при направлении воздействия, противоположном возбуждающему. Такой характер реагирования отмечался в ответах как невромастов каналов боковой линии (Sand, 1937), так и поверхностно расположенных невромастов (Gorner, 1963). Дирекционная чувствительность рецепторов боковой линии была продемонстрирована и в опытах на акулах во время их свободного плавания (Roberts, 1972). Такая чувствительность волосковых механорецепторов к направлению воздействия обусловлена особенностями их строения (см. главы 1 и 10), а также различной ориентацией в органах боковой линии.
Импульсная активность сенсорных элементов вестибулярной системы
Изучение активности афферентных волокон лабиринта у высших позвоночных представляет значительные трудности. Поэтому наиболее подробные исследования выполнялись прежде всего на рыбах (скаты) и земноводных (лягушки) (Ashcroft, Hallpike, 1934; Ross, 1936; Lowenstein, Sand, 1940a, 1940b; Lowenstein, Roberts, 1949; Groen et al., 1952; Groen, 1961a, 1961b; Gualtierotti, Gerathewohl, 1965; Precht et al., 1971; Precht, Llinas, 1972). Впрочем, имеется мнение (см., например, Gernandt, 1959), что нет принципиальной разницы в поведении рецепторных элементов вестибулярной системы у высших и низших позвоночных. Можно отметить, что определенная часть сведений о рецепторах лабиринта может быть почерпнута из экспериментов на втором сенсорном нейроне, т. е. на нейронах вестибулярных, а при изу
чении рецепторов органа слуха — соответственно на нейронах кохлеарных ядер продолговатого мозга [см., например, эксперименты на кошках (Adrian, 1943); обзор этих исследований дан в работе Кислякова и др. (1972)].
В последние годы начали появляться работы, выполненные и на вестибулярных нервных волокнах млекопитающих (Werner, Whitsei, 1968 — цит. по: Tamar, 1972; Fernandez, Goldberg, 1971; Goldberg, Fernandez, 1971; Vidal et al., 1971; Fernandez et al., 1972; Loe et al., 1973, и др.).
Обычно в афферентных волокнах наблюдается спонтанная активность. У млекопитающих она может быть иногда очень высокой (у обезьян до 200имп./сек., в среднем — 90 имп./сек.). В одних волокнах она регулярна, а в других — нерегулярна. Спонтанная активность может изменяться под влиянием раздражения как в сторону увеличения, так и уменьшения (Ross, 1936; Lowenstein, Sand, 1940а, 1940b; Groen et al., 1952; Groen, 1961b, и др.). После возбуждения наблюдается период угнетения активности, а после торможения — усиление импульсации. Следовательно, с помощью рецепторов вестибулярной системы (как и с помощью рецепторов системы органов боковой линии) нервная система может оценивать действие как возбуждающих, так и тормозящих стимулов. Различное действие стимулов зависит от направления раздражения (например, тока жидкости в полукружных каналах) и от ориентации сенсорного эпителия. В основе этой дирекционной чувствительности (подробнее см. главу 10), как и в случае других волосковых сенсорных клеток иных органов чувств позвоночных, лежит морфологическая асимметрия рецептирующих элементов. Эта асимметрия заключается в том, что кйноцилия располагается с одного края по отношению к сте- реоцилиям (Lowenstein, Wersall, 1959). Например, в полукружных каналах киноцилии всех волосковых клеток ориентированы в одну сторону: в горизонтальных каналах они расположены в сторону утрикулюса, а в вертикальных каналах — в направлении от него (см. главу 10, рис. 111). Возбуждающим является стимул, действующий в направлении от стереоцилий к киноцилии (Lowenstein, Wersall, 1959; Lowesnstein et al., 1964; Wersall et al., 1965; Flock, 1971a, и мн. др.).
Исследования импульсной активности волокон полукружных каналов показали, что у скатов можно выделить три основных вида сенсорных элементов (Groen et al., 1952; Groen, 1961b). Первые из них возбуждались при одном направлении раздражения и угнетались при другом; другие угнетались при действии сильного стимула; наконец, последние, не обладавшие спонтанной активностью, только возбуждались при сильных воздействиях. Принципиально такие же элементы были описаны и у млекопитающих (Gernandt, 1949).
Угловое ускорение может рассматриваться как адекватное воздействие для рецепторов полукружных каналов. Проведенное на лягушках исследование явления адаптации в рецепторах с помощью такого рода стимулов (адаптация определялась по снижению импульсной активности в афферентных волокнах) показало (Precht et al., 1971; Precht, Limas, 1972), что 35% волокон от горизонтального канала связаны с очень медленно адаптирующимися (неадаптирующимися?) элементами. Остальные 65% волокон обладали быстро адаптирующимся импульсным ответом. Усиление раздражения (в экспериментах применяли шагообразное увеличение ускорения) давало линейное приращение эффекта (частоты импульсации) в волокнах, связанных с медленно адаптирующимися рецепторами. Быстро адаптирующиеся структуры не обладали подобной способностью.
По характеру импульсных ответов в нервных волокнах от отолитовых рецепторов у скатов можно также различить медленно и быстро адаптирующиеся элементы (Lowenstein, Roberts, 1949). Рецепторы первого типа (статорецепторы) сигнализируют о положении головы в пространстве, а второго типа — о перемещениях в пространстве. Принципиально сходные данные были получены и на рецепторах утрикулярной и саккулярной макул у обезьян (Werner, Whitsei, 1968 — цит. по: Tamar, 1972).
Рецепторы отолитовых органов могут воспринимать не только линейное ускорение, но и звуковые, а также вибрационные стимулы (Ashcroft, Hallpike, 1934; Ross, 1936; Lowenstein, Roberts, 1949, 1951; Enger 1968; Lowenstein, 1971).
В статорецепторах лягушки частота импульсации может меняться пропорционально логарифму силы воздействия (Gual- tierotti, Gerathewohl, 1965).
Рецепторы утрикулюса лягушки исследовались и в условиях невесомости. Оказалось, что в состоянии невесомости частота фоновой импульсации вначале возрастает, а затем может угнетаться (Gualtierotti, Alltucker, 1967).
В работах последнего времени (см.: Oman, Young, 1972) проводилась расчетная оценка пороговых смещений, необходимых для возбуждения механорецепторов полукружных каналов у человека. Эта величина равняется примерно 100 А.
Более подробные сведения об афферентной сигнализации от вестибулярных рецепторов приведены в ряде специальных обзоров (Gernandt, 1959; Lowenstein, 1967а, 1972; Кисляков и др., 1972).XI Для данного рассмотрения важно лишь то, что особых отличий от общих закономерностей импульсации других механорецепторов здесь не обнаруживается.
Импульсная активность рецепторов органа слуха
Импульсная активность нервных элементов нижних уровней слуховой системы изучалась в большом числе работ (обзоры: Hawkins, 1964; Kiang, 1965; Гершуни, 1967, 1971; Fex, 1968; Grinnell, 1969; Радионова, 1971; Eldredge, Miller, 1971; Альтман^ 1972; Evans, Wilson, 1973; Kiang et al., 1973; Pfeiffer, On Kim, 1973, и др.). В рамках данной книги нет возможности детально рассматривать вопросы этого раздела физиологии, особенно учитывая то обстоятельство, что детальный разбор их был сделан в предыдущих томах настоящего руководства (Альтман и др., 1972; Гершуни, 1972а, 19726; Молчанов, Радионова, 1972). Однако следует указать, что по сравнению с другими отделами слуховой системы импульсная активность афферентных слуховых волокон изучена, в значительно меньшей степени. Это связано, как уже отмечалось, с большими техническими трудностями при экспериментировании на лабиринте, особенно у высших позвоночных.
У кошки практически все волокна слухового нерва, которые весьма однородны по диаметру (следовательно, скорости проведения нервных импульсов в них сходны), обладают спонтанной активностью. Эта активность может быть весьма высокой и превышает иногда 100 имп./сек. (Kiang, 1965).
Фоновая активность афферентных слуховых волокон, по-видимому, обусловлена спонтанным выделением медиатора волосковыми механорецепторными клетками, что и обусловливает генерацию «спонтанных» ВПСП в афферентных окончаниях. Возникновение таких ВПСП было показано с помощью микроэлектродной техники (Furukawa, Ishii, 1967а) в экспериментах на рецепторах саккулярной макулы и лагены костистых рыб (у золотой рыбки) (рис. 59, А и 60, А).
Все изученные к настоящему времени рыбы обладают слухом, т. е. восприятием в дальнем акустическом поле периодических изменений давления в определенном частотном диапазоне. Эта функция отличается от восприятия более низкочастотных вибрационных смещений среды в ближнем от источника поле, что обеспечивают в основном механорецепторы боковой линии (см.: Вег- gejik, 1967). Правда, у рыб имеется в определенном частотном диапазоне некоторое перекрытие функций боковой линии, с одной стороны, и внутреннего уха, с другой. С помощью веберова аппарата (системы сочлененных косточек) изменения давления в плавательном пузыре (у рыб, обладающих им, в том числе у золотой рыбки) передаются на внутреннее ухо. Рецепторы саккулярной макулы воспринимают эти изменения и, следовательно, участвуют в осуществлении слуховой функции. При нанесении раздражения возникновение ВПСП начинает осуществляться синхронно со стимулами (рис. 59, £>). При низких частотах и высокой
интенсивности звука каждый ВПСП, имевший в этом случае большую амплитуду и длительность, вызывал появление двух и более спайков (рис. 61, Г). Звуки высокой частоты (порядка 600/сек.) обычно не вызывали множественных разрядов (Furu-
Рис.
59. ВПСП и ПД двух различных афферентных
волокон саккулюса у костистой рыбы
(по: Furukawa, Ishii, 1967а).


А — спонтанная активность; Б — ответ на звук умеренной интенсивности (75 дб, 250 пер./сек.). Сверху вниз: последовательные пробеги луча; нижняя запись на Б — отметка раздражения. Отметка времени: на А — 5, на Б — 2 мсек.; калибровка: на А — 10, на Б — 5 мв.
Рис. 60. Ответы афферентных волокон лагены у костистой рыбы (по: Furukawa, Ishii, 1967а).
А — групповые спонтанные разряды; Б — ответы на вибрацию в дорзо-вентральном направлении с частотой 28 пер./сек. Нижняя запись на Б — отметка раздражения. Отметка времени — 20 мсек.; калиб- овка — 5 мв.
kawa, Ishii, 1967а). Низкочастотная вибрация водной среды (28 пер./сек., рис. 60, Б) приводит к появлению пачкообразных разрядов в афферентных нервных волокнах от механорецепторов лагены (Furukawa, Ishii, 1967а).
В одних волокнах возникновение импульсов было синхронизировано с одной фазой (фазой сдавления) звукового колебатель-
кого воздействия, в других — с другой (фазой разрежения), в третьих — с обеими фазами (рис. 61, А—В). Авторы полагают, что это обусловлено наличием различной ориентации волосковых клеток в саккулярной макуле рыб. Это предположение находит подтверждение в данных морфологии (Lowenstein et al., 1964; Wersall et al., 1965; Hama, 1969). В других экспериментах было показано, что волосковые рецепторы действительно обладают дирекционной чувствительностью (Furukawa, Ishii, 1967b). Так,

Рис. 61. Ответы различных афферентных волокон саккулюса у костистой рыбы (по: Furukawa, Ishii, 1967а).
А, Б — ответы на обе звуковые фазы — сжатие и разрежение (А — частота 500, Б — 700 пер./сек.); В — ответы волокна, связанного с рецепторами, реагирующими только на фазу сжатия звука (частота — 500 пер./сек.); Г — множественные ответы волокна на сжатие звука (частота 180 пер./сек.). Сверху вниз’, импульсная активность; отметка раздражения. Отметка времени: на А—В — 1, на Г — 2 мсек.; калибровка: на А, Б, Г — 5, на В — 20 мв.
при одном направлении воздействия (смещение отолита саккулы в одну сторону) можно было отметить появление локальной деполяризации (ВПСП, или генераторного потенциала) в постсинаптических афферентных структурах, которая приводила к появлению импульсного разряда. Смещение отолита в другом направлении не вызывало появления ВПСП (см. главу 10, рис. 109).
Афферентные слуховые волокна обычно обладают способностью отвечать тоническим разрядом на длительное действие звукового стимула, но часть структур имеет фазный ответ (на включение и выключение сигнала). Судя по ответам волокон, можно говорить о наличии явлений периферического подавления, воз- пикающего, например, при одновременном действии двух звуков (см.: Радионова, 1971).
Отдельные волокна слухового нерва характеризуются различными порогами вовлечения их рецепторов в реакцию (Радионова, 1962; Katsuki et al., 1962). Зависимость порога реакции от частоты сигнала может быть выражена на основе опытов с регистрацией активности нервных волокон в виде так называемых частотнопороговых кривых (Kiang, 1965). Следует заметить, что пороговые величины раздражений, воспринимаемые механорецепторами органа слуха, чрезвычайно низки (Bekesy, 1960; Гершуни, 1972а).XII Это обстоятельство, по-видимому, связано не только со свойствами самих волосковых механорецепторов, но и с условиями их функционирования, зависящими от особенностей среды, в которой они располагаются (см. подробнее главу 7).
В целом очевидных особенностей в реакции слуховых афферентных волокон при действии адекватного стимула, которые могли бы говорить о существовании принципиальных различий в деятельности слуховых рецепторов по сравнению с другими волосковыми механорецепторными структурами позвоночных, описано не было.
* * *
Сравнение механорецепторов органов чувств с тканевыми механорецепторами позвоночных показывает, что между ними имеется определенное различие. Оно обусловлено тем, что в первом случае имеются синаптические контакты между волосковыми клетками и окончаниями афферентных волокон. Поэтому в сенсорных приборах органов чувств возникновение импульсной активности происходит в результате развития возбуждающих постсинаптических процессов, а в тканевых рецепторах — как следствие появления деполяризационных рецепторных потенциалов, обусловливающих генерацию ПД в том же самом нейрональном элементе.
Механорецепторы органов чувств обладают выраженной ди- рекционной чувствительностью. В тканевых рецепторах это явление хотя и существует, но распространено оно в меньшей степени и выявляется не столь отчетливо (подробнее см. главу 10).
Волосковые механорецепторы органов чувств обладают высокой чувствительностью, намного превышающей чувствитель
ность тканевых механорецепторных приборов. Можно считать, что эта функциональная особенность в органах чувств, по-видимому, является наиболее существенным их отличием от рецепторов в тканях (Ильинский, Красникова, 1972). Никаких иных существенных особенностей, тестируемых по импульсному разряду в афферентных волокнах, между тканевыми (первичными) и волосковыми (вторичными) механорецепторными приборами не обнаруживается. Во всех случаях наблюдаются быстро и медленно ослабляющиеся ответы, детали узора которых, равно как и следовые эффекты, определяются в первую очередь характером наносимого раздражения.
Механизмы, обусловливающие высокую чувствительность волосковых сенсорных элементов органов чувств, будут рассмотрены далее (глава 7). Отдельно будут рассматриваться и явления эфферентного регулирования, по-разному протекающего для первичных и вторичных механорецепторов позвоночных (глава 5), что, можно думать, прежде всего связано с различной чувствительностью этих сенсорных приборов к действию адекватного раздражения.
ИМПУЛЬСНАЯ АКТИВНОСТЬ
МЕХАНОРЕЦЕПТОРОВ БЕСПОЗВОНОЧНЫХ
Активность волосковых механорецепторных органов
Механорецепторые сенсиллы насекомых могут рассматриваться как типичные представители этого весьма распространенного у беспозвоночных животных типа сенсорных приборов. Импульсная активность волосковых рецепторных образований у насекомых изучалась рядом авторов на примере ветрочувствительных рецепторов (Haskell, 1958; Guthrie, 1966; Сви- дерский, 1967, 1968; Gamhi, 1969; Smola, 1970b; Свешников, 1972, и др.; см. также обзор: Свидерский, 1973).
Было установлено, что адекватным раздражением рецепторов является не их микровибрация, а отклонение потоком воздуха от исходного состояния (Свидерский, 1967а). Чем больше это отклонение, тем больше ГП рецепторов и выше частота импульсной активности. Считают, что область генерации ПД в механорецепторных сенсиллах находится у основания дендрита (Thurm, 1967, 1970) и ПД, по-видимому, не проникает в область генерации РП (Wolbarsht, 1960). Величина локальной стационарной деполяризации и частота ответа связаны между собой линейной зависимостью (рис, 62, Л), существующей в области не очень больших углов отклонения волоска (до 25°). Именно такого рода отклонения и имеют место при воздействиях, соответствующих реальным в жизни животных (Свидерский, 1969а, 19696; Smola, 1970b).
Наличие динамической и статической составляющих РП определяет проявление этих же компонентов в импульсном ответе рецептора на воздействие (рис. 62, Б), Статический компонент по-разному выражен в ответах разных рецепторов: скорость адаптации их импульсного разряда может варьировать. В целом же рецепторы должны рассматриваться как весьма медленно адаптирующиеся, фазно-тонические (Свидерский, 1967).
Динамическая фаза импульсного ответа существенно зависит от скорости нарастания стимула: чем выше эта скорость, тем более значителен динамический компонент изменения частот им-

Рис. 62. Характеристика импульсной активности ветрочувствительных рецепторов у азиатской саранчи (4 — по: Smola, 1970b; Б, В — по: Свидерский, 1967а).
А — зависимость между величиной РП и частотой импульсного ответа рецептора. По оси абсцисс — амплитуда РП (мв); по оси ординат — частота импульсации (имп./сек.). Б — действие длительного обдувания рецептора I области головы. По оси абсцисс — время от начала обдувания (сек.); по оси ординат — частота импульсации (имп./сек.). В — влияние скорости отклонения волоска на характер начального ответа рецептора III области головы. По оси абсцисс — время от начала отклонения (сек.); по оси ординат — частота импульсации (имп./сек.). 1 — быстрое («мгновенное») отклонение волоска; 2 — более медленное отклонение; з — очень медленное (несколько мкм/сек.) отклонение (угол отклонения волоска — 30°).
пульсации (рис. 62, В). Такой характер ответа весьма типичен для различных кутикулярных волосков (Haskell, 1959; Chapman, Smith, 1963; Лапицкий, 1967). При повторных раздражениях этих рецепторов можно было обнаружить уменьшение частоты ответов уже после 10—20 воздействий (Свидерский, 1967).
Представляет интерес оценка порогов возбуждения волосковых механорецепторов у насекомых. Согласно некоторым авторам (Antrum, Schneider, 1948), эта величина может не превышать 1 А. По другим данным (Thurm, 1965а), эта величина соответствует примерно 30 А. Сходные величины пороговых смещений были установлены и для других кутикулярных (лирообразных) механорецепторных структур членистоногих: 25 А (Walcott, van der Kloot, 1959) и 10 A (Liesenfeld, 1961).
Активность механорецепторов слуховой системы у насекомых
Электрофизиологическое изучение этих рецепторов осуществлялось рядом авторов (Pumphrey, Rawdon-Smith, 1936; Pumphrey, 1940; Autrum, 1960; Katsuki, Suga, 1960; Suga, i960; Попов, 1965, 1971; Michelsen, 1966, 1971a, 1971b; Петровская, 1969; Петровская, Каплан и др., 1970; Петровская и др., 1970; Жантиев, 1971; Жантиев, Чуканов, 1972, и др.; см. также обзоры: Suga, 1960; Попов, 1966; Попов, Светлогорская, 1973). В работах, посвященных изучению рецепторов тимпанального органа, было показано, что они отличаются между собой по ряду свойств: избирательной чувствительности к частоте звуков, общей чувствительности, динамическим характеристикам ответов. Однако можно полагать (см.: Попов, Светлогорская, 1973), что эти особенности определяются не свойствами собственно рецепторных элементов, а механическими характеристиками их вспомогательных аппаратов (участков тимпанальной мембраны, к которым прикреплены рецепторы). Незначительные же различия между самими рецепторами, возможно, имеются лишь в отношении скорости развития адаптационных процессов, оцениваемых по изменению импульсного разряда.
Активность волосковых механорецепторов статоциста
Импульсная активность механорецепторов органа восприятия гравитации у беспозвоночных изучена еще сравнительно неполно, несмотря на очевидную важность таких исследований (Cohen, 1955, 1960; Wolff, 1970а; Wood, Baumgarten, 1972; Alkon, Bak, 1973; Detwiler, Alkon, 1973).
Эксперименты с внутриклеточными отведениями от рецепторов статоциста у моллюска Harmissenda crassicornis показали, что при механическом раздражении рецептора возникает деполяризационный РП, который ведет к появлению импульсной активности (Alkon, Bak, 1973). Частота импульсной активности изменялась линейно с варьированием силы тока, протекавшего через рецептор.
Волосковый рецептор статоциста в покое генерирует фоновую активность небольшой частоты — от 8 до 200 имп./мин. (Alkon, Bak, 1973; Detwiler, Alkon, 1973). Рецепторы статоцистов характеризуются фазно-тоническим разрядом на стимуляцию (Cohen, 1955; Wolff, 1970а; Wood, Baumgarten, 1972). Каждый рецептор, например в статоцисте у моллюска Pleurobrachaea californica, реагирует возбуждением лишь в определенном диапазоне положений волоска. Каждое положение в пределах этого диапазона характеризуется одинаковой стационарной частотой разряда. Начальная же частота ответа может варьировать и зависит от на
правления воздействия, т. е. имеет место дирекционная чувствительность рецепторов (Wood, Baumgarten, 1972). Ответы, принципиально сходные с только что описанными, наблюдались также у одного из двух выявленных типов сенсорных элементов у омара (Cohen, 1955). Дирекционная чувствительность была отмечена также в статорецепторах у краба Scylla (Sandeman, Okajima, 1972). ~
Активность мышечных рецепторов растяжения у ракообразных
По достижении деполяризационным РП критического уровня (10—15% от величины потенциала покоя в случае МРОХ и несколько больше в случае МРО2) рецепторы растяжения начинают генерировать импульсную активность (Eyzaguirre, Kuffler, 1955а). Амплитуда спайков МРО2 обычно больше, чем в’МРОр Импульсации МРОг продолжается в течение всего периода действия раздражения (рис. 63, 4), что позволяет считать его мед-

Рис. 63. Ответ медленно (Л) и быстро (Б) адаптирующихся рецепторов растяжения на длительное раздражение (по: Eyzaguirre, Kuffler, 1955а).
Стрелки: 1 — нанесение раздражения, 2 — его усиление (на Б дважды), з — прекращение раздражения. Прерывистая линия — критический уровень деполяризации. Отметка времени — 1 сек.; калибровка — 50 мв.
ленно адаптирующимся рецептором. В то же время МРО2 бывает активным лишь в начальный период действия стимула (рис. 63, Б), поэтому МРО2 рассматривается как быстро адаптирующийся механорецептор (Wiersma et al., 1953; Kuffler, 1954; Eyzaguirre, Kuffler, 1955a, 1955b). Каждому ПД предшествует появление препотенциала, а за генерацией спайка обычно следует и реполяризация рецепторного нейрона, при которой уровень мембранного потенциала оказывается на 5—10 мв ниже исходного значения потенциала клетки.
В опытах на ракообразных с их периферически расположенным сенсорным нейроном обнаружено, что генерация ПД происхо дит в аксоне на некотором расстоянии (200—500 мкм) от тела нейрона (Edwards, Ottoson, 1958; Grampp, 1966; Washizu, Ter- zuolo, 1966), где на протяжении нескольких десятков микрон имеется некоторое сужение нервного волокна и где плотность деполяризующих токов, обусловливающих появление генераторного потенциала, максимальна. Порог возникновения ПД в этой области наименьший. От места своего возникновения ПД распространяется как ортодромно по аксону в центральном направлении, так и антидромно на сому клетки и область дендритных разветвлений (до 150 мкм вдоль проксимальной части дендритов). В целом по направлению к дендритам электрическая возбудимость мембраны самой клетки значительно снижается.. Сами же дендриты, невидимому, или обладают незначительной электрической чувствительностью, или не обладают ею совсем (Grampp, 1966).
Наличие триггерной зоны в аксоне сенсорного нейрона не означает, что ПД не могут возникать и в других, менее чувствительных областях рецептора. При определенных условиях ПД могут возникать и в области самой клетки (Kuffler, 1960). В частности, такого рода явления могут иметь место при возникновении так называемых групповых разрядов (Eyzaguirre, Kuffler, 1955b). В их основе лежит длительная деполяризация сомы. Она может быть вызвана рядом причин, например нагреванием (Florey, 1956; Burkhardt, 1959; Grampp, 1966), действием стрихнина (Washizu et al., 1961; Washizu, 1965; Русинов, Эзрохи, 1967a, 19676, 1967в; Эзрохи, 1969, 1972), влиянием ряда других веществ и ионов (Washizu, 1965; Washizu, Terzuolo, 1966) и т. д. Деполяризация нейрона связана с появлением в соме более длительных ПД. Увеличение длительности ПД развивалось за счет резкого замедления его реполяризационной фазы. На фоне длительной деполяризации сомы может иметь место осциллирование мембранного потенциала, что в свою очередь приводит к генерации ПД. При этом количество ПД, генерируемых сомой и аксоном, в связи с появлением аксо-соматической задержки может существенно разниться как в одну, так и в другую сторону, и даже в несколько раз (Washizu, 1965; Русинов, Эзрохи, 1967а; Эзрохи, 1969). (Следует заметить, что не всегда длительная деполяризация рецептора может приводить к появлению групповых разрядов — см., например, о действии пенициллина: Ayala et al., 1970).
В целом, явления, развертывающиеся в рецепторной клетке при генерации ПД, оказались принципиально такими же, как и в других нейронах, в частности в мотонейронах позвоночных (Araki, Otani, 1955, 1959; Eccles, 1957, 1964; Fuortes et al., 1957; Sasaki, Otani, 1962, и др.).
Наличие на периферии сомы сенсорной нервной клетки приводит к определенным особенностям в генерации ПД в рецепторах растяжения ракообразных по сравнению, например, с мышечными веретенами. Однако можно считать, что нет принципиальных различий между этими сенсорными приборами ни в последовательности развертывающихся событий, ни даже в рисунке импульсного разряда, регистрируемого в нервных волокнах при сходных воздействиях на рецепторы.
Исследование влияния амплитуды стационарной локальной деполяризации (генераторного потенциала) на частоту генерации импульсной активности медленно адаптирующегося рецептора растяжения показало существование пропорциональной зависимости между этими величинами (Terzuolo, Washizu, 1962). Соответственно, частота импульсации была линейно связана с величиной растяжения (Burkhardt, 1959; Brown, Stein, 1966) и с логарифмом груза, приложенного к МРО (рис. 34). Считают, что частота импульсации почти линейно связана с длиной мышцы и с мышечным напряжением (Brown, Stein, 1966). Эти же авторы на основании количественного анализа полагают, что параметр напряжение мышцы, по-видимому, более тесно, чем длина, связан с параметром частоты импульсной активности.
При увеличении раздражения выше некоторого критического значения частота импульсации в МРО падает, и активность может даже полностью блокироваться (появление феномена перерастя- жения рецептора). Механизм этого явления связан с развитием катодического блока (Eyzaguirre, Kuffler, 1955а; Terzuolo, Washizu, 1962).
Наряду с зависимостью от амплитуды раздражения величина ответа МРО обусловливалась также и скоростью нарастания стимула. Чем выше была эта скорость, тем больше была частота импульсации в динамической фазе реакции. Статическая же составляющая ответа не зависела от скорости воздействия, а определялась исключительно установившейся величиной растяжения рецептора.
Исследование действия синусоидального растяжения показало, что при небольших амплитудах воздействия реакция МРО линейно связана с частотными характеристиками раздражения (Borsellino et al., 1965).
В состоянии покоя рецепторы находятся не всегда. Во многих случаях (что зависело, например, от сезонных изменений) отмечается стационарная фоновая активность МРО1? а иногда даже — фоновая импульсация МРО2. При нанесении раздражения оба вида рецепторов отвечают всплеском импульсации, которая затем (в пределах нескольких десятков секунд для МРОХ и значительно быстрее для МРО2) снижается у МРОХ до какого-то постоянного уровня, определяемого величиной стационарного растяжения, или же до нуля у МРО2. При уменьшении раздражения фоновая активность рецепторов может резко снижаться, вплоть до исчезновения, а затем может постепенно восстанавливаться у МРОГ до уровня, опять-таки зависящего от новой величины стационарного воздействия.
Кривая восстановления фоновой активности зависит от силы и длительности раздражения. Угнетение рецепторной активности в период, следующий за моментом снятия раздражения, определяется возникновением гиперполяризационного РП и развитием посттетанической депрессии (Eyzaguirre, Kuffler, 1955а; Nakajima, Takahashi, 1966; Nakajima, Onodera, 1969b).
Таким образом, картина изменения импульсации рецепторов растяжения у ракообразных весьма сходна с картиной изменения активности других рецепторов при аналогичных условиях раздражения.
Изучение следовых гиперполяризационных сдвигов потенциала МРО в период сразу после генерации импульсной активности явилось предметом специального рассмотрения (Nakajima, Takahashi, 1966). Было установлено существование двух групп явлений. Во-первых, отмечалась кратковременная гиперполяризация после генерации одиночного спайка или небольшой группы спайков. Во-вторых, было обнаружено существование долго длящейся гиперполяризации после возникновения длительных разрядов рецептора (посттетаническая гиперполяризация). Удалось найти способы различного изменёния этих видов гиперполяризации. Здесь имеется в виду, что под влиянием анодического тока происходит уменьшение кратковременной длительной посттетанической гиперполяризации. Опыты с применением антидромной стимуляции, ионов калия, натрия и лития, а также 2,4-динитрофенола позволили заключить, что кратковременная гиперполяризация обусловлена увеличением проницаемости мембраны для ионов калия, в то время как длительная посттетаническая гиперполяризация определяется работой электрогенного натриевого насоса.
Импульсная активность рецепторов отражает не только те или иные стороны раздражающего воздействия. Она несет в себе и изменения, обусловленные меняющимися свойствами самих сенсорных приборов. В частности, она испытывает влияния клеточных «шумов» и молекулярных процессов, аналогичные тем, которые имеют место в самых разных нервных образованиях (Verveen, 1961; Derksen, 1965; Verveen, Derksen, 1965; Verveen, DeFelice, 1974). Исследование флуктуаций межспайковых интервалов импульсной активности, генерируемой рецептором растяжения у ракообразных, показало (Firth, 1966), что они чрезвычайно малы. При средней величине межспайкового интервала (г) (80 мсек, коэффициент вариаций был менее 0.8%. Аналогичный показатель для других механорецепторов был значительно выше: 3% для тактильной корпускулы в коже кошки (Werner, Mountcastle, 1965), 2.7% (Buller et al., 1953) и 31% (Hagiwara, 1954) для мышечного веретена кошки (в последнем случае т равнялось 10 и 100 мсек, соответственно). Для сравнения можно указать, что, например, аналогичная величина для таламического лемнискового нейрона равнялась 42% (Poggio, Viernstein, 1964). У рецепторов растяжения величина стандартного отклонения интервалов линейно связана с длительностью среднего межспай- кового интервала в диапазоне исследованных длительностей вплоть до 80 мсек, и возрастает по мере увеличения продолжительности т (Firth, 1966).
В ряде публикаций анализировалась передаточная функция МРО при использовании более или менее сложных моделей работы этого сенсорного прибора (Borsellino et al., 1965; Brown, Stein, 1966; Terzuolo, Bayly, 1968; Enger et al., 1969).
Активность эластических органов
В литературе имеется ряд исследований, посвященных изучению деятельности различных проприоцепторных хордотональных и миохордотональных органов у высших беспозвоночных, и в частности типичного представителя этих сенсорных образований — проподит-дактилоподитного органа (ПД-органа) ракообразных (Burke, 1954; Wiersma, 1959; Wiersma, Boettiger, 1959; Cohen, Dijkgraaf, 1961; Hwang, 1961; Cohen, 1963, 1965; Mendelson, 1963; Wyse, Maynard, 1965; Hartman, Boettiger, 1967; Boettiger, Hartman, 1968; Clarac, 1968; Бурданов и др., 1971a, 19716, 1973; Купцова, Шурубура, 1971; Бурданов, Сотник, 1972, и др.; см. также обзор: Finlayson, 1968).
При микроэлектродном исследовании было показано, что возникновение импульсной активности рецепторов ПД-органа связано с появлением локальной деполяризации (ГП) в сенсорном нейроне (Mendelson, 1963).
С помощью техники записи от одиночных волокон было выявлено существование двух основных видов механорецепторов в ПД- органе: один вид рецепторов (тонический) сигнализирует о положении сустава, а другой (фазный) реагирует в ответ на движение в суставе (Wiersma, Boettiger, 1959). Рецепторы последнего типа отвечают на движение только в одном направлении. Часть рецепторов отвечает на движение в направлении, вызывающем укорочение эластического пучка, с которым связаны сенсорные клетки, а часть — при движениях, обусловливающих его удлинение (при флексии или экстензии сустава) (Wiersma, 1959; Mendelson, 1963). Таким образом, адекватным стимулом для рецепторов можно считать изменение длины эластического пучка суставного органа.
Рецепторные элементы обладают различными порогами к влиянию скорости воздействия. Пороговая скорость колеблется от 0.2 до 5 град./сек. При скорости выше пороговой частота импульсов, генерируемых рецепторами движения, изменяется в логарифмической зависимости от изменения скорости (Бурданов, Сотник, 1972). При увеличении скорости раздражения разряды становятся более регулярными и частота их возрастает обычно до величины порядка 40 в 1 сек. (Mendelson, 1963).
Адаптация рецепторов движения наблюдается в течение 1 — 1.5 сек. после нанесения раздражения. Рецепторы положения могут генерировать импульсную активность после фиксации нового положения сустава в течение 25 мин. Для нейронов положения максимальная частота (5—50 имп./сек.) меньше, чем для рецепторов движения (до 200 имп./сек.) (Бурданов, Сотник, 1972).
Для каждого рецепторного нейрона существует свой рабочий диапазон углов положения сочленений, который, как правило, меньше полного угла (60°) раскрытия клешни. В пределах каждого рабочего диапазона имеется угол, при котором можно отметить максимальный ответ рецептора. Обычно такое положение существует вблизи одной из границ диапазона работы рецептора. Практически все рецепторы могут быть подразделены на нейроны, частота активности которых может быть максимальной либо в начале, либо в конце движения. Рецепторные элементы функционируют независимо друг от друга (Бурданов и др., 1973). Наличие в ПД-органе большого количества рецепторных нейронов (порядка 160), имеющих различные рабочие диапазоны, несомненно позволяет с помощью данного сенсорного органа сигнализировать нервной системе о всех перемещениях в ПД-суставе.
* * *
Как видно из изложенного, метод регистрации импульсной активности от различных частей сенсорных нейронов (чаще всего от их афферентных нервных волокон) позволил выявить наличие у животных разнообразных механорецепторных структур, локализованных во всех органах и тканях организма. При этом не всегда удается определить, какая именно сенсорная структура ответственна за механорецепторную функцию. В ряде случаев (например, при изучении нервных окончаний мышечных веретен) именно характеристики нервных проводников способствуют идентификации механорецепторных приборов.
В настоящее время нет оснований считать, что структуры, генерирующие спайки в механорецепторных приборах, отличаются какими-либо принципиальными особенностями по сравнению со свойствами регенеративных механизмов обычных нервных проводников, хотя и нельзя исключить наличия у механорецепторов некоторых особенностей. Указанное сходство позволяет привлечь для анализа импульсной активности в рецепторах сведения, полученные при изучении других электровозбудимых мембран нервных элементов.
По своему импульсному ответу на однотипное воздействие все механорецепторы могут быть подразделены на две основные группы: быстро и медленно адаптирующиеся (фазные и тонические). В импульсной реакции медленно адаптирующихся механорецепторов на включение раздражения обычно легко могут быть прослежены две фазы: динамическая и статическая, характеристики которых определяются, с одной стороны, параметрами стимула (его амплитудой и скоростью нарастания) и, с другой стороны, индивидуальными особенностями данного сенсорного прибора. У быстро адаптирующихся рецепторов регистрируется только динамическая фаза ответа. При снятии раздражения в реакции рецепторов тонического типа, как правило, наблюдается период сниженной возбудимости и торможение фоновой импульсации (если фоновая активность имела место). У рецепторов фазного типа в этом случае часто возникает всплеск активности (ответ на выключение).
В тех случаях, когда имелась возможность сопоставить импульсный разряд механорецептора с характером развития его генераторного потенциала, можно было обычно проследить линейную зависимость частоты импульсации от величины локальной деполяризации элементов сенсорного нейрона. Выявление такой зависимости позволяет с большой долей вероятности постулировать наличие такого же рода связи и для случаев, когда генераторный потенциал механорецепторных приборов не изучался. Именно данное сопоставление позволяет полагать, что динамическая и статическая фазы разряда медленно адаптирующихся рецепторов также обусловлены соответствующими фазами в развитии ГП.
Период угнетения деятельности медленно адаптирующихся рецепторов, по-видимому, обусловлен развитием гиперполяризационного РП, а также посттетанической депрессией, характерной для всех регенеративных систем. Выше приводились основания для утверждения, что ответы на выключение у быстро адаптирующихся механорецепторов, вероятно, в большинстве случаев (если не во всех) обусловлены механическими свойствами вспомогательного аппарата рецепторных приборов, а также свойствами окружающих их тканей. Наконец, у многих механорецепторов позвоночных и беспозвоночных животных на основании изучения их импульсных ответов обнаружена чувствительность к направлению действия адекватного раздражения.
Хотя отдельные параметры раздражения, такие как амплитуда, скорость нарастания и спадения, длительность, направление и т. д., могут быть эффективно оценены индивидуальными механорецепторами, по-видимому, следует полагать, что основное кодирование признаков воздействия должно осуществляться популяциями рецепторов. Этому способствует перекрытие рецептивных полей, наличие различных фазных и тонических рецепторов, разнопороговость сенсорных элементов, наличие рецепторов, обладающих различной чувствительностью к направлению действия стимула и т. д.
Резюмируя весь представленный материал, можно утверждать, что нет какой-либо принципиальной разницы в общем характере импульсных ответов механорецепторов, относящихся к первичным и вторичным сенсорным приборам, равно как и в случае механорецепторов позвоночных и беспозвоночных животных. Конечно,
цепь процессов, протекающих в первичных и вторичных механорецепторах, существенно различается, однако импульсный ответ на однотипное воздействие, наблюдаемый в этих двух основных типах сенсорных приборов позвоночных, может мало разниться. Наиболее заметные отличия в рисунках импульсных разрядов разных механорецепторов зависят от характера адекватного для них раздражения: движений частей тела, звуковых стимулов, пульсовых толчков, сокращений стенок полых органов, тактильных воздействий и т. д.
Единственное существенное различие, которое может быть выявлено при анализе импульсной активности механорецепторов, заключается в их неодинаковой чувствительности к действию адекватных раздражений. (Следует отметить, что прежде всего именно порог возникновения импульсного ответа обусловливает пороги, определяемые в психофизических и поведенческих экспериментах). У позвоночных животных обнаруживается значительно более высокая чувствительность волосковых механорецепторов органов чувств по сравнению с чувствительностью тканевых механорецепторов. В этой связи можно полагать, что именно волосковая сенсорная клетка и обеспечивает повышенную чувствительность механорецепторов органов чувств к действию адекватного раздражения. У беспозвоночных животных во всех механо- рецепторных приборах, которые являются первичными сенсорными приборами, также можно обнаружить структуры, значительно различающиеся по своей чувствительности (например, волосковые рецепторы органа слуха и мышечные рецепторы). Существенным является то, что и в этом случае более чувствительные механорецепторы содержат в своей структуре реснитчатые или волосковые образования. Более детально вопросы, связанные с чувствительностью механорецепторов, будут рассмотрены далее (главы 7 и 11, стр. 378, 474).