

Глава 10
ЧУВСТВИТЕЛЬНОСТЬ МЕХАНОРЕЦЕПТОРОВ К НАПРАВЛЕНИЮ ДЕЙСТВИЯ РАЗДРАЖИТЕЛЯ.
ГИПЕРПОЛЯРИЗАЦИОННЫЕ РЕЦЕПТОРНЫЕ ПОТЕНЦИАЛЫ
Среди различных параметров раздражающего воздействия организму очень важно оценить направление действия стимула. Достигается это в различных случаях по-разному. Так, прежде всего животное само может перемещаться по отношению к источнику сигнала и менять тем самым его действие на данную рецепторную поверхность, т. е. ориентировать рецепторные приборы в направлении источника раздражителя. Возможно избирательное перемещение рецепторного аппарата или даже отдельных его частей в направлении действующего стимула, например, движение ушных раковин. Способность оценивать направление действия может достигаться и за счет специального устройства вспомогательных структур органа или даже рецептора. Так, например, у беспозвоночных может быть обеспечена направленная (дирекционная) чувствительность некоторых механочувстви- тельных волосков, которые имеют специальные вспомогательные образования, обеспечивающие избирательное направление собственных колебаний волоска. Другими примерами могут служить полукружные каналы вестибулярного аппарата или каналы боковой линии, расположение которых исходно ограничивает возможные направления перемещения заключенной в них жидкости и тем самым помогает оценивать направление действия раздражителя.
Наконец, что и представляет наибольший интерес в связи с механизмами деятельности механорецепторов, сами рецепторные структуры также могут обладать способностью оценивать направление воздействия. Эта способность, или дирекционная чувствительность (directional sensitivity), выражается в том, что рецептор максимально возбуждается лишь при каком-то определенном направлении действия стимула, а при другом направлении он не возбуждается или же даже угнетается. Угнетение рецепторного прибора, тестируемое по импульсной активности, возникает в связи с развитием гиперполяризационных явлений.
Гиперполяризационные ответы рецепторных приборов изучались значительно меньше, чем деполяризационные, хотя во многих работах отмечалось существование явлений периферического торможения (см. обзоры: Granit, 1955; Ильинский, 1967; Fuortes, 1971; Tomita, 1972).
Можно различать два типа гиперполяризации сенсорных образований: первичную и вторичную. Первый тип гиперполяризационных ответов развивается первично под действием адекватного раздражения и предшествует возникновению каких бы то ни было деполяризационных реакций. Второй тип ответов в той или иной степени обусловлен начальными деполяризационными процессами, развивающимися в рецепторах, и следует за ними, т. е. появляется вторично. Первичные гиперполяризационные ответы в принципе могут возникать и после деполяризационных явлений, но они не обусловлены ими (Ильинский, 1966а).
Изучение первичной гиперполяризации было начато, по-видимому, на волосковых механорецепторах органов чувств позвоночных. При этом исследователи вначале изучали не гиперполяризацию рецепторов, о существовании которой они вначале и не подозревали, а явление чувствительности сенсорных приборов к направлению действия раздражающего воздействия.
Следует заметить, что развитие гиперполяризации в механорецепторах сопровождается угнетением импульсной активности, если таковая наблюдалась в исходном состоянии. Если же исходно импульсной активности не было, то обнаружить возникновение гиперполяризационных реакций, приводящих к угнетению рецепторов, не представлялось возможным. Однако практически всегда можно было наблюдать (даже по интегральным показателям, основанным на реакциях различных функциональных систем), что не при всех направлениях воздействия имеет место возбуждение. На ранних этапах исследований деятельности сенсорных приборов интегральные формы оценки были ведущими. Поэтому первоначальные сведения о торможении в механорецепторах были получены именно в работах авторов, обративших внимание на роль направления раздражения в деятельности механорецепторов.
В данной главе будут представлены вначале материалы, касающиеся дирекционной чувствительности и гиперполяризационных РП в механорецепторах органов чувств, а затем в инкапсулированных тканевых сенсорных структурах. В литературе имеется довольно много данных по дирекционной чувствительности ме- ханорецепторных приборов у беспозвоночных, однако сведений о явлениях торможения в этих рецепторах под влиянием адекватных воздействий почти нет. Неравномерность разработки темы настоящей главы применительно к разным типам механорецепторов выражена весьма сильно. Естественно, что и в тексте главы это положение также проявляется: представления о гиперполяризационных РП в волосковых механорецепторах органов чувств позвоночных и в тельцах Пачини аргументированы детально, тогда как аналогичные феномены для некоторых других механорецепторных структур лишь предполагаются.
Чувствительность механорецепторов органов чувств позвоночных к направлению воздействия
Рецепторы вестибулярного аппарата. Физиологами уже давно были получены данные, говорящие о том, что кристы вестибулярных каналов являются сенсорными структурами, весьма чувствительными к направлению действия раздражения (Brown, 1874; Ewald, 1894; Maxwell, 1923; McNally, Tait, 1925, 1933; Lorente de N6, 1931; Steinhausen, 1935, и др.). Так, например, было отмечено (Ewald, 1892), что ампулоцетальная девиация купул горизонтальных каналов вызывает более сильную реакцию, чем соответственная ампулофугальная девиация. Напротив, в вертикальных полукружных каналах действенным оказывается ам- пулофугальное воздействие.
По мере применения более совершенной техники исследования удалось не только подтвердить, но и уточнить ранее полученные данные. Оказалось, что многие рецепторы полукружных каналов находятся даже при покое в состоянии постоянного возбуждения. В этих условиях ампулопетальная девиация приводит к увеличению фоновой активности рецепторов горизонтальных каналов, а ампулофугальная тормозит ее. В вертикальных полукружных каналах функциональное значение направления раздражения было обратным. В момент прекращения раздражения имела место «реакция отдачи»: за торможением следовало возбуждение и наоборот — после возбуждения отмечалось угнетение (Ashcroft, Hallpike, 1934; Mowrer, 1935; Lowenstein, Sand, 1936, 1940a, 1940b; Ross, 1936; Zotterman, 1943; Ledoux, 1949, 1958; Groen et al., 1952; Lowenstein, 1955; Groen, 1961b; см. также обзоры: Lowenstein, 1950, 1967a, 1972; Gernandt, 1959; Trincker, 1962; Кисляков и др., 1972; Precht, Llinas, 1972). Вместе с тем некоторые авторы (Adrian, 1943; Gernandt, 1949; Eckel, 1954), регистрируя биоэлектрическую активность в вестибулярных ядрах, наряду с нейронами, связанными с обычными «однонаправленными» сенсорными элементами, отмечали существование также и таких клеток, которые отвечали увеличением числа разрядов на раздражение, действовавшее в обоих направлениях. Это привело к предположению, что для некоторых рецепторов фактор направления не имеет значения.
Электрофизиологическими методами удалось продемонстрировать роль направления раздражающего стимула в деятельности рецепторных элементов утрикулярной макулы. В этом случае фоновая импульсная активность в эфферентных нервных волокнах также увеличивалась при одном направлении раздражения
и уменьшалась при противоположном (Lowenstein, Roberts, 1949). Однако и в этом случае были обнаружены отдельные рецепторные элементы, которые отвечали возбуждением вне зависимости от направления раздражения.
 Механорецепторы
органов боковой линии. Была
также продемонстрирована чувствительность
механорецепторов органов боковой
линии к направлению действия стимула
(Sand, 1937). В дальнейшем эти результаты
были подтверждены и другими исследователями
(Katsuki et al., 1951с; Dijkgraaf, 1956; Corner, 1961, 1963; Flock,
1965a, 1965b, 1967; обзоры: Dijkgraaf, 1963; Flock, 1971a,
1971b). В этих опытах было установлено, что
при раздражении боковой линии некоторых
рыб и земноводных струей
Механорецепторы
органов боковой линии. Была
также продемонстрирована чувствительность
механорецепторов органов боковой
линии к направлению действия стимула
(Sand, 1937). В дальнейшем эти результаты
были подтверждены и другими исследователями
(Katsuki et al., 1951с; Dijkgraaf, 1956; Corner, 1961, 1963; Flock,
1965a, 1965b, 1967; обзоры: Dijkgraaf, 1963; Flock, 1971a,
1971b). В этих опытах было установлено, что
при раздражении боковой линии некоторых
рыб и земноводных струей
Рис. 109. Чувствительность волосковых клеток саккулюса костистой рыбы к направлению возбуждающего воздействия (по: Furukawa. Ishii, 1967b).
1 — ВПСП и ПД в ответ на повышение давления (смещение отолита в одном направлении); 2 — отсутствие ответа при уменьшении давления (смещение отолита в противоположном направлении); з — запись повышения давления; 4 — запись понижения давления. Внутриклеточное отведение от афферентных нервных волокон. Отметка времени — 5 мсек.; калибровка — 5 мв.
жидкости, направленной в одну сторону, наблюдается увеличение фоновой импульсации в одних и тех же афферентных волокнах, в то время как при изменении направления тока жидкости на 180° отмечается ее угнетение. В других афферентных волокнах функциональная значимость направления раздражения была обратной.
Рецепторы органа слуха. Исследование вопроса о дирекционной чувствительности рецепторов органа слуха было задержано по методическим причинам. Тем не менее и в этом случае удалось доказать чувствительность механорецепторов к направлению воздействия. Так, например, в экспериментах на саккулярной макуле рыб было показано (Furukawa, Ishii, 1967а), что в одних афферентных волокнах регистрируется импульсная активность синхронно с компрессионной фазой звукового стимула, в других — с декомпрессионной фазой, а в третьих — синхронно с обеими фазами (рис. 61). Интересно, что в последнем случае частота импульсации в некоторых волокнах достигала 1500/сек. при звуковом раздражении 750/сек.
Смещение отолита в одном направлении вызывало появление значительного ВПСП в афферентном волокне и генерацию в нем 426
серии распространяющихся импульсов (рис. 109) (Furukawa, Ishii, 1967b). Напротив, смещение отолита в противоположном направлении "не вызывало деполяризационных ответов.



Рис. ПО. Связь между структурной асимметрией волосковых клеток и их чувствительностью к направлению стимула (Л — по: Flock, Wersall, 1962; Б — по: Flock, 1971b).
А — гипотетическая схема зависимости микрофонных потенциалов от ориентации волосковых клеток, а — смещение купулы ампулярной кристы (вверху) и купулы органа боковой линии (внизу). ВНл1 и ВНл2 — волосковые клетки, ориентированные в одном и том же направлении в ампулярной кристе и различно в органе боковой линии. На схемах: белые кружки — стереоцилии, черный кружок — киноцилия. Направление смещения купулы, совпадающее с направлением от стереоцилий к киноцилии, приводит к возбуждению клетки. По оси абсцисс — время; по оси ординат — направление и амплитуда смещения купулы. б — микрофонные потенциалы ампулярной кристы и органа боковой линии. Сплошная линия — суммарный потенциал, прерывистые — предполагаемые ответы ВКл1 и ВКл2. По оси абсцисс — время; по оси ординат — амплитуда потенциала. Б — внутриклеточная запись РП двух (а, б) противоположно ориентированных волосковых клеток органа боковой линии налима Lota lota; в — запись стимула. Отметка времени — 10 мсек.
Влияние направления раздражения на микрофонные потенциалы. Изучение микрофонного потенциала показало, что обычно в кортиевом. органе и в ампулярных кристах частота колебаний суммарного потенциала следует за частотой приложенного стимула (Bleeker, de Vries, 1949; Bekesy, 1952; Tasaki et al., 1954). В органах боковой линии наблюдается удвоение частоты микрофонного потенциала по сравнению с частотой раздражения (Jielof et al., 1952; Hairis, Bergeijk, 1962; Flock, 1965a, 1971b). В первом случае деполяризационная реакция возникала либо лишь на восходящую, либо лишь на нисходящую части приложенного сигнала (например, синусоиды). Во втором же случае каждая из обеих частей раздражающего стимула вызывала появление ответа (рис. 110, А).
В кортиевом органе (Bekesy, 1953) и в ампулярной кристе полукружных каналов (Lowenstein, Sand, 1940а; Trincker, 1957) было обнаружено, что смещение соответственно тенториальной мембраны и купулы лишь в определенном направлении приводит к деполяризационным изменениям (снижение эндолимфатического потенциала) и одновременно к увеличению импульсации в афферентных нервах. Напротив, смещение в противоположном направлении ведет к гиперполяризационным изменениям (повышение эндолимфатического потенциала) и к уменьшению сцай- ковой активности сенсорных нервов. Естественно, что представляло большой интерес выяснить причину функциональной полярности волосковых механорецепторов.
Морфологическая асимметрия
как основа дирекционной чувствительности
Из электронномикроскопических исследований (стр. 47) известно, что в сенсорных клетках органов боковой линии и вестибулярного аппарата киноцилии всегда располагаются на краю пучка волосков (рис. И). На этом основании можно было рассматривать клетки как морфологически асимметрично организованные структуры. У рецептирующей клетки органа слуха млекопитающих киноцилия хотя и отсутствует (иногда имеется лишь ее рудимент), но сохраняется эквивалент базального тельца — центриоля, — которая также лежит асимметрично по отношению к W-образно расположенным стереоцилиям. Лё- венстайн и Верселл (Lowenstein, Wersall, 1959), одними из первых описавшие полярную организацию рецепторных элементов в ампулярном гребешке, обратили внимание на то обстоятельство, что киноцилии волосковых клеток полукружных каналов всегда ориентированы определенным образом: в горизонтальных каналах они расположены в сторону утрикулюса, а в вертикальных каналах — от него (рис. 111). Указанные авторы сопоставили эти морфологические наблюдения с известными физиологическими данными о направленной чувствительности элементов crista ampullaris к раздражающему воздействию и пришли к заключению, что волосковые клетки полукружных каналов возбуждаются при действии механического раздражения лишь в том случае,
Налим
II
Лягушка
Черепаха
/
Латер.
Голубь
/Мед.
Варан
Рис.
111. Схема ориентации волосковых клеток
различных участков сенсорного
эпителия лабиринта у налима (Z) и
утрикулюса у некоторых позвоночных
(II)
(I
— по: Wersall et al., 1965; II
—
по: Винников и др., 1971).













Стрелки — ориентация киноцилий. Пунктир — разграничение областей различной ориентации волосковых клеток.
если оно направлено от стереоцилий к киноцилии. При обратном направлении раздражения волосковые клетки угнетаются.
Указанная гипотеза, хотя и не вскрывала причины функциональной полярности рецепторов, однако выявила его морфологический эквивалент и могла служить отправной точкой для дальнейших исследований. Последующие работы ряда авторов показали полное соответствие между морфологической и функциональной асимметрией волосковых клеток в различных органах (Gibbons, 1961; Wersall, 1961; Engstrom et al., 1962; Flock, Wersall, 1962, 1963; Flock et al., 1962; Dijkgraaf, 1963; Gorner, 1963; Flock, 1964, 1965a, 1965b, 1971a, 1971b; Lowenstein et al., 1964; Spoendlin, 1964; Wersall, Flock, 1964; Flock, Duvall, 1965; Furukawa, Ishii, 1967a, 1967b; Hama, 1969, и др.).
На основании этих экспериментов можно заключить, что во всех случаях максимальное возбуждение сенсорной клетки происходит лишь при направлении раздражающего воздействия от стереоцилий к киноцилии с ее базальным тельцем и ножкой (или к цен- триоле). При противоположном направлении стимула наблюдается максимальное угнетение. Интересно, что сама киноцилия располагается таким образом, что прямая, соединяющая две ее центральные фибриллы (рис. 13), всегда оказывается перпендикулярной к направлению максимального действия стимула.XVII В ампулярной кристе горизонтальных каналов киноцилии практически всех сенсорных клеток направлены в сторону утрикулюса, а в вертикальных каналах —от него (рис. Ill, I). Поэтому понятно, что в первом случае возбуждающим будет ампулопеталь- ное раздражение, а во втором — ампулофугальное. В утрикуляр- ной макуле ориентация сенсорных элементов оказалась более сложной (рис. 111, /). Одни рецепторы клетки оказались расположенными киноцилиями к периферии макулы, другие же — киноцилиями внутрь ее. У разных животных ориентация рецепторов в пределах макулы, как показали дальнейшие исследования, оказалась различной (см. рис. Ill, II). Кроме того, в макуле были обнаружены отдельные сенсорные клетки, которые имели две киноцилии, направленные в противоположные стороны. Такие элементы, состоящие как бы из двух обычных клеток, по-видимому, могут возбуждаться при обоих направлениях раздражения. В органах боковой линии волосковые структуры механорецепторов оказались расположенными строго вдоль продольной оси каналов, однако часть волосковых клеток направлена своими киноцилиями в одну сторону, а другая — в противоположную. В кортиевом органе наружные волосковые клетки ориентированы в радиальном направлении, а внутренние — в продольном.
Исходя из этих морфологических данных, даются объяснения (Flock, Wersall, 1962; Wersall et al., 1965; Flock, 1965a, 1965b, 1971b) тем физиологическим наблюдениям, о которых говорилось выше. Так, микрофонный потенциал, являющийся отражением активности многих сенсорных элементов и следующий за частотой раздражения в полукружных каналах, представляет собой сумму активностей однонаправленных рецепторов, а в органах боковой линии, где наблюдается удвоение частоты ответа, воспроизводится активность двух противоположно ориентированных групп клеток (рис. 110). При этом предполагается, что амплитуда суммарных ответов противоположного знака не равна. Причина данного обстоятельства пока остается неясной (возможно, что амплитуда деполяризационных ответов превышает величину гиперполяризационных реакций).
Дирекционная чувствительность и рецепторные потенциалы
За последние годы были получены и другие данные, говорящие о дирекционной чувствительности сенсорных волосковых клеток органов чувств. Так, в экспериментах с внутриклеточным отведением потенциалов удалось непосредственно зарегистрировать де- и гиперполяризационные ответы в волосковых клетках боковой линии (рис. 33) (Harris et al., 1970; Flock, 1971a, 1971b; Flock et al., 1973a) и органа слуха (Mulroy et al., 1974; Weiss et al., 1974), закономерно связанные с направлением воздействия на рецепторные элементы. При этом в органах боковой линии можно было зарегистрировать противоположные по характеру ответы рецепторов, ориентированных противоположным образом. Свойства этих ответов хорошо соответствовали тем предположениям, которые ранее высказывались на основании анализа микрофонных потенциалов (рис. 110, сравн. А и Б).
Чувствительность первичных тканевых механорецепторов позвоночных к направлению действия раздражения
Способность этих сенсорных приборов различать направление раздражающего воздействия изучалась в значительно меньшей степени, чем эти же свойства вторичных рецепторных приборов. Здесь прежде всего следует упомянуть об опытах на вибриссах. По-видимому, начальной работой в этом направлении следует считать исследование Фитцджеральда (Fitzgerald, 1940), в котором было показано, что при сгибании вибриссы в одном направлении в нервных волокнах возникает значительный афферентный разряд, а при сгибании в противоположном он практически отсутствует. При этом вибриссы, расположенные на разных участках тела, имеют различное «предпочтительное» направление наклона. В активности одиночных нейронов тригеминального ганглия крысы, связанных с вибриссами, также было отмечено существование дирекционной чувствительности (Zucker, Welker, 1969). Однако морфологический субстрат данного феномена остается невыясненным. Потенциалы отводились в этой работе на большом расстоянии от рецепторов, так что невыясненным остается вопрос, возникают ли при «неактивном» сгибании вибриссы в ее нервных окончаниях гиперполяризационные процессы.
Существование дирекционной чувствительности механорецепторов внутренних органов отмечалось в опытах В. Н. Черниговского и его сотрудников (Черниговский, 1960). В этих экспериментах растяжение стенок полых органов сопровождалось значительной ответной реакцией лишь при определенном направлении раздражающего воздействия. Специально данное явление исследовано не было.
Наиболее полное изучение чувствительности к направлению действия стимула у тканевых рецепторов было проведено в опытах на тельцах Пачини (Ильинский, 1965, 1966г, 1966е, 1969, 1970, 1971; Ильинский, Волкова, 1966; Ильинский и др., 1968; Nishi, Sato, 1968; Ильинский, Волкова и др., 1974).
В отличие от исследований, проведенных на волосковых сенсорных клетках органов чувств, работы по изучению чувствительности механорецепторов к направлению воздействия, рассматриваемые в данном разделе, проводились одновременно с изучением гиперполяризационных процессов.
Указания на существование гиперполяризационных сдвигов потенциала в тельцах Пачини при их раздражении имелись в работах ряда авторов (Alvarez-Buylla, Remolina, 1959; Ильинский, 1963; Ozeki, Sato, 1965). Однако механизм данной гиперполяризации был неясен. Предполагали, что речь идет о вторичной следовой гиперполяризации (Ozeki, Sato, 1965), но детальный анализ (Ильинский, 1965, 1966а, 19666, 1966г) заставил признать, что в тельцах Пачини возникают первичные гиперполяризационные ответы. В этих же исследованиях впервые была описана дирек- ционная чувствительность телец Пачини. Оказалось, что в ее основе лежит способность этих рецепторов генерировать как деполяризационные, так и первичные гйперполяризационные ответы.
Первичные гиперполяризационные рецепторные потенциалы телец Пачини
Эти гиперполяризационные ответы возникали в тельцах Пачини наряду с генерируемыми ими обычными деполяризационными локальными и распространяющимися реакциями (рис. 112). По величине они обычно были меньше деполяризационных РП, хотя в ряде случаев могли достигать весьма большой амплитуды (рис. 65, Б, 7). Блокирование ПД новокаином не влияло
на гиперполяризационные ответы, так же как оно не влияет и на деполяризационные РП (см. стр. 76 и рис. 52). Характер реакции у рецепторов in vitro и in vivo был примерно одинаковым. Декапсулирование телец Пачини изменяло общий рисунок ответной реакции, но гиперполяризационные ответы сохранялись (рис. 112, 1).
Рис.
112. Гиперполяргь/ционные рецепторные
потенциалы телец Пачини при различных
условиях эксперимента (по: Ильинский,
1966а).


Активность семи (1—7) различных рецепторов. Запись стимула: на 2—6 —верхний луч, на 7} а—-б — нижний луч, на z отсутствует. На 1, 2 ПД блокированы новокаином. На 7, а—б — запись ответа одного и того же рецептора при разной скорости нарастания стимула. 1 — декапсулированный, 2 — 7 — интактные рецепторы.
Проведенный анализ (Ильинский, 1965, 1966г, 1966е) позволил заключить, что указанные ответы являются истинными первичными гиперполяризационными РП. Отмечавшееся в опытах многообразие ответов было обусловлено тем обстоятельством, что гиперполяризационные РП возникали лишь при определенном направлении раздражения нервного окончания. Рассмотрение механизма возникновения гиперполяризационных ответов было осуществлено на основе экспериментов с использованием плавно
433
изменяющихся треугольных или синусоидальных стимулов (в этом случае побочные колебательные процессы в капсуле рецепторов практически отсутствовали). В данных опытах было обнаружено, что деполяризация возникает только при действии одного из фронтов механического толчка. Второй же фронт либо не оказывал никакого действия, либо вызывал появление позитивного колебания потенциала — гиперполяризационного рецепторного потенциала (рис. 112, 2, 5, 6) (см* также рис. 24).
Как было ясно из экспериментов по изучению on- и off-ответов в тельцах Пачини (Ильинский, 1966в; см. также стр. 120), нервное окончание рецептора при использовании длительных стимулов деформируется дважды: в момент включения раздражения (действие переднего фронта стимула) и в момент выключения воздействия (влияние заднего фронта стимула). Деформация нервного окончания при выключении раздражения возникала за счет упругих сил капсулы рецептора и происходила в направлении, перпендикулярном направлению действия переднего фронта стимула.
Гипотеза о механизме возникновения деполяризационных и гиперполяризационных РП. Чтобы объяснить, почему различные фронты раздражающего стимула могут вызвать разного рода де- и гиперполяризационные эффекты, была выдвинута гипотеза (Ильинский, 1965, 1966г), основывающаяся на некоторых известных из литературы данных (Hubbard, 1958). Из привлеченных данных следовало, что на область центральной колбы оказывает влияние лишь динамическая составляющая механического стимула, вызывавшая только радиальные перемещения, которые были довольно равномерными вдоль всего хода нервного окончания. Так как нервное окончание в тельцах Пачини имеет вид эллиптического цилиндра (Pease, Quilliam, 1957; Черепнов, 1968а; см. рис. 5, на котором представлено поперечное сечение нервного окончания), было предположено (Hubbard, 1958), что в момент раздражения меняется соотношение между поперечными осями нервного окончания. А это (при неизменном объеме цилиндра) должно сопровождаться изменениями величины поверхности. Увеличение площади поверхностной мембраны нервного окончания и являлось, по мнению Хабберда, фактором, приводящим к возникновению возбуждения в тельцах Пачини.
Ильинский обратил внимание на то обстоятельство, что увеличение площади рецепторной мембраны может иметь место лишь в случае деформации нервного окончания при раздражении, направленном вдоль малой оси его поперечного сечения. При раздражении же тельца Пачини вдоль большой поперечной оси его нервного окончания площадь поверхностной мембраны должна уменьшиться. На этом основании было высказано предположение (Ильинский, 1965), что увеличение площади рецепторной мембраны (растяжение мембраны) приводит к возникновению деполяризации, а уменьшение поверхности мембраны (сжатие мембраны)
Рис. ИЗ. Изменение плшЦади (S) поверхностной мембраны нервного окончания тельца Пачини при изменении соотношения К=а!Ъ между его большой (а) и малой (Ь) полуосями (по: Ильинский, 1966а).
Рис.
114. Дирекционная чувствительность телец
Пачини (по: Ильинский, 1965).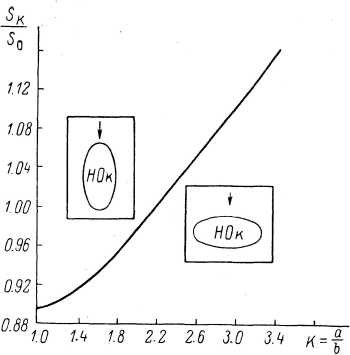


А — соотношение между направлением стимула (стрелки) и положением поперечного сечения нервного окончания (черный эллипс). Слева направо: начальное положение (0°) и после поворотов рецептора на 90 и 180° вокруг продольной оси; черная точка—метка начального положения. В — ответы двух (1, 2) различных рецепторов (на 2 ПД блокированы новокаином). Отметка времени — 4 мсек.; калибровка — на 1 — 10, на 2 — 5 мкв.
приводит к появлению гиперполяризации (рис. ИЗ). Таким образом, механическое раздражение, наносимое вдоль малой оси поперечного сечения нервного окончания, должно вызвать появление деполяризационного РП и спайковой активности, а раздражение, приложенное в направлении большой оси поперечного сечения, должно обусловить возникновение гиперполяризационных ответов. Наконец, гипотеза предсказывала, что ответная реакция должна быть менее четкой в случае, когда механический стимул наносится под каким-либо острым углом к главным осям поперечного сечения нервного окончания.
Эксперименты подтвердили гипотезу. Оказалось, что характер реакции существенно менялся при поворотах тельца Пачини вокруг продольной оси и изменении тем самым направления раздражения по отношению к поперечным осям нервного окончания. При этом ответы в исходном положении рецептора относительно стимулирующего устройства и при вращении рецептора на 180° были практически идентичными. Напротив, при повороте тельца на 90 и 270° ответная реакция претерпевала существенные изменения по сравнению с исходной.
Если в начале исследования тельце расположить таким образом, чтобы спайк возникал практически точно в момент начала раздражения, т. е. когда его латентный период был минимальным, то при повороте рецептора на 90° ПД начинал появляться на вершине треугольного стимула, т. е. скрытый период его возникновения относительно начала раздражения был максимальным. Этого и следовало ожидать, исходя из изложенной гипотезы, ибо появление деполяризационного ответа (деполяризационного РП и ПД) с минимальным скрытым периодом должно было происходить лишь при совпадении направления раздражающего стимула с малой осью поперечного сечения нервного окончания; при повороте же тельца на 90° направление удара совпадало с большой осью, и поэтому ПД в начале раздражения не возникал. Он появлялся лишь в тот момент, когда за счет сил упругости капсулы при снятии раздражения нервное окончание начинало деформироваться по своей малой оси (момент, следующий за максимумом воздействия).
Если первоначально передний фронт стимула, соответствующий нарастанию раздражения, вызывал появление деполяризации, а задний, нисходящий фронт — гиперполяризации, то при повороте рецептора на 90° начало толчка вызывало развитие гиперполяризационных колебаний РП, а снятие раздражения — появ^ ление деполяризации (рис. 114).
Гистологическое исследование телец Пачини, маркированных в конце электрофизиологического эксперимента, показало, что практически во всех случаях характер электрических ответов соответствовал ожидавшемуся положению осей поперечного сечения нервного окончания относительно раздражающего устройства (Ильинский, Волкова, 1966; Ильинский и др., 1968).
Изложенные данные и предложенная гипотеза в дальнейшем были подтверждены в опытах других исследователей (Nishi, Sato, 1968), которые провели изучение дирекционной чувствительности как интактных, так и декапсулированных телец Пачини. В последнем случае, раздражая нервное окончание с помощью двух пьезокристаллов во взаимно перпендикулярных направлениях, указанные авторы также обнаружили появление деполяризации в ответ на действие переднего фронта стимула только тогда, когда он наносился на рецептор вдоль малой поперечной оси нервного окончания. На действие заднего фронта стимула



Рис. 115. Гиперполяризационные и деполяризационные ответы тельца Пачини при включении и выключении раздражения до (Л, Б) и после (В, Г) аппликации тетродотоксина (5« 10~7 г/мл) (по: Nishi, Sato, 1968).
Раздражение в направлении большой поперечной оси нервного окончания; увеличение силы раздражения от А к Б и от В к Г. Гипер- и деполяризационные РП на включение и выключение раздражения соответственно. На В и Г постгиперполяризационные ПД блокированы тетродотоксином. Отметка времени — 5 мсек.; калибровка — 50 мкв.
деполяризация возникала при нанесении раздражения в направлении большой поперечной оси (Nishi, Sato, 1968). Данные исследователи также отметили, что нанесение тетродотоксина (5*10“7 г/мл), полностью блокируя возникновение ПД, не уменьшает гиперполяризационный РП (рис. 115). Величина гиперполяризационного РП увеличивалась при возрастании скорости нанесения раздражения и его амплитуды.
Изменения возбудимости рецепторов. Развитие гиперполяризационных РП сопровождалось изменениями возбудимости телец Пачини (Ильинский, 1966а, 1969, 1970, 1971). В период возникновения гиперполяризационного РП возбудимость нервного окончания снижалась, и для получения деполяризационных ответов требовалась большая величина стимула. Вслед за развитием гиперполяризационной реакции наблюдалось противоположное
изменение возбудимости — ее повышение, которое способствовало возникновению ПД (рис. 116). Появлению ПД всегда предшествовало развитие деполяризационного РП, однако после гиперполяризации критический уровень деполяризации был обычно ниже, чем до начала гиперполяризационного РП, иногда даже весьма значительно (рис. 112, 6, 7), что могло способствовать появлению

Рис. 116. Постгиперполяризационные ответы телец Пачини (по: Ильинский, 1966а).
Активность пяти (I—5) различных рецепторов (на 1—3, 5 — верхний луч, на 4, а—б — нижний луч). Снижение критического уровня деполяризации для генерации ПД после возникновения гиперполяризационных РП. На 4, а, 4, б и 5 — увеличение амплитуды ПД после возникновения гиперполяризационных РП. На 4 от а к б интенсивность стимула нарастает. Отметка времени: на 1—3, 5 — 4, на 4 — 2 мсек.; калибровка — 50 мкв.
и ритмических ответов. Если у рецепторов с пониженной возбудимостью ПД не возникали в начальный момент раздражения, несмотря на отчетливо выраженный деполяризационный РП, то после развития гиперполяризации РП спайковая активность обычно возникала (рис. 116, 1—3). Если же генерировались спайки, амплитуда которых варьировала при изменении силы раздражения, то после появления гиперполяризационных РП величина последующих ПД возрастала, т. е. функциональное состояние регенеративной системы улучшалось (рис. 116, 4—5).
Отмеченные особенности гиперполяризационных реакций не являются какими-то исключительными, свойственными только ответам телец Пачини. Напротив, они представляют собой явления, весьма характерные для самых разных нервных структур, реагирующих на прекращение гиперполяризационного сдвига потенциалов. В литературе имеются данные, говорящие о том, что при определенных условиях снятие анода постоянного тока может вызвать появление возбуждения в различных нервных элементах. Это показано в опытах на одиночных нервных волокнах (Ooyama, Wright, 1959, 1961, 1962; Tasaki, 1959b; Wright, Ooyama, 1961; Ходоров, 1965), мотонейронах спинного мозга (Fukami, 1962), гигантских нервных клетках беспозвоночных (Герасимов и др., 1965), рецепторах ракообразных (Edwards et al., 1963) и т. д.
Гиперполяризация (устранение которой вело к появлению импульсной активности) могла быть обусловлена и влиянием тормозных синапсов (рис. 86) (опыты на рецепторах растяжения — Kuffier, Eyzaguirre, 1955; Эзрохи, 1974; гигантских нейронах беспозвоночных —Таис, 1956), влиянием химических агентов, имитирующих действие тормозных медиаторов (Kerkut, Thomas, 1963) и т. п. В рецепторах ампул Лоренцини также наблю далось возбуждение в момент выключения | тормозного анодиче- ского стимула (Murray, 1962, 1965а; Акоев, Ильинский, 1972, 1973; Акоев, Жадан и др., 1974). Однако следует заметить, что общим условием появления анодоразмыкательного (постгиперпо- ляризационного) возбуждения являлось наличие фоновой деполяризации субстрата. Эта деполяризация могла быть следствием специальных ^экспериментальных воздействий, таких, например, как пропускание деполяризующего тока (Ooyama, Wright, 1961, 1962; Fukami, 1962), повышение концентрации ионов калия в наружной среде (Frankenhaeuser, Widen, 1956; Tasaki, 1959b; Ooyama, Wright, 1961), уменьшение концентрации ионов кальция (Edwards et al., 1963). Она могла быть вызвана ухудшением функционального состояния субстрата (Tasaki, Mi- zuguchi, 1948; Coombs et al., 1955; Frankenhaeuser, Widen, 1956; Герасимов и др., 1965), а также могла быть обусловлена и чисто физиологическими причинами (Kuffier, Eyzaguirre, 1955; Таис, 1956; Murray, 1962, 1965а; Акоев, Ильинский, 1972; Акоев и др., 1974а, 19746).
Анодоразмыкательные ответы, наблюдавшиеся перечисленными авторами, как правило, возникали при более низких величинах потенциала по сравнению с ПД, появлявшимися при раздражении объектов исследования деполяризующими стимулами. Нередко можно было наблюдать появление ритмического разряда при выключении гиперполяризующего толчка тока. Наконец, если до выключения гиперполяризующего воздействия амплитуда ПД была уменьшена в силу тех или иных причин, то первые спайки после выключения поляризации восстанавливались до нормальной величины и лишь потом их амплитуда вновь падала. Точно такие же особенности ответных реакции при исчезновении гиперполяризационных РП были установлены и у телец Пачини (см. выше). В этой связи появилось предположение о наличии фоновой деполяризации нервного окончания в тельцах Пачини и возникла необходимость выяснить ее причины. Было выдвинуто предположение (Ильинский, 1966а, 1969) о существовании вокруг нервного окончания среды с необычным составом (повышенным содержанием калия). Это положение в дальнейшем подвергалось экспериментальной проверке (Ильинский, Красникова, 1972). Оказалось (см. главу 7), что действительно в жидкости телец Пачини содержится повышенное по сравнению с плазмой крови и тканевой жидкостью содержание ионов калия. Наличие повышенной концентрации ионов калия в жидкости телец Пачини, по-видимому, способствует повышению чувствительности этих механорецепторов к действию адекватного раздражения (см. стр. 375).
♦ * ♦
Таким образом, следует признать, что в тельцах Пачини существуют условия для развития истинного постгиперполяризацион- ного возбуждения. Нужно учесть, что тельца Пачини являются одним из основных типов тканевых сенсорных приборов, сигнализирующих о моменте появления (или прекращения) механического раздражения. Поэтому они должны обладать механизмами, обеспечивающими их максимальную чувствительность, а также механизмами, способствующими быстрому восстановлению их исходного функционального состояния. Гиперполяризационные ответы, по-видимому, вносят свой вклад в реализацию этих задач. Изучение гиперполяризационных ответов телец Пачини и явлений постгиперполяризационного возбуждения позволило более глубоко проанализировать природу off-ответов этих рецепторов. Первоначальная гипотеза (Ильинский, 1966в), объясняющая происхождение ответов на выключение, исходила только из механических особенностей капсулы рецепторов, а также из предположения, что деформация нервного окончания этих рецепторов равнозначна в любом направлении. Опыты по дирекционной чувствительности телец Пачини показали, что это не так. Благодаря этим опытам стало возможным понять различные особенности изменений не только латентных периодов, но и порогов возникновения ПД при включении и выключении раздражения. При нанесении раздражения в направлении малой поперечной оси нервного окончания латентный период ответа на включение всегда короче ответа на выключение, однако пороги генерации ПД в ответе на выключении часто могут быть ниже, чем пороги генерации ПД в реакции на включение за счет постгиперполяриза- ционных эффектов (рис. 117). При больших силах воздействия, как это уже отмечалось, могут появляться множественные от-

Рис. 117. Изменение он- и off-ответов телец Пачини при повороте рецептора вокруг продольной оси (Ильинский, 1966а).
На А: 1—5 — направление раздражения совпадает с малой поперечной осью нервного окончания. На Б: 1—з — между направлением раздражения и малой поперечной осью —» острый угол. На В: 1, 2 — между направлением раздражения и малой поперечной осью — прямой угол. На А—В — сила раздражения возрастает от первой записи к последующим. Отметка времени — 4 мсек.; калибровка — 200 мкв.
веты — следствие, прежде всего, колебательных процессов в капсуле рецептора. Они также могут быть результатом и постгипер- поляризационных явлений.
В процессе эволюции рецепторные приборы достигли очень высокой степени совершенства. Это совершенство выражается помимо их высокой чувствительности, надежности, большого диапазона воспринимаемых нагрузок и т. д. также и в удивительной приспособленности всех структур рецепторов к выполнению своей специфической функции. Можно с полным правом утверждать, что практически каждая деталь строения рецептора тем или иным способом вносит свой вклад в его функционирование. Поэтому в настоящее время становится крайне необходимым комплексное морфо-функциональное исследование сенсорных приборов. Следует отметить, что такого рода комплексные работы в самых различных областях исследований механорецепции сейчас становятся скорее правилом, чем исключением (см., например: Skoglund, 1956; Lowenstein, Wersall, 1959; Flock, Wersall, 1962; Wersall et al., 1965; Ильинский, Волкова, 1966; Ильинский и др., 1968; Свидерский, Князева, 1968; Iggo, Muir, 1969; Попов, Светлогорская, 1971; Chambers et al., 1972; Fillenz, Widdicombe, 1972; Burgess et al., 1974, и мн. др.).
Приведенные выше факты, касающиеся дирекционной чувствительности, дали основание утверждать, что форма нервного окончания в тельцах Пачини имеет прямое отношение к механизму деятельности этих механорецепторов (Ильинский, 1965, 1966г; Ильинский, Волкова, 1966). В этой связи были специально измерены размеры нервного окончания в тельцах Пачини [большая (2а) и малая (25) оси поперечного сечения нервного окончания в его средней части у рецепторов правильной формы]. По данным световой микроскопии, эти величины оказались равными: большая ось — 5.48+1.95, малая — 2.56 + 1.01 мкм, их отношение К—а/Ъ —2.26+0.44. По данным электронной микроскопии, 2а=5.73+1.56, 25-2.48+0.77 мкм, #=2.35+0.31 (Ильинский и др., 1968; Ильинский, Волкова и др., 1974; Волкова, Черепнов, 1971).
Указанные данные были сопоставлены с результатами математического анализа изменений формы нервного окончания в тельцах Пачини. При этом исследовался вопрос: каково должно быть соотношение осей (K=alb) нервного окончания постоянного объема для того, чтобы минимальные механические стимулы, приложенные в направлении малой поперечной оси (возбуждающем направлении для телец Пачини), давали бы наибольшую деформацию — относительное увеличение площади поверхности (т. е. максимальный возбуждающий эффект в случае реального нервного окончания). Оказалось (Ильинский и др., 1968; Ильинский, Волкова и др., 1974), что оптимальные характеристики наблюдаются у цилиндра эллиптической формы при К—2.2 (рис. 118, прерывистая линия), т. е. именно при том соотношении осей,
 которые
имеют место в реальных нервных окончаниях.
«Чувствительность» оптимального по
своим характеристикам тела (эллиптического
цилиндра) к пороговым для телец Пачини
стимулам наиболее высока: абсолютное
приращение площади в 7—70 раз превышает
соответствующее приращение площади
поверхности круглого цилиндра того же
объема.
которые
имеют место в реальных нервных окончаниях.
«Чувствительность» оптимального по
своим характеристикам тела (эллиптического
цилиндра) к пороговым для телец Пачини
стимулам наиболее высока: абсолютное
приращение площади в 7—70 раз превышает
соответствующее приращение площади
поверхности круглого цилиндра того же
объема.
Рис. 118. Влияние эллипти ческой формы нервного окончания тельца Пачини на относительное изменение площади его поверхности при деформации (по: Ильинский, Волкова и и др.. 1974).
По оси абсцисс — отношение осей эллипса (К); по оси ординат — относительное приращение площади поверхности нервного окон- чания^.ю<].
Таким образом, можно заключить, что немиелинизированное нервное окончание телец Пачини имеет форму, наиболее приспособленную для восприятия механических воздействий, ведущих к изменению площади его поверхности. Удалось установить, что эллиптическая форма поперечного сечения нервного окончания в тельцах Пачини являлась следствием билатерального строения внутренней колбы и возникала в онтогенезе по мере развития структур капсулы рецепторов (Ильинский, Волкова, 1966; Ильинский и др., 1968; Ильинский, Волкова и др., 1974; Волкова, Черепнов, 1971; Волкова, 19726).
Дирекционная чувствительность других тканевых рецепторов
Кроме телец Пачини, другие тканевые механорецепторы на сегодняшний день не подвергались детальному изучению в отношении их чувствительности к направлению действия механического раздражения, и этот вопрос еще ждет своего исследования. Все же существование у ряда инкапсулированных рецепторов таких черт поразительного сходства в строении с тельцами Пачини, как наличие билатеральной организации внутренней колбы, эллиптическая форма поперечного сечения нервного окончания, наблюдающаяся у некоторых других сенсорных образований (телец Гербста, некоторых «безымянных» инкапсулированных телец, телец Гольджи—Маццони и др.) (Armstrong, Quilliam, 1961а, 1961b; Quilliam, 1963, 1966; Quilliam, Armstrong, 1963; Andres, 1969), дают основание полагать, что механизмы их деятельности, в том числе и относящиеся к дирекцион- ной чувствительности, могут быть очень сходными с теми, которые уже исследованы у телец Пачини (Ильинский, 1966а, 1967).
Вместе с тем имеются экспериментальные данные, как уже отмечалось в главе 3, относящиеся к ряду тканевых механорецепторов, например, кожным рецепторам II типа и суставным рецепторам — сухожильным органам Гольджи и тельцам * Руффини. Согласно полученным данным, эти рецепторы обладают в той или иной степени дирекционной чувствительностью. Фактически чувствительностью к направлению воздействия обладают благодаря своему расположению в мышцах и мышечные веретена, что позволяет проводить те широко используемые в литературе приемы идентификации этих рецепторов, которые рассматривались ранее. У рецепторов мышц также наблюдаются и гиперполяризационные ответы. Однако гиперполяризационные реакции, отмечаемые у мышечных веретен, не связывались с явлениями дирекционной чувствительности.
Гиперполяризационные ответы мышечных веретен. Такого рода ответы впервые были описаны в виде ответа на выключение у мышечных веретен лягушки (Katz, 1950b). В дальнейшем гиперполяризационные сдвиги потенциала при снятии раздражения с мышечных веретен были отмечены и другими авторами, работавшими как на кошках (Lippold et al., 1960а), так и на лягушках (Алексеев, Макаров, 1969; Ottoson, Shepherd, 1971а; Алексеев, 19736). Таким образом, гиперполяризация механорецепторов в этих случаях всегда была вторичной, следовавшей за деполяризационными ответами.
Природа данной гиперполяризации обсуждалась в литературе, однако пока нет единого мнения на этот счет. Отчасти, когда у рецепторов сохраняется импульсная активность, следовая гиперполяризация является, по-видимому, по своей природе посттетанической и обусловлена работой электрогенного натриевого насоса,v который связан с регенеративной системой генерации (Алексеев, Макаров, 1970; Пошина, 1973). (Посттетаническая гиперполяризация подробно рассматривалась для других мышечных рецепторов — рецепторов растяжения у ракообразных — Nakajima, Takahashi, 1966). Однако гиперполяризационные сдвиги потенциала наблюдаются и у рецепторов, ПД которых были заблокированы (Katz, 1950b). Высказывалось мнение (Алексеев, Макаров, 1969, 1970; Алексеев, 1972, 19736), что эти ответы также являются результатом деятельности электрогенного натриевого насоса, который включается в работу в ходе генерации деполяризационного РП и который, следовательно, связан с нерегенеративной системой генерации. В пользу такой точки зрения как будто бы говорят опыты с применением ряда веществ, влияющих на процессы активного транспорта и метаболизм (стр. 307). Однако сейчас еще преждевременно делать какие-либо заключения по этому вопросу. Остается неясным и та- кои существенный вопрос: в какой мере гиперполяризационные ответы мышечных рецепторов, следующие за возникновением деполяризационных РП, можно сопоставлять с явно первичными гиперполяризационными реакциями, отмечаемыми у других механорецепторов?
Чувствительность механорецепторов беспозвоночных к направлению действия раздражения
Механорецепторные сенсиллы. Наличие чувствительности к направлению действия раздражения отмечалось во многих экспериментах на механочувствительных сенсиллах у беспозвоночных (Pumphrey, 1936; Pringle, 1938; Laverack, 1962, 1963; Mellon, 1963; Thurm, 1964a, 1964b, 1965a; Gorner. 1965, и др.). Данное явление хорошо видно, например, у ветрочувствительных волосков — рецепторов, назначение которых со-

Рис. 119. Ориентация ветрочувствительных рецепторов головы у саранчи (по: Smola, 1970).
Карты двух (1р и Ир) рецепторных полей. 1 — щетинки; 2 — волоски длиной менее 150 мкм; 3 — волоски длиной между 150 и 230 мкм; 4 — волоски длиной около 230 и более микрон; 5 — изображение направления изгиба.
стоит в сигнализации о направлении движения воздуха (Haskell, 1958; Свидерский, 1967; Camhi, 1969; Smola, 1970а, и др.). При действии механического стимула на ветрочувствительные сенсиллы рецепторы генерируют максимальный ответ при смещениях стержня волоска каудально; величина ответа уменьшается при боковых воздействиях, и, наконец, реакция отсутствует при смещениях волоска рострально. Следует отметить, что расположение волосков на теле насекомого (например, на голове) таково, что раздражение рецепторов потоками воздуха происходит, по-ви-
димому, как раз в направлении их максимальной чувствительности (Свидерский, 1973). Так как при полете насекомого потоки воздуха образуют завихрения, рецепторы, расположенные в разных точках, раздражаются преимущественно в различных направлениях, что и находит свое отражение в различных особенностях их расположения (рис. 119). Следует заметить, что максимальная чувствительность сенсилл совпадает с плоскостью искривления их стержней, что позволяет легко определить направление их максимальной чувствительности (Camhi, 1969).
Механизм дирекционной чувствительности механорецепторных сенсилл пока еще не выяснен. В основе его, по-видимому, может лежать группа явлений. К ним относятся особенности строения вспомогательного аппарата (асимметричность структуры кутикулярного валика, окружающего волосок, асимметричность структуры самого волоска и наличие у волоскового стержня изгиба), особенности локализации дендрита рецепторной клетки, который располагается асимметрично в основании волоска и др. (см. рис. 19) (Thurm, 1964а, 1964b, 1965а; Camhi, 1969; Свидерский, 1973; Иванов, 1973, и др.). Остается пока неясным, ограничивается ли механизм дирекционной избирательности волосковых рецепторов только перечисленными факторами, среди которых ведущую роль играет строение и свойства вспомогательного аппарата, или же составной его частью являются и какие-либо функциональные особенности самих рецептирующих элементов.
Другие механорецепторные приборы. Чувстви1ельность других механорецепторов беспозвоночных к направлению действия стимула неоднократно отмечалась в литературе. Например, такого рода наблюдения были сделаны при изучении фазных механорецепторов ПД-органа (см. стр. 188), волосковых механорецепторов статоцистов (см. стр. 184) и т. д. Природа этих явлений детально не изучалась.
Дирекционная чувствительность у простейших. В литературе имеются также сведения о чувствительности к направлению механического стимула у простейших, обладающих ресничками (см., например, обзор: Серавин, 1967). В экспериментах на различных инфузориях было показано (Kino- sita et al., 1964; Eckert, 1965, 1972; Naitoh, Eckert, 1969a, 1969b, 1973;Eckert, Naitoh, 1972; Eckert et al., 1972, и др.), что механическое раздражение поверхности организма влияет не только на цилиарную активность, но и на мембранный потенциал. При этом раздражение передней поверхности организма вызывало падение сопротивления мембраны и развитие деполяризации, а раздражение задней поверхности^обусловливало появление гиперполяризации. Авторы именуют эти градуальные ответы механорецеп- торными потенциалами и сопоставляют их с локальными реакциями механорецепторов, в частности телец Пачини.
Исследование у парамеций ионных механизмов указанных ответов на механическое раздражение показало, что первичные де- 446
поляризационные реакции были обусловлены входящими токами, а первичные гиперполяризационные сдвиги потенциала — выходящими токами. В последнем случае наблюдалось увеличение проводимости мембраны к ионам калия и потенциал клетки сдвигался в направлении равновесного калиевого потенциала (Eckert et al., 1972; Naitoh, Eckert, 1973).
В какой мере локальные электрические явления, возникающие при механических воздействиях у простейших, можно сопоставлять с рецепторными потенциалами у механорецепторов, могут показать только дальнейшие эксперименты.
* * *
Представленные материалы дают основание утверждать, что дирекционная чувствительность является важным свойством очень многих механорецепторных приборов. В ряде случаев она возникает благодаря особенностям организации вспомогательного аппарата, в других же случаях — она результат специфического строения и свойств самих рецептирующих элементов. При этом дирекционная чувствительность оказывается связанной с развитием локальной гиперполяризации — гиперполяризационными РП. Свойства последних изучены в настоящее время значительно меньше, чем деполяризационных РП, однако несомненно, что анализ их является необходимым условием расшифровки процессов, протекающих в механорецепторах при действии на них адекватных стимулов.