

Лаааааааааааааааааааааааааалалаааааалллаа| аааааааааааааааааад
Рис. 67. Действие ритмического раздражения на мышечное веретено лягушки (по: Ottoson, Shepherd, 1970с).
Изменение ПД (А, Б) и РП (В) при раздражении рецептора С частотой 150 (А), 300 (Б), 100 (В, а), 200 (В, б) и 400 (В, в) стимулов в 1 сек. Сверху вниз: биоэлектрическая активность; раздражение. На А и Б а—в — последовательные записи. Отметка времени: на А, Б — 25, на В — 100 мсек.; калибровка (только для В) — 1 мв.
экстракапсулярных интрафузальных мышечных волокон), факторы, обусловленные механическими свойствами вспомогательных структур рецептора, определяют лишь 15—20% общего процесса адаптации. Эти же авторы обращают внимание на тот факт, что у самых разных рецепторных приборов, обладающих тоническими свойствами (фото-, хемо-, механо-, электро-, терморецепторов), имеется динамическая фаза развития РП (см. обзоры: Fuortes, 1971; Tamar, 1972), хотя вспомогательные структуры этих разных рецепторов совершенно различны. Динамический компонент реакции наблюдается даже при действии постоянного электрического тока на некоторые нервные волокна (Hodgkin, 1948). Все это позволяет говорить и о возможной роли самих возбудимых мембран (прежде всего рецепторной) в развитии процессов адаптации.
Удаление ионов калия затрудняет развитие динамической составляющей ответа вплоть до ее полного исчезновения. Статический компонент ответа изменяется в меньшей степени. Ионы же натрия в большей степени влияют на статическую часть реакции (Husmark, Ottoson, 1971а, 1971с). На этом основании эти авторы полагают, чю основная часть начальной адаптации мышечного веретена к действию стационарного раздражения имеет в своей основе ионные процессы. Сходные взгляды развивают и другие исследователи (Алексеев, Макаров, 1970; Алексеев, 19736, см. также стр. 311). Вопрос о механизме адаптации мышечного веретена сейчас нельзя считать решенным. Только дальнейшие эксперименты могут показать, какие факторы и в какой степени оказывают влияние на развитие этого процесса.
В экспериментах на мышечных веретенах также исследовалось действие ритмического раздражения разной частоты (см. стр. 163). Было показано (Ottoson, Shepherd, 1970с), что и в этих опытах наблюдается хорошо заметная адаптация (рис. 67), степень которой возрастает при увеличении часюты воздействия. По мере угнетения ответов амплитуда ПД сильно варьировала (рис. 67, А, б). В дальнейшем нормальные ПД генерировались лишь на каждый второй (третий, четвертый и т. д.) стимул (рис. 67, А, в и Б, в). РП в этих случаях сохранялся. Была также отмечена суммация РП (рис. 67, В). В целом общая картина ответа веретена была сходной с той, которая отмечалась в тельцах Пачини (сравн. Loewenstein, 1958). На формировании суммарной реакции веретена, по-видимому, сказываются анатомические (наличие разветвлений сенсорного аксона в интрафузальной мышце) и механические свойства рецептора.
Сравнение адаптационных свойств двух видов рецепторов растяжения у ракообразных
Для изучения процессов адаптации данный тип механорецепторных приборов особенно удобен. Это определяется наличием расположенных рядом МРОХ и МРО2, весьма сходных
морфологически, а основное функциональное различие между ними как раз и заключается в разнице их адаптационных особенностей (рис. 68, 7, 3). Кроме того, как и в мышечном веретене, наличие динамической и статической фаз ответа позволяет исследовать явления адаптации на одном и том же МРО.
Первоначально (Florey, 1956, 1957а) высказывалось предположение о том, что в основе адаптационных свойств МРО лежит особенность самой рецепторной мембраны. Далее была выдвинута точка зрения, согласно которой причиной снижения частоты импульсации в ходе действия стационарного раздражителя являются механические особенности рецепторной мышцы (Krnjevic,
Рис.
68. Сопоставление адаптационных свойств
медленно (МРО^
и быстро (ЛГРО2)
адаптирующихся рецепторов растяжения
(по: Nakajima, Onodera, 1969b).

![]()
![]()


1, 3 — до, 2, 4 — после обработки рецепторов тетродотоксином (2-10-7 г/мл). Сверху вниз: биоэлектрическая активность; растяжение мышцы (шкала справа). Отметка времени — 5 сек.; калибровка —10 мв.
van Gelder, 1961). Наконец, было отмечено, что оба указанных фактора принимают участие в явлениях адаптации, а, кроме того, существенным является и развитие процессов адаптации в аксоне сенсорной клетки (Wendler, Burkhardt, 1961; Wendler, 1963). Весьма детальное рассмотрение явлений адаптации МРО и оценка вклада различных структур этого органа в суммарную картину адаптационного процесса были осуществлены Накаджима и Онодера (Nakajima, 1964; Nakajima, Onodera, 1969а, 1969b). В самое последнее время появились данные, посвященные роли ионных механизмов в явлениях адаптации МРО (Ottoson, 1973; см. также Klie, Wellhoner, 1973).
Как показало сравнительное исследование МРОХ и МРО2 (Nakajima, Onodera, 1969b), характер временного течения деполяризационных РП в них при одинаковом растяжении был одним и тем же: за первоначальной динамической фазой ответа
следовала стационарная фаза, величина которой мало менялась в обоих случаях (рис. 68, <2, 4). Этот результат позволил4утверждать, что за различия в адаптационных способностях между МРОХ и МРО2 ответственна только регенеративная система генерации этих рецепторов, а не их вспомогательные структуры и не их нерегенеративные механизмы. Последний вывод, правда, оспаривается сейчас Оттосоном (Ottoson, 1973), который удалял ионы калия из окружающего рецепторы раствора и обнаружил различные изменения динамической фазы РП МРОХ и МРО2. В быстро адаптирующемся рецепторе динамическая фаза РП полностью пропадала в бескалиевом растворе, в то время как у медленно адаптирующегося рецептора она значительно уменьшалась, но не исчезала совсем.
Рис.
69. Изменение рецепторного потенциала
МРОХ
при раздражении в условиях
стабилизированной длины (Л) или
стабилизированного напряжения (Б)
(по:
Nakajima, Onodera, 1969b).

Сверху вниз: РП; изменение длины (А) или напряжения (Б) мышцы рецептора (шкалы справа). Импульсная активность блокирована тетродотоксином (2-10~7 г/мл). Отметка времени — 500 мсек.; калибровка — 10 мв.
Изучение хода временного изменения деятельности МРОХ показало (Krnjevic, van Gelder, 1961; Wendler, 1963), что в ходе действия раздражения частота импульсации рецептора изменяется параллельно изменению напряжения его мышцы. Это относилось как к изменению активности МРОХ, так и МРО2.
Более тщательное исследование изменения деятельности МРО с регистрацией РП позволило установить, что при стабилизации длины и (или) напряжения уменьшение РП в ходе действия раздражения (адаптация) ослабляется, особенно во втором случае (рис. 69) (Nakajima, Onodera, 1969b). В силу этого адаптацию МРО можно было попытаться описать на основании его вязкоупругих особенностей (сравни данные по мышечному веретену: В. Matthews, 1933; Р. Matthews, 1964, 1972). Однако если в начальном периоде (первая секунда) изменений РП в ходе действия раздражения 71% адаптационного ослабления ответа можно было объяснить простыми вязко-упругими свойствами МРО, то изменения РП за более длительный отрезок времени (6 сек.) лишь на 49% могли быть описаны в рамках такой модели. Это означает существование либо каких-то неучтенных механических свойств МРО, либо наличие совсем иных механизмов, участвующих в изменении РП в ходе действия раздражителя. Дальнейшие эксперименты должны прояснить этот вопрос. Однако уже сейчас имеются данные, говорящие об участии й адаптационных изменениях РП и собственно нерегенеративных механизмов генерации (Ottoson, 1973). Возможно, что они являются к тому же достаточно универсальными, благодаря чему и можно наблюдать у разных рецепторов динамическую и статическую составляющие РП. (См. также данные экспериментов с фиксацией потенциала в соме MPOf. Klie, Wellhoner, 1973).

Рис. 70. Различные аккомодационные свойства аксона МРОХ (по: Nakajima, Onodera, 1969а, схематизировано).
j — аксон MPOf, II — импульсные разряды различных участков аксона —А4) на действие постоянного тока амплитудой около 0.5 мка; Сверху вниз: биоэлектрическая активность; отметка раздражения. Отметка времени — 500 мсек.; калибровка — 5 мв.
Изучение особенностей регенеративной системы МРОХ и МРО2 показало (Nakajima, 1964; Nakajima, Onodera, 1969а), что при электрической стимуляции сенсорного нервного волокна МРОХ отвечает фазно-тоническим, а МРО2 — лишь фазным разрядом. Обнаружилось, что способностью генерировать тонический ответ обладают не все части медленно адаптирующегося нейрона, а только область аксона, находящаяся вблизи сомы] клетки, т. е. тот самый участок, который имеет наименьший порог для генерации спайка. Более удаленные части аксона могли генерировать только фазный разряд при действии постоянного тока (рис. 70). Таким образом, разные участки нервного волокна МРОХ обладают различными аккомодационными свойствами. Причины этого явления пока неясны.
Следует указать, что ионные механизмы ритмической активности при действии постоянного тока на нервные структуры остаются сейчас еще недостаточно выясненными (см.: Hodgkin, 1948; Tomita, Wright, 1965; Grundfest, 1966; Stein, 1967, и др.), а ведь именно расшифровка этого механизма является необходимой для понимания процессов адаптации МРО (действия постоянного генераторного тока на регенеративную систему генерации). Пока лишь удалось выяснить (Nakajima, Onodera, 1969а), что прекращение генерации ПД под действием постоянного тока как в МРОг (под влиянием сильного тока), так и в МРО2 не связано с изменением концентрации ионов калия и натрия снаружи или изнутри от мембраны. Возможные же изменения проницаемости мембраны к этим ионам, а также значение электрогенного насоса, пока остаются невыясненными.
Если суммировать данные всех авторов, относящиеся к проблеме адаптации рецепторов растяжения у ракообразных, то можно заключить, что основными факторами адаптации этих механорецепторов к действию стационарного растяжения являются свойства вспомогательных структур (участие в формировании динамического компонента ответа на этапе возникновения РП) и участие регенеративной системы генерации, ответственной за наличие или отсутствие импульсации, в период статической фазы реакции. Первый механизм адаптации играет более важную роль в случае медленно, а второй — в случае быстро адаптирующихся рецепторов растяжения. Вклад регенеративной системы оценивается для МРО/как 2/7 против 5/7, определяемых другими, более существенными ”в адаптации звеньями системы. Большая часть эффекта относится к более ранним этапам формирования ответной реакции (Nakajima, Onodera, 1969b).
Рассмотренные выше явления адаптации МРО исследовались на рецепторах, изолированных из организма (т. е., если пользоваться терминологией, употребляемой в отношении мышечных веретен, на пассивных МРО). Поведение же активных МРО, т. е. сохранивших нормальные эфферентные связи с вышележащими отделами нервной системы, может существенно отличаться (Спиваченко и др., 1972; Ильинский, Спиваченко, Штирбу, 1974). В частности, это может выражаться в значительном ускорении процесса адаптации. При наличии фоновой активности тормозного эфферентного нейрона МРОХ в ответ на раздражение мог отвечать только фазным разрядом (рис. 94). Рассмотрение этих явлений будет приведено далее в главе 5, где излагаются вопросы эфферентного контроля.
* * *
Имеющиеся в литературе данные по явлениям адаптации, развивающимся в первичных пассивных (в отсутствие эфферентной
вход
Механический
Механический Электрический
стимул стимул
|
СЕНСОРНЫЙ орган
вспомогательные
Рецептирующие структуры ( ' - - - >
стимул
Выход
Фазный
рецептор
II
вход
Тонический
рецептор
Нервное
волокно (регенеративные элементы)
(нерегенеративные)
элементы
О
10 20 мсек.
Выход
рецептор .АЛЛАЛ...
Ж.
Механический
Механический. Электрический
Рис.
71. Схема процессов адаптации в первичных
механорецепторах.




![]()









![]()






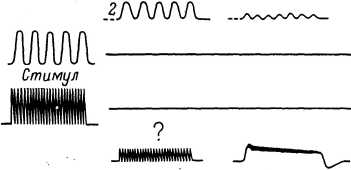
I — действие статической деформации, а — интегральная кривая изменения частоты импульсов; б — запись импульсного разряда. II — действие ритмических раздражений разной частоты. 1 — начало, 2 — конец воздействия.
14 Физиология механорецепторов регуляции) механорецепторах, суммированы в виде схемы на рис. 71. Для того чтобы было ясно, из чего складывается конечный ответ рецептора, регистрируемый на его выходе (рис. 71, справа) при действии раздражающего воздействия (слева), на рисунке показана также раздельная реакция основных составных частей сенсорного прибора при изолированном действии на них стимула такой же амплитуды, формы и длительности, что и действующий на весь рецептор в целом. В случае вспомогательных элементов и нерегенеративной системы генерации таким стимулом является механический раздражитель, а в случае регенеративной системы генерации — электрический. На схеме представлена реакция тонического (медленно адаптирующегося) и фазного (быстро адаптирующегося) механорецепторов (соответственно типа «медленного» рецептора растяжения у ракообразных и тельца Пачини у кошки) в ответ на действие стационарной деформации, а также ответы фазного рецептора (нижняя часть рис. 50, II) на длительное прерывистое воздействие (типа вибраций) различной частоты. Схема показывает, что адаптационные явления, отмечаемые в ответах рецепторов, в различных случаях являются следствием разных причин, обусловленных тем, что составные части сенсорных приборов по-разному участвуют в формировании суммарной реакции.