

АКАДЕМИЯ НАУК СССР
НАУЧНЫЙ СОВЕТ ПО КОМПЛЕКСНЫМ ПРОБЛЕМАМ
ФИЗИОЛОГИИ ЧЕЛОВЕКА И ЖИВОТНЫХ •
РУКОВОДСТВО
ПО ФИЗИОЛОГИИ
ФИЗИОЛОГИЯ
СЕНСОРНЫХ
СИСТЕМ
ЧАСТЬ ТРЕТЬЯ
ФИЗИОЛОГИЯ
МЕХАНО РЕ ЦЕ ПТОРОВ
ИЗДАТЕЛЬСТВО «НАУКА» ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ ЛЕНИНГРАД • 1975
АВТОР ТОМА О. Б. ИЛЬИНСКИЙ
УДК 612.812
Физиология сенсорных систем. Часть третья. Физиология механорецепторов. Ильинский О. Б. В серии «Руководство по физиологии». 1975. Изд-во «Наука», Ленингр. отд-ние, Л. 1—560.
В книге рассматриваются вопросы строения, цитохимии и функции основных видов механорецепторов в коже, внутренних органах, опорно-двигательном аппарате, органах чувств позвоночных и некоторых беспозвоночных. Излагаются особенности рецепторных потенциалов и импульсной активности первичных и вторичных механорецепторных приборов. Затрагивается деятельность некоторых других рецепторов (электрорецепторов и др.), чувствительных к механическим воздействиям. Рассматриваются механизмы адаптации, эфферентной регуляции и дирекционной чувствительности механорецепторов. Подробно излагаются данные по составу и физико-химическим свойствам жидкостей лабиринта. Характеризуются эндолимфатические потенциалы. Рассматривается роль ионов (натрия, калия и др.) и различных веществ (катехоламинов, ацетилхолина и др.) в деятельности механорецепторов. Затрагиваются вопросы развития и регенерации рецепторов. Рассматриваются механизмы действия механического раздражения. Излагаются проблемы морфо-функциональной организации первичных и вторичных механорецепторов. Илл. — 128, табл. — 32, библ. — 2155 назв.
Редакционная коллегия Руководства
50300-685
Ф
055(02)-75 бз-7114-75
© Издательство «Наука», 1975
ВВЕДЕНИЕ
Живые организмы не могут существовать, не получая информации о состоянии внешней и внутренней среды и об изменениях, происходящих в них. Чем более быстро и точно организм получает такую информацию, тем выше, при прочих равных условиях, его шансы в борьбе за существование. Поэтому в ходе эволюции животные организмы выработали различные экстеро- и интероцепторы — специальные образования, предназначенные для трансформации энергии раздражения в специфическую активность нервной системы, в сигналы, несущие нервным центрам информацию о действующем агенте. Одни из рецепторов оказались приспособленными для преобразования в нервный импульс лишь одного вида раздражения (светового, механического, температурного и т. д.), другие же — нескольких (механического и температурного, механического и химического и т. п.).
Несмотря на то что в животном мире существует огромное разнообразие рецепторных аппаратов, исследователям удалось, особенно за последнюю четверть века, добиться больших успехов в изучении функционирования этих структур. Многие факты, имеющие первостепенное значение для понимания общих принципов деятельности рецепторов, были получены в опытах на одиночных механорецептррах позвоночных и беспозвоночных животных.
Механорецепторам уделяется сейчас большое внимание. Различные аспекты их строения и функций обсуждаются в многочисленных публикациях. Однако в мировой литературе до настоящего времени отсутствует специальное руководство, посвященное рассмотрению физиологии механорецепторов, хотя многие важные стороны работы механорецепторов и даже отдельные виды рецепторов, например мышечные веретена, и являлись предметом обширных и превосходных сводок. В предыдущих томах руководства, относящихся к сенсорной физиологии, подробно рассматривалась деятельность рецепторных структур вестибуляр-
механорецепторы. Изучение механорецепторов, по-видимому, может быть полезным для понимания деятельности других рецепторных структур еще и потому, что во многих из них предполагается существование первичных механо-химических явлений. Наконец, не следует забывать и тот факт, что рецепторы вообще и механорецепторы в частности представляют собой очень удобные объекты для изучения многих проблем общей нейрофизиологии.
Успехи, достигнутые в изучении механорецепции, не должны скрыть, что наши знания по этому вопросу еще явно недостаточны. Многие проблемы физиологии механорецепции далеки от своего разрешения. Это, очевидно, проявится в данном томе как естественная неполнота освещения ряда вопросов. Это же оказало и известное влияние на распределение материала в книге. Так, недостаточность изучения механизмов первичных трансформационных процессов, протекающих в механорецепторах при их раздражении, не позволила начать изложение именно с этих вопросов, что, возможно, было бы более оправдано.
Поэтому изложение начинается с традиционного подхода, с анализа структуры многочисленных механорецепторных приборов. При этом достаточно четко выявляется, что механорецепторы позвоночных, как и многие другие рецепторные приборы, могут быть подразделены на две группы: первичночувствующие (первичные) и вторичночувствующие (вторичные). У первичных рецепторов действие адекватного стимула осуществляется непосредственно на окончания сенсорного нейрона, который таким образом «первично» встречается с раздражителем. В сенсорных приборах второго типа между окончаниями сенсорного нейрона и точкой приложения адекватного стимула располагается специализированная, рецептирующая клетка. Под влиянием раздражителя эта клетка может изменять свое функциональное состояние, что, в свою очередь, оказывает при участии медиатора определенное воздействие на сенсорный нейрон. Последний, следовательно, испытывает на себе действие стимула опосредованно, «вторично». Все механорецепторы органов чувств позвоночных являются вторичными сенсорными приборами. К первичным же относятся механорецепторы в теле позвоночных (тканевые рецепторы): рецепторы опорно-двигательного аппарата, кожи, внутренних органов, сердечно-сосудистой системы. У беспозвоночных животных все механорецепторы являются первичными. Однако их можно подразделить также на две основные структурные группы. В основе такого деления лежит наличие у беспозвоночных двух типов сенсорных нейронов. Сенсорные нейроны I типа имеют один не- разветвленный дендрит, который у высших беспозвоночных обычно заканчивается в весьма специализированных образованиях. Сенсорные нейроны II типа имеют, как правило, разветвляющиеся дендриты, которые оканчиваются непосредственно в тканях.
В первичном рецепторе может быть выявлено два вида электрической активности; локадьцые ответы, так называемые ре* приторные потенциалы (РП), и распространяющиеся ответы, или штчщиалы действия (ПД). У вторичных рецепторов, помимо этих ответов, имеет место феномен синаптического проведения, и частности появление постсинаптических потенциалов, возникающих на постсинаптической мембране окончаний сенсорного noiipona. Изучение рецепторных потенциалов, подробно рассматриваемых в главе 2, представляет большой интерес в связи с анализом первичных процессов, протекающих в сенсорных приборах, но является по методическим причинам весьма сложным делом. РП в настоящее время исследованы У сравнительно небольшого числа рецепторов. Среди механорецепторов к ним относятся прежде всего тельца Пачини, мышечные веретена позвоночных и рецепторы растяжения ракообразных. В отличие от этого сведения о деятельности основной массы механорецепторов почерпнуты почти исключительно из экспериментов с регистрацией импульсной активности, рассмотрение которых составляет содержание главы 3.
При анализе ответов рецептора на действие адекватного раздражения, например ответов на выключение, т. е. ответов на длительную стимуляцию, процессов адаптации (глава 4) и т. д., так же как и ответов на действие различных других физических и химических раздражающих факторов среды (глава 6), важно выяснить, какая часть сенсорного прибора (вспомогательные структуры, элементы, ответственные за возникновение рецепторных и пиковых потенциалов), в какой степени и каким образом участвует в формировании ответной реакции. В тексте показано, что в деятельности механорецепторов очень большое значение имеют механические свойства их вспомогательных структур, а также окружающих тканей. В главе 6 рассмотрено также влияние некоторых факторов среды на рецепторы, которые наряду с механической чувствительностью обладают способностью воспринимать действие и других видов стимулов, например электрического тока.
Рецепторы могут испытывать на себе различные воздействия со стороны вышележащих отделов нервной системы, а также многообразные влияния соседних рецепторных приборов. Наиболее тонкое и эффективное регулирование наблюдается в более совершенных и сложных механорецепторных приборах: органе слуха, вестибулярном аппарате, сенсорных структурах мышц. Эти вопросы, и прежде всего эфферентная регуляция, будут рассматриваться в главе 5.
Как показывают исследования, выполненные в последние 10— 15₽лет, в деятельности сенсорных приборов, и прежде всего механорецепторов, очень большое значение имеют факторы той среды, которая окружает собственно рецептирующие элементы. К таким факторам, подробно рассматриваемым в главе 7, относится, например, содержание различных ионов в жидкостях, омывающих рецепторы, наличие особых потенциалов (например, эндолимфатических потенциалов) в структурах, связанных с рецепторами и т. д. Учитывая, что в литературе практически отсутствуют специальные сводки, касающиеся регулирования деятельности различных механорецепторных структур, а также рассматривающие факторы среды, в которых функционируют эти сенсорные приборы, данные вопросы излагаются в книге более детально.
В литературе существует большое количество фактов, относящихся к изучению метаболизма и содержания биологически активных веществ в механорецепторах. Эти данные излагаются в главе 8. Далее рассматриваются вопросы, относящиеся к проблеме образования рецепторных приборов. Исследование причин, влияющих на развитие сенсорных структур и становление их специфичности, является важным для всей проблемы механорецепции. Это определило выделение данного материала в специальную главу (глава 9).
Наряду с деполяризационными рецепторными потенциалами механорецепторы способны генерировать и гиперполяризационные ответы. Эти ответы в настоящее время изучены в значительно меньшей степени, чем деполяризационные реакции. У механорецепторов изучение гиперполяризационных реакций часто проводится одновременно с исследованием способности оценивать направление раздражающего воздействия. Это объясняется тем обстоятельством, что во многих случаях в основе чувствительности рецепторов к направлению действия раздражителя} лежит развитие локальной гиперполяризации, угнетающейJ импульсную активность. Все эти вопросы рассматриваются в^главе 10.
Исследование явлений гиперполяризации и чувствительности механорецепторов к направлению действия раздражения позволило выяснить ряд важных факторов, относящихся к механизму действия механического раздражения. Изложению этих и других данных, касающихся механизмов механорецепции, и их современной трактовке посвящена глава 11.
Наконец, в последней главе книги формулируются некоторые общие принципы функционирования механорецепторов позвоночных и беспозвоночных животных, насколько это было возможно в рамках резюмирования материала предыдущих разделов текста.
Большое разнообразие механорецепторных структур и в связи с этим крайнее обилие самых различных сведений об их строении и деятельности не позволило даже в рамках специального руководства охватить все имеющиеся в литературе факты. Представлялось целесообразным остановиться преимущественно на данных, характеризующих наиболее важные стороны функциональной организации механорецепторов. Эти данные в настоящее время получены на некоторых наиболее доступных и «удобных» для экспериментирования, а посему и наиболее изученных образованиях, таких как тельца Пачини, мышечные веретена, ре-
цечггоры растяжения ракообразных. Поэтому их строение и деятельность рассматриваются более подробно. Однако следует укапать, что даже в пределах материалов об основных рассматриваемых видах механорецепторов требовалось провести отбор наибо- лсо существенных и достоверных фактов.
Необходимость объединения очень большого и разрозненного фактического материала поставила перед автором трудную задачу. Поэтому автор не претендует на исчерпывающую полноту изложения по всем затронутым вопросам и будет благодарен читателям за деловые критические замечания, касающиеся как построения книги, так и рассматриваемого фактического материала.
♦ * *
Во многие разделы книги включены экспериментальные данные, полученные на протяжении ряда лет в Институте физиологии им. И. П. Павлова АН СССР вначале в Лаборатории общей физиологии, руководимой акад. В. Н. Черниговским, а затем в Лаборатории общей физиологии рецепции. Все мое научное становление проходило под влиянием идей В. Н. Черниговского и иод его руководством. Развиваемые в книге общие трактовки полученных фактов родились из основных концепций, выдвинутых и разрабатываемых В. Н. Черниговским. За все это я глубоко благодарен Владимиру Николаевичу.
Я признателен всем сотрудникам Лаборатории общей физиологии рецепции, работы которых представлены в настоящем руководстве и помощь которых была полезна на различных этанах работы над этой книгой. Я особенно благодарен канд. биол. паук Т. Л. Красниковой, совместно с которой была написана глава 7, и канд. мед. наук Н. И. Чалисовой, участвовавшей в написании главы 9. Кроме того, я глубоко признателен им, а также Г. В. Макухиной за помощь при завершении работы над книгой. Я выражаю мою самую искреннюю признательность всем сотрудникам Лаборатории общей физиологии рецепции, принимавшим участие в подготовке рукописи к печати, й особенно Э. А. Васильевен! за участие в оформлении этой книги. Обращаясь со словами благодарности к учителю, названным и неназванным моим сотрудникам и друзьям, считаю своим долгом сказать, что всегда сохраню глубокую признательность моему отцу и самую благодарную память о моей матери. Без участия всех этих людей выполнение данной работы едва ли было бы возможным.
Глава 1 структура механорецепторов
Строение механорецепторных приборов удивительно разнообразно. Это связано в значительной степени с тем, что они окружены самыми различными структурами тела, которые в той или иной степени участвуют в образовании рецепторного прибора, прежде всего вспомогательного аппарата. A priori можно утверждать, что кожные механорецепторы млекопитающих и кутикулярные механорецепторы насекомых должны значительно отличаться по своей структуре, несмотря на то что все они располагаются на внешней поверхности тела животных. Кроме того, функциональное назначение механорецепторов очень многообразно: одни механорецепторы предназначены для восприятия перемещений воздуха или воды, другие — перемещения крови или пищевых масс, третьи — перемещения частей тела, четвертые — действия сил земного тяготения и т. д. Такое разнообразие видов механических стимулов, воспринимаемых механорецепторами, также не может не сказаться на их строении. Далее, часть рецепторов приспособлена для регистрации быстро изменяющихся событий, другая — стационарных состояний и т. д. И все это получает соответствующее^отражение в структуре воспринимающих приборов.
В настоящее время одной из широко распространенных классификаций механорецепторов является их подразделение по месту расположения. Естественным является также их подразделение на рецепторные приборы, локализованные непосредственно в тканях тела и в органах чувств. Целесообразно также раздельно рассматривать структуру механорецепторов позвоночных и беспозвоночных животных. Все эти подходы используются в данном изложении.
При рассмотрении структуры механорецепторов следует учесть одно важное обстоятельство. Механорецепторы, как и все рецепторные приборы, состоят из нескольких основных частей: 1) вспомогательного аппарата рецептора с окружающими его структурами; 2) собственно рецептирующих структур, ответственных за восприятие энергии специфического стимула (ими могут быть кнк окончания сенсорного нейрона, так и специализированные, рг цеп тирующие клетки); 3) структур, обеспечивающих возникновение и распространение импульсной активности (ими являются элементы сенсорного нейрона). Основные различия в строении ризных механорецепторных приборов состоят в особенностях структуры их вспомогательного аппарата, а также в наличии пли отсутствии у них специализированных рецептирующих клеток1. С позиций физиологии второе обстоятельство является особенно существенным (Davis, 1961; Ильинский, 1972а). Поэтому ип эту сторону дела при изложении материала обращено особое ннимание.
МЕХАНОРЕЦЕПТОРЫ ПОЗВОНОЧНЫХ
Рецепторные окончания кожи
Различные аспекты морфологии нервных окончаний в коже, а также соотношения их структуры и функции суммированы в ряде обстоятельных сводок (Weddell et al., 1955; Zo I terman, 1959; Weddell, 1960; Winkelmann, 1960b, 1960c; Melzack, Wall, 1962; Gauna, 1966, 1968, 1973, 1974; Hensel, 1966, 1973; Iggo, 1966a, 1968, 1974; Malinovsky, 1967a, 1967b, 1968; Sinclair, 1967; Iggo, Muir, 1969; Есаков, Дмитриева, 1971; Munger, 1971; Ильинский, 19726; Andres, 1973; Andres, During, 1973; Burgess, Perl, 1973). В данном разделе книги мы касаемся лишь тех вопросов строения кожных рецепторов, рассмотрение которых позволит в дальнейшем лучше понять механизмы деятельности этих образований.
В коже находится большое количество разнообразных нервных окончаний, распределенных весьма неравномерно. Их особенно много в пальцах рук, ладонях, подошвах, губах, половых органах, что и обеспечивает этим областям повышенную чувствительность (Weddell et al., 1955; Winkelmann, 1960b, 1960c). Основными видами нервных окончаний в волосистой коже (90% всей кожной поверхности) позвоночных животных и человека являются свободные нервные окончания, диски (тельца) Меркеля, нервные окончания вокруг волосяных сумок. В коже, лишенной волосяного покрова, основными рецепторными приборами являются свободные нервные окончания, диски Меркеля, тельца Мейсснера п Пачини.
Свободные нервные окончания. Они являются наиболее распространенным видом кожных рецепторов и связаны главным образом с немиелинизированными волокнами, составляющими примерно 80% кожных афферентов. Со свободными окончаниями связаны также тонкие и средние мякотные волокна, диаметр которых обычно менее 6 мкм. Следует заметить, что хотя исследователи и по сей день употребляют термин «свободные нервные окончания», строго говоря, окончания не являются таковыми (рис. 1), ибо они в той или иной степени окружены оболочкой из шванновских клеток и обязательно основной мембраной (Пор- тугалов, 1955; Сайпа, 1968, 1969, 1973). Вместе с тем несомненно и то, что из всех видов афферентных нервных окончаний они более всего свободны от вспомогательных структур.
В дерме на участках, покрытых толстым эпидермисом и имеющих высокие сосочки, основная масса свободных нервных окон- чаний находится в сосочках. Там же, где слой эпидермиса невелик и его граница с дермой не очень извитая, нервные волокна образуют горизонтальные плексиформные разветвления, расположенные в поверхностных структурах дермы (Сайпа, 1966). В эпидермисе большинство свободных нервных окончаний располагается в нижних слоях, однако в некоторых местах, например в пальцах рук, они наблюдаются в большом количестве и в зернистом слое. В роговом слое окончания постепенно истончаются и атрофируются. Непрерывный регенеративный рост (вместе с эпидермисом) интрадермальных нервных волокон обеспечивает восстановление рецепторов. Свободные нервные окончания часто сильно ветвятся, в результате чего одно нервное волокно может иннервировать весьма большую площадь: например, в роговице эта область может достигать примерно 0.5 см2. Области иннервации отдельных нервных волокон обычно значительно перекрывают друг друга.
Диски Меркеля. Считается, что диски образуются в нижней части эпидермиса за счет свободных нервных окончаний совместно с модифицированными эпителиальными структурами. Их точное строение описано рядом авторов (Саппа, 1962; Munger, 1965; Andres, 1966, 1973; К. R. Smith, 1967, 1970; Iggo, Muir, 1969; обзор: Winkelmann, Breathnach, 1973). Они наиболее часто встречаются в области пальцев рук, где иногда могут быть связаны мякотными нервными волокнами с тельцами Мейсснера. Безмякотные волокна, отходящие от нескольких рецепторов Меркеля, могут объединяться в одно толстое миелинизированное волокно, образуя таким образом единую сенсорную структуру довольно сложного строения {тактильная корпускула, touch corpuscle, — см.: Iggo, 1963, 1966а; Iggo, Muir, 1969; Merkel cell— neurite complex, — Andres, 1966; Munger, 1966, 1971).
Эти структуры (рис. 2) встречаются в волосистой коже и представляют собой образования диаметром 100—500 мкм, возвышающиеся над поверхностью кожи. Часто они закрывают выход больших сторожевых волосков, но могут располагаться и вне их. Ядро такого тельца образует плотная коллагеновая ткань. Рецепторные окончания (тельца Меркеля) локализуются непосредственно под базальной мембраной утолщенного и модифицированного эпидермиса. В одной корпускуле может находиться 30—50 телец Меркеля. Одно мякотное волокно (его диаметр в среднем равен 9.5 мкм) может быть связано с 2—3 тактильными
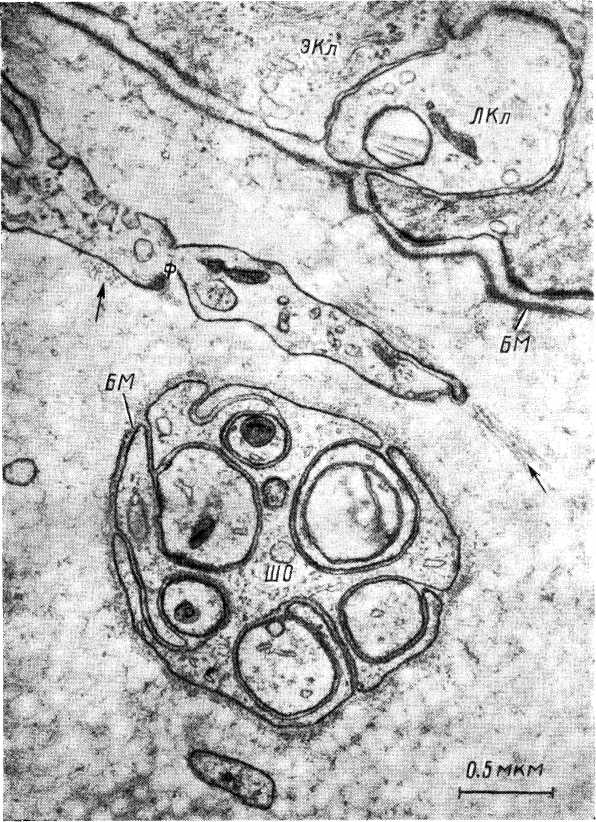
Рис. 1. Горизонтальное нервное окончание в коже крысы (по: Саппа, 1969).
ШО — шванновская оболочка, заключающая в себе 6 аксонов нервных окончаний, 4 из которых содержат митохондрии различных размеров и плотности; БМ — базальные мембраны нервных окончаний и эпидермиса; ЭКл — основная эпидермальная клетка (кератиноцит); ЛКл — отросток клетки Лангерганса; Ф — отросток фиброцита, сопровождаемый пучком микротрубочек, видимых на продольном и поперечном сечении {стрелки).
Рис.
5. Электронномикроскопическая картина
тельца Пачини (по: Ильинский, Волкова
и др., 1974).
I
—
поперечный срез рецептора; II
—
пластины наружной капсулы; а
—
участок пластины с везикулами. НОк
—
нервное окончание; М
—
митохондрии; Щ
—
щель, разделяющая колбу; П
—
пластины; В
—
везикулы.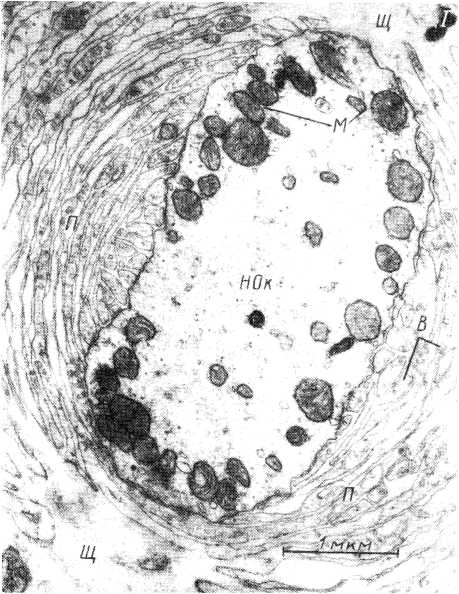

3
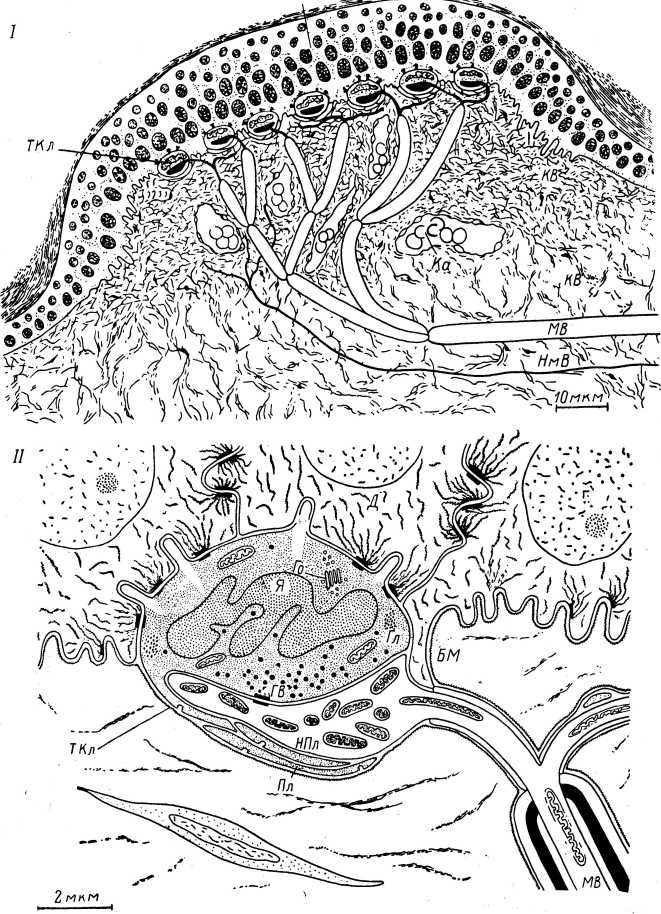
Рис. 2. Тактильное тельце по данным световой и электронной микроскопии (по: Iggo, Muir, 1969).
1 — общий вид тельца; II — тактильная клетка (ТКл) и связанная с ней нервная пла? стинка (НПл). МВ — миелинизированный аксон; НмВ — немиелинизированное волокно» В — эпидермис тактильного тельца; КВ — тонкие и толстые пучки коллагеновых волокон; Ка — капилляр; БМ — базальная мембрана; Д — десмосома и ее выросты; ГВ — гранулярные везикулы; Го — аппарат Гольджи; Гл — гликоген; Пл — пластинки, лежащие в основании нервного окончания; Я — многолопастное ядро.
корпускулами. В корпускулу могут проникать и тонкие дополнительные немиелинизированные волокна. В основании корпускулы имеется обширное скопление капилляров (Iggo, 1963а; Tapper, 1964; Brown, Iggo, 1967; Iggo, Muir, 1969).
Основу воспринимающей структуры тактильной корпускулы образует дископодобно расширяющееся окончание афферентного волокна с тесно примыкающей к нему специальной клеткой Меркеля. Нервное окончание содержит большое количество митохондрий. Клетка Меркеля характеризуется пальцеобразными выростами, проникающими в окружающие структуры. В цитоплазме клетки содержится большое о количество гранулированных везикул диаметром 800—1000 А. Клетки Меркеля имеют десмосомные, а также синапсоподобные контакты с нервным окончанием. Вблизи этих контактов наблюдается скопление гранулированных везикул. Однако типичные синаптические везикулы, которые могли бы рассматриваться как структуры, содержащие медиатор, среди них отсутствуют (Andres, 1973; Winkel- mann, Breathnach, 1973). Вопрос о природе этих контактов пока остается открытым.
Несомненно, что диски Меркеля в настоящее время являются единственной структурой среди тканевых механорецепторов, у которых на основании морфологических данных можно было бы предполагать наличие специализированных рецептирующих клеток. Существует мнение, что клетки Меркеля, которые, по-видимому, проникают в эпидермис во время внутриутробного развития, являются всего лишь своего рода вспомогательной структурой, способствующей медленной адаптации рецепторного прибора (Winkelmann, Breathnach, 1973).
Нервные окончания вокруг волос. Известно, что в волосяном покрове содержатся волоски нескольких видов: обычные, сторожевые (в том числе и большие сторожевые волоски, фолликулы которых окружены венозным синусом — tylotrich follicle — Straile, 1960) и чувствительные волоски — вибриссы (синусные волосы, sinus hairs). Наиболее сложно устроены вибриссы, которые отсутствуют у человека, но встречаются у многих приматов и других млекопитающих. Они обычно локализуются на щеках и вокруг ротового отверстия, но могут встречаться и в других местах.
Волосяные фолликулы достаточно богато иннервированы, и поэтому волоски являются высокочувствительными механо- рецепторными образованиями кожи (Weddell et al., 1955; Winkelmann, 1960b; Забусов, Маслов, 19616; Iggo, 1966a; Munger, 1971; Andres, During, 1973). Нервные окончания, заложенные в обычных волосяных фолликулах, очень многочисленны и представляют собой сеть тонких немиелинизированных нервных волокон, плотно оплетающих структуры волосяной сумки. Рецепторы волосяного мешочка связаны с мякотными нервными* волокнами, большинство из которых имеет диаметр более 6 мкм.
Каждый волос иннервируется несколькими волокнами. В свою очередь одно и то же волокно может иннервировать большое количество волос. Так, в ухе кролика одно волокно может иннервировать около 300 волосяных фолликулов (1 см2). Таким образом, зоны иннервации отдельных волокон широко перекрывают друг друга. Каждое немиелинизированное волокно в волосяной сумке окружено шванновскими клетками, которые как бы сплющивают нервное окончание, отгораживая его от окружающих образований. Нервные окончания содержат большое количество митохондрий, микровезикул и гранул, по-видимому, гликогена (Сайпа, 1969). Шванновские клетки содержат значительное количество пиноцитозных пузырьков (Munger, 1971).
Следует отметить, что в литературе везикулярные образования в клетках часто называют пиноцитозными без достаточных на то оснований. Как известно, пиноцитоз — это явление, связанное с захватыванием клеткой материала из окружающей среды. Следовательно, пиноцитозные везикулы содержат материал внешней среды, поглощенный клеткой. Наряду с такими везикулами в клетках встречаются пузырьки, содержащие продукты обычного клеточного метаболизма, подлежащие выделению вовне. Понятно, что такие везикулы не являются пиноцитозными. Так как процесс их выделения обратен процессу пиноцитоза (эндоцитоза}., его называют обратным пиноцитозом (экзоцитозом). Поэтому везикулы, предназначенные для выделения из клетки, должны называться обратнопиноцитозными (или экзо- 1щтозными) (De Dave, 1969). Однако далеко не всегда удается отличить пиноцитозные везикулы от обратнопиноцитозных. В связи с этим в морфологической литературе часто все везикулярные образования, не содержащие медиатор, т. е. не являющиеся синаптическими пузырьками, называют пиноцитозными. Таким способом синаптические везикулы отграничивают от несинаптических, хотя такое употребление термина «пиноцитозные везикулы», строго говоря, нельзя признать правильным. Заметим, что синаптические везикулы, конечно, тоже являются результатом клеточного метаболизма, однако очень специализированного.
.Вибриссы имеют обычно длинный и тонкий волос, выходящий из фолликула, обладающего рядом характерных морфологических свойств и прежде всего наличием кровяного синуса, который окружает наружную корешковую оболочку. Благодаря существованию синуса, вибриссы и получили наименование синусных волосков. Вибриссы обладают иннервацией, в целом аналогичной, но еще более богатой, чем обычные волоски (Tretjakoff, 1911; Vincent, 1913; Andres, 1966). Свыше 100 волокон могут проникать внутрь' капсулы и образовывать внутри нее свои разветвления. У вибриссы можно обнаружить тельца Меркеля, Гольджи—Мац- цони, различные ланцетовидные окончания, окончания немиели- низированных волокон (Andres, During, 1973).
Тельца Мейсснера. В коже, лишенной волосяного покрова, обнаруживаются тельца Мейсснера, а также сходные с ними образования (Саппа, 1966). Они располагаются в сосочковом слое дермы и локализуются преимущественно в пальцах рук и ног, ладонях, подошвах, губах, языке, половых органах, сосках. Тельца имеют конусообразную или овальную форму (90—120 мкм в длину). Их длинная ось располагается перпендикулярно поверхности кожи. Снаружи тельце Мейсснера окружено тонкой соединительнотканной капсулой, тесно связанной с окружающими тканями. Собственно тельце образуется большим количеством пластинчатых клеток, между которыми ветвятся нервные волокна, идущие параллельно поверхности кожи. Одиночный рецептор может иннервироваться несколькими (до 9, обычно 2—6) толстыми мякотными волокнами, которые после вхождения в тельце теряют миелин (Саппа, 1956, 1966). В свою очередь одно и то же волокно может разветвляться и иннервировать несколько телец. Иногда помимо толстых нервных волокон к рецептору подходят и тонкие волокна, которые рассматриваются некоторыми исследователями (Догель, 1903; Pease, Pallie, 1959) как эфферентные структуры. Нервные окончания в тельце содержат большое количество митохондрий и везикул различного размера. Обилие митохондрий является вообще характерной чертой самых различных рецепторов, говорящей об очень высоком уровне обменных процессов в этих структурах. Вместе с тем интересно отметить, что кровеносные сосуды, обеспечивающие приток питательных веществ, никогда не проникают в тельца Мейсснера (Pease, Pallie, 1959; Hashimoto, 1973).
В пластинчатых клетках телец Мейсснера встречаются неизвестной природы везикулы, расположенные обычно вдоль поверхностной мембраны. Первоначально (Саппа, Ross, 1960; Саппа, 1962) их пытались трактовать как пузырьки, содержащие медиатор, а места контактов пластинчатых клеток, содержащих везикулы, с мембраной нервных окончаний как синапсы, тем более что в отдельных случаях удавалось наблюдать утолщение (как бы постсинаптическое) мембраны нервного волокна в местах прилегания пластин. Однако в дальнейшем более детальное исследование показало, что везикулы в пластинчатых клетках отнюдь не сконцентрированы в области контакта с нервным окончанием. Они встречаются и на больших расстояниях от зоны контакта и имеются все основания рассматривать их как пиноцитозные (Chonchkov, 1973а; Hashimoto, 1973), что привело также и Цавна к отказу от первоначальной точки зрения (Саппа, 1966, 1968; см. также дискуссию на симпозиумах по тканевым рецепторам и по рецепторам кожи: Саппа, 1966, стр. 128; Саппа, 1968, стр. 29). Видимые же утолщения мембраны нервногсг окончания, которые иногда наблюдаются у инкапсулированных рецепторов, легко объясняются наличием небольшой гофрированности поверхностной мембраны. В результате этого на срезах, проходящих косо, в отдельных участках мембрана будет выглядеть как бы расширенной (Черепнов, 1969). Вместе с тем несомненно нельзя исключить также и реального утолщения и уплотнения мембраны нервного окончания в местах, где она испытывает максимальные деформирующие усилия, передаваемые пластинами капсулы. Возможно, что утолщения мембраны нервного окончания, которые
были обнаружены и в тельцах Меркеля (Andres, 1966; Iggo, Muir, 1969; Andres, During, 1973), также обусловлены тем, что именно в этих точках наблюдается максимальная деформация нервного окончания.
Следует заметить, что обнаруженные соединения между нейрональными и ненервными элементами в рецепторах кожи очень сходны^с теми соединениями, которые имеют место^между шван-
Рис.
3. Неокрашенное тельце Начини, изолированное
из брыжейки кишечника кошки (по:
Ильинский, 1966а).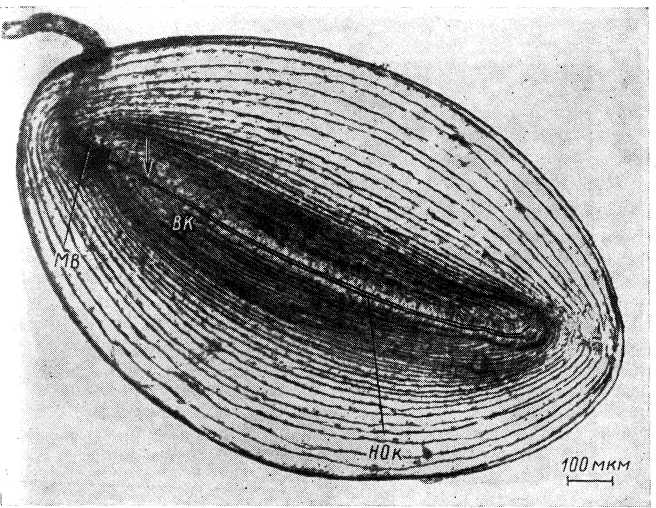

МВ — миелинизированное нервное волокно; НОк — нервное окончание; ВЯ — структура внутренней колбы. Стрелка — примерное начало миелинизированной части нервного окончания. ’
невскими клетками и проводниковыми частями нервных волокон (Сайпа, 1968).
Тельца Пачини. Этим инкапсулированным окончаниям в коже посвящено большое количество работ (обзор основной литературы см.: Pease, Quilliam, 1957; Ильинский, 1966а; Quilliam, 1966; Черепнов, 1968а, 1969; Nishi et al., 1969, 1970; Волкова, Черепнов, 1971; Волкова, 1972а). Типичные тельца Пачини и многочисленные их разновидности относятся к числу наиболее распространенных в организме специализированных тканевых ре-
17
цепторов. Поэтому следует остановиться на строении этих рецепторов подробно, тем более что детали их структуры необходимы для понимания ряда важных особенностей их деятельности. Сведения, полученные в опытах на этих рецепторах, составляют немалую часть материалов о механизмах деятельности механорецепторов в целом.
Тельца Пачини представляют собой наиболее крупные (у человека размер их может достигать 4—5 мм в длину и 1—2 мм в ширину) и одни из наиболее дифференцированных рецепторных образований в тканях. Они локализуются примерно в тех же областях, что и тельца Мейсснера, но располагаются глубже, в нижних слоях дермы и в подкожной жировой клетчатке. Они состоят из многослойной наружной капсулы, внутренней колбы и не миелинизированного нервного окончания, связанного с мякотным нервным волокном (рис. 3). Пространство между пластинами капсулы заполнено ликвором.
Наружная капсула телец Пачини образована 30—40 концентрически организованными пластинами, расстояния между которыми уменьшаются по мере приближения к внутренней колбе. У телец правильной формы изменение расстояния между пластинами может носить весьма закономерный характер (Hubbard, 1958).
Каждый слой наружной капсулы образован несколькими плоскими клетками толщиной порядка 0.2 мкм. Края клеток плотно соприкасаются, образуя непрерывную пластину. Слои поддерживаются значительным количеством коллагеновых волокон и базальными мембранами. Соединений между соседними слоями относительно немного. Их значительно больше лишь во внутренних 5—10 слоях (в зоне роста), являющихся переходной областью к внутренней колбе (Pease, Quilliam, 1957). На поверхности тельца несколько пластин, тесно связанных друг с другом, образуют единую наружную оболочку всей капсулы рецептора. В ней отсутствуют какие-либо отверстия, что делает ее практически непроницаемой даже для ионов (Gray, Sato, 1955; Ильинский, Красникова, 1972).
Внутренняя колба в тельцах Пачини образована тесно прилегающими друг к другу пластинами (их около 60), которые разделены узкой щелью на две равные симметричные части (Schumacher, 1911; Пилат, 1924). В центре пластинчатые структуры вплотную примыкают к нервному волокну, которое на поперечном срезе имеет вид эллипса (Pease, Quilliam, 1957). Хорошо заметная щель, разделяющая внутреннюю колбу, точно совпадает с направлением большой поперечной оси нервного окончания (рис. 4).
Тельца Пачини иннервируются толстыми мякотными волокнами диаметром от 3 до 13 мкм (в среднем 5—7 мкм), а изредка и больше. К каждому тельцу подходит одно волокно, которое обычно в месте вхождения во внутреннюю колбу теряет миелин, сужается, а затем вновь увеличивается в размерах (Ильинский, Волкова и др., 1974). Было показано (Quilliam, Sato, 1955), что внутри наружной капсулы миелиновое нервное волокно имеет обычно 1—2 перехвата Ранвье, хотя эти отношения могут быть и более вариабельными (Волкова, 1972а). Мякотное нервное волокно внутри наружной капсулы имеет весьма извитой ход, а его
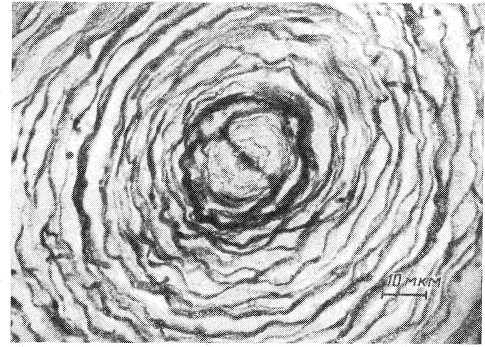
Рис. 4. Микрофотография поперечного среза в средней части тельца Пачини (по: Ильинский, Волкова, 1966).
окраска по Кампосу.
протяженность колеблется довольно значительно — от 50 до 500 мкм и более. По мере удаления от немиелинизированной части нервного окончания и далее от самого тельца расстояния между перехватами Ранвье возрастают.
Немиелинизированное нервное окончание, как правило, не ветвится, а у телец правильной формы, которые чаще встречаются в брыжейке, чем в подкожных тканях, оно идет прямо в середине внутренней колбы, вплоть до ее дистального конца, где оканчивается утолщением или дихотомическим разветвлением. В отдельных случаях нервное волокно проходит через тельце, покрывается опять миелином, входит в следующее тельце, где вновь утрачивает мякотную оболочку.
В области внутренней колбы нервное волокно имеет форму эллиптического цилиндра (рис. 5, /), поперечные размеры которого у телец правильной формы довольно постоянны на всем его протяжении. Специально проведенные измерения показали, что большая поперечная ось нервного окончания составляет в среднем 5.5—5.7 мкм, а малая поперечная ось — 2.5—2.6 мкм (Ильинский и др., 1968).
У телец с необычным строением внутренней колбы может наблюдаться и иная форма нервного окончания. У рецепторов среднего размера длина немиелинизированной части равняется примерно 500—700 мкм.
По последним данным (Spencer, Schaumburg, 1973), нервное окончание телец Пачини может образовывать пальцеобразные отростки, располагающиеся среди структур внутренней колбы. В проксимальной и средней части нервного окончания таких отростков немного. Число их увеличивается в дистальном отделе.
Аксоплазма немиелинизированного нервного волокна содержит большое количество митохондрий, число которых резко возрастает по мере приближения к дистальной части окончания: в самом конце внутренней колбы нервное окончание может быть буквально забито митохондриями. В средней части нервного окончания они располагаются преимущественно около поверхностной мембраны, часто вплотную прилегая к ней. В одних случаях митохондрии равномерно распределяются по всей поверхности нервного окончания, иногда же они скапливаются в области щели. В этой же области могут встречаться пузырьки с электроннопрозрачным содержимым (Pease, Quilliam, 1957; Quilliam, 1966; Миркин и др., 1967; Черепнов, 1968а, 1969). Электронная плотность нервной мембраны неравномерна. Это объясняется, в частности, тем, что поверхность волокна не идеально гладкая, а как бы слегка гофрированная. Однако важно отметить, что не наблюдается никакого совпадения участков утолщенной мембраны с теми структурами внутренней колбы, которые^могли бы рассматриваться как пресинаптические.
При изучении тонкого строения телец Пачини в [элементах внутренней колбы были описаны (Черепнов, 1968а) везикулы, которые в некоторых клетках скапливались в большом количестве, образуя своего рода везикулярные пакеты. Пластинчатые клетки с везикулами были довольно равномерно распределены по всей внутренней колбе, и какой-либо концентрации их около нервного окончания обнаружено не было. Встречаются они и в элементах наружной капсулы (рис. 5, II). Это не позволяет отнести данные везикулы к разряду синаптических пузырьков. Синаптических контактов между элементами внутренней колбы и нервным окончанием обнаружено не было (рис. 5, II). При вибра- ционнных воздействиях на изолированные рецепторы количество везикулярных образований возрастало (Черепнов, 19686, 1969). В ряде мест можно было обнаружить выход везикул в межклеточное пространство (рис. 5, II). По-видимому, данные везикулы могут рассматриваться как результат метаболической деятельности элементов внутренней колбы. Тем не менее эти структуры заслуживают обсуждения в связи с проблемой эфферентной иннервации телец Пачини.
Морфологами давно (Тимофеев, 1896) были описаны тонкие немиелинизированные волокна, подходящие к тельцам Пачини
(и колбам Краузе) и образующие внутри них сложные разветвления — так называемый аппарат Тимофеева, Существование дополнительных волокон было описано и в других инкапсулированных окончаниях: тельцах Гербста, Грандри, Мейсснера, дисках Меркеля (Догель, 1903; Синицын, 1916; Плечкова, 1947; Сайпа, 1956). Еще Догель рассматривал такого типа волокна как симпатические. В пользу этой же точки зрения высказывалась и Юрьева (1927). Наличие немиелинизированных эфферентных волокон в инкапсулированных рецепторах было продемонстрировано и с помощью электронного микроскопа (у телец Гербста — Andres, 1969; Nafstad, Andersen, 1970; у телец Гольджи— Маццони — Andres, 1969; у аннуло-спиральных окончаний мышечных веретен — Andres, During, 1973; см. также: Banker, Girvin, 1971; Santini, Ibata, 1971).
Другие исследователи оспаривали симпатическую природу дополнительных волокон в инкапсулированных рецепторах и отстаивали их соматическое происхождение и сенсорную функцию (Плечкова, 1947; Колосов, 1953; Назарова, 1953; Португалов, 1955; Забусов, Маслов, 1963). Недавно появились новые работы (Santini, 1968, 1969; Santini et al., 1971), в которых авторам с помощью электронного микроскопа и метода флуоресцентной гистохимии как будто бы удалось показать наличие во внутренней колбе телец Пачини тонких немиелинизированных нервных волокон адренергической природы. Адренергические нервные волокна были описаны и в колбах Краузе (Patrizi, Munger, 1965).
В пользу симпатической природы тонкого дополнительного волокна в тельцах Пачини говорят и другие данные (Goto, Loewenstein, 1961; Loewenstein et al., 1962). Указанные авторы показали наличие эфферентной иннервации телец Пачини как с помощью морфологического, так и физиологического методов. Эффект раздражения выявленного ими тонкого С-волокна был аналогичен действию адреналина на тельца Пачини. Эти авторы, а также и другие исследователи (Ильинский, 1966а; Santini, 1969) отмечают, что в брыжейке кишечника кошки тельца Пачини весьма редко содержат дополнительные тонкие волокна, в то время как в области половых органов такая дополнительная иннервация рецепторов, по-видимому, встречается значительно чаще (Тимофеев, 1896), что, возможно, объясняется различной функциональной ролью указанных рецепторов в разных частях тела. На отсутствие адренергических волокон в тельцах Пачини указывал ряд авторов (Fuxe, Nilsson, 1965; Ghouchkov, 1968; Улумбеков, 1971).
Если резюмировать сказанное, то следует признать, что тельца Пачини, равно как и другие инкапсулированные рецепторы, по-видимому, могут иметь дополнительную эфферентную иннервацию. Возможно, что эти эфферентные волокна происходят из нескольких источников, в том числе и симпатических. В свете указанных фактов везикулярные пакеты, обнаруженные в клеточных элементах внутренней колбы, могут рассматриваться как структуры, связанные с эфферентной иннервацией рецепторов, хотя, конечно, данный вопрос требует еще дальнейшего изучения (сравн.: Spencer, Schaumburg, 1973). Влияние со стороны симпатической нервной системы могут испытывать не только инкапсулированные механорецепторы (Loewenstein, 1956а; Дмитриева, Есаков, 1969). Морфологический субстрат для такого рода влияний был описан (напр.: Fuxe, Nilsson, 1965).
Определенный интерес представляет вопрос о кровоснабжении телец Пачини. Как показали исследования ряда авторов (Schumacher, 1911; Пилат, 1924; Sheehan, 1933; Отелин, 1953, 1965; Pease, Quilliam, 1957; Diamond et al., 1958b; Черепнов, 1969; Nishi et al., 1970; Волкова, 1972a), кровоснабжение рецепторов может осуществляться весьма различно. В одних тельцах имеется довольно сложная сеть капилляров, а в другие рецепторы капилляры вообще не заходят. Внутри тельца капилляры чаще заканчиваются, не доходя до внутренней колбы, в области перехвата Ранвье, либо разветвляются в наружной колбе. Во внутренней колбе капилляры обнаружены не были. Если капилляры оказываются вблизи внутренней колбы, то они располагаются против щели, разделяющей последнюю на две части.
Другие инкапсулированные рецепторы. В подсосочковом слое дермы можно обнаружить концевые колбы Краузе. Они представляют собой сферические, окруженные соединительнотканной капсулой образования, внутри которых нервные волокна образуют сложную систему переплетений (Ormea, Goglia, 1969; Chou- chkov, 1973b). В соединительной ткани в области половых органов встречаются различные варианты так называемых генитальных телец (Polacek, Malinovsky, 1971; Malinovsky, Sommerova, 1973). В глубине дермы встречаются также образования, полу- чившие^названия телец Руффини и Гольджи—Маццони. Тельца Руффини представляют собой веретенообразные структуры, окруженные капсулой (рис. 6). Сложно переплетенные коллагеновые волокна составляют основу рецептора. Они тесно связаны как с коллагеновыми пучками окружающих тканей, так и с многочисленными разветвлениями афферентного нервного волокна (Chambers et al., 1972). Одно основное волокно (диаметр 5—10 мкм) связано с несколькими кустиковидными образованиями. Внутрь капсулы рецептора в ряде случаев может проникать дополнительное тимофеевское волокно. Внутрикапсулярное пространство достаточно велико и заполнено жидкостью неизвестного состава. Тельца Гольджи—Маццони очень похожи на тельца Пачини, однако имеют меньшие размеры (электронномикроскопическое строение этих рецепторов см.: Andres, During, 1973). Некоторые авторы рассматривают тельца Руффини и Гольджи—Маццони не как самостоятельные типы рецепторов, а как различные разновидности колб Краузе и телец Пачини. Вариабельность инкапсулированных рецепторов в коже обсуждалась в литературе (Ма-
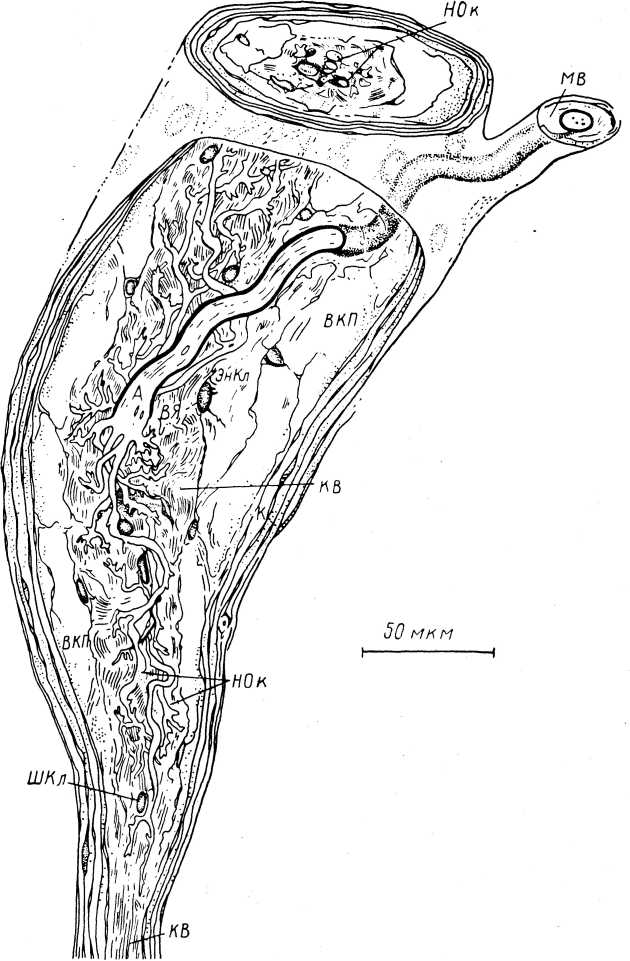
Рис. 6. Реконструкция на основании серийных срезов тельца Руффини из волосистой кожи кошки (по: Chambers et al., 1972).
А — аксон; К — капсула; ВНП — внутрикапсулярное пространство; ЭнКл — эндо- невральная клетка; В Я — внутреннее ядро; НВ — коллагеновые волокна; МВ — миелинизированное волокно с периневральной оболочкой; ШКл — шванновская клетка; НОк — разветвленные нервные окончания.
linovsky, 1966a, 1966b, 1966c). По-видимому, на основе чисто морфологических принципов вообще трудно удовлетворительно классифицировать инкапсулированные нервные окончания (Weddell et al., 1955).
* # *
Таким образом, следует заключить, что в коже имеет место большое разнообразие рецепторов. Наиболее крайними формами являются свободные нервные окончания и инкапсулированные рецепторы типа телец Пачини. Однако между ними существует большое число промежуточных форм. Это не позволяет провести какую-либо четкую границу между различными видами сенсорных структур на основе чисто морфологических критериев. Основное структурное различие между разными рецепторами заключается в строении их вспомогательного аппарата. В структуре рецепторов кожи в настоящее время не обнаружено образований, которые позволили бы рассматривать их как вторичные сенсорные структуры. Напротив, имеются основания считать кожные рецепторы первичными чувствительными приборами.
Рецепторные окончания сердечно-сосудистой системы и внутренних органов
Вопрос о строении интероцепторов, обладающих чувствительностью к действию механических стимулов, является недостаточно ясным, несмотря на обилие исследований, посвященных морфологии чувствительных аппаратов внутренних органов (см., например, сводки: Лаврентьев, 1943, 1947; Иванов, 1945; Смирнов, 1945; Григорьева, 1954; Колосов, 1954, 1958; Куприянов, 1955, 1959; Гилинский, 1958; Хабарова, 1961; Лашков, 1963; Милохин, 1963; Семенов, 1965; Швалев, 1965; Хайсман, 1966; Brettschneider, 1966; Kadanoff, 1966; Abraham, 1969; Fillenz, Widdicombe, 1972). Дело в том, что рецепторные приборы, локализованные в сердечно-сосудистой системе и внутренних органах, могут воспринимать раздражители разной природы (Черниговский, 1943в, 1960, 1967). При этом провести точную морфо-физиологическую идентификацию механорецепторных аппаратов оказывается достаточно трудно. Поэтому лишь сравнительно небольшое число изученных морфологами интероцепторных приборов может рассматриваться как бесспорно механочувствительные.I
К интересующей нас группе прежде всего следует отнести инкапсулированные рецепторы специфического строения, механоре- цепторная функция которых не вызывает сомнений, а также не- инкапсулированные рецепторы, локализованные в определенных рефлексогенных зонах.
Инкапсулированные механорецепторы. Тельца Пачини и па- чиниподобные рецепторы встречаются в брыжейке кишечника, в области поджелудочной железы, мочевом пузыре, уретре, по ходу сосудов и т. д. Строение этих рецепторов было рассмотрено выше.
Неинкапсулированные рецепторы. К ним следует отнести нервные окончания, локализованные в специальных рефлексогенных областях, избирательная механочувствительная функция которых показана с большой убедительностью (например, аортальная рефлексогенная зона или область каротидного синуса). С известной долей вероятности к ним могут быть отнесены и некоторые рецепторы ряда полых органов, механическое раздражение которых является основным в ходе их нормальной деятельности (рецепторы мочевого пузыря, легких, сердца). На деталях строения всех этих рецепторов у нас нет возможности останавливаться. Они хорошо рассмотрены в приведенных выше источниках. Здесь мы коснемся лишь некоторых структур этого вида.
Следует отметить, что немиелинизированные интероцепторы могут локализоваться в тканях как довольно диффузно, так и весьма концентрированно. В последнем случае совместно с окружающими их тканевыми элементами они могут даже образовывать как бы своего рода специализированную рецепторную структуру. Примером такой структуры может служить барорецепторный участок дуги аорты.
Нервные структуры дуги аорты подвергались систематическому изучению в работах ряда исследователей (Nonidez, 1935а, 1935b, 1937а, 1937b, 1941; Григорьева, 1954; Хайсман, 1966; Abraham, 1969). Барорецепторная область дуги аорты у птиц и большинства млекопитающих представляет собой достаточно четко очерченный участок неправильной округлой формы, расположенный на вентральной или краниовентральной поверхности сосуда, вблизи от заращенного устья боталлова протока. Величина его у собаки достигает примерно 1.0 X 1.5 см и не меняется существенно в ходе постнатального развития. Собственно барорецепторное образование, в структурах которого располагаются нервные окончания, имеет вид плоско-выпуклой линзы, обращенной своей выпуклой частью к t. media. Толщина его варьирует у разных животных от 200 до 2000 мкм. Основу барорецепторного участка образуют элементы адвентициальной оболочки, которые внедряются в структуры t. media. При этом они претерпевают существенные изменения, благодаря появлению значительного количества коЛлаге- ново-эластических элементов, образующих слои, связанные между собой в единое соединительнотканное образование. По мнению Хайсмана (1966), оно имеет свою систему гладкой мускулатуры, независимую от мышечных структур собственно стенки аорты. А это при наличии мощной эфферентной иннервации данной области позволяет предполагать возможность независимой регуляции деятельности барорецепторов аорты со стороны нервной системы.II
Барорецепторное образование получает значительную васкуляризацию и содержит в себе разветвленную сеть капилляров, чем оно отличается от других участков стенки аорты (Хайсман, 1966).
Нервные окончания барорецепторной области можно подразделить на два типа (Nonidez, 1935а, 1935b, 1937а, 1937Ь, 1941). Один тип — окончания более толстых мякотных волокон — представлен различными бляшками, кустиками, утолщениями, колечками. Другой тип — окончания более тонких миелинизированных волокон — представлен в виде диффузных разветвлений. Кустиковидные окончания являются доминирующими среди барорецепторных структур. В нервных окончаниях содержится большое количество митохондрий (Хайсман, 1966).
Все афферентные нервные волокна аортальной барорецепторной зоны у млекопитающих относятся к системе левого аортального нерва. С правым же аортальным нервом связаны аналогичные барорецепторы области плече-головного артериального ствола (Boss, Green, 1956; Bianconi, Green, 1959). Интересной особенностью нервных волокон, а также претерминальных и терминальных разветвлений является неравномерность их диаметров. Колебания диаметров по ходу нервных волокон могут быть весьма значительными: у собак, например, от 12—15 мкм в местах расширения до 2—3 мкм в местах сужений (Хайсман, 1966). Хорошо миелинизированные участки волокон сменяются участками с плохо развитой (или даже отсутствующей) мякотной оболочкой. Отражением такого рода явлений может служить различие в скоростях проведения по ходу волокон блуждающего нерва (Paintai, 1962).
Нервные окончания барорецепторной области аорты весьма сходны с аналогичными чувствительными элементами в других местах сосудистого русла, и в частности каротидного синуса. Исследования каротидного синуса (De Castro, 1926, 1927, 1929; Смирнов, 1944; Abraham, 1969) показали существование в нем чрезвычайно богатой сети нервных окончаний, которые де Кастро подразделил на два основных типа. Тип I характеризовался диффузной арборизацией, захватывающей значительные пространства. Тип II занимал значительно меньшую площадь ц имел клубкообразный вид. По другим данным (Abraham, 1969), существует много разновидностей нервных окончаний в каротидном синусе, и в целом они сходны с теми, которые встречаются и в других сосудах.
Рецепторы каротидного синуса располагаются среди коллагеновых структур адвентиции, в ее более глубоких слоях. Каких бы то ни было специализированных клеточных элементов, которым можно было бы приписать функцию первичного восприятия раздражения, как в хеморецепторных структурах каротидного клубочка, обнаружено не было.
Многие морфологи при описании структуры рецепторных приборов а и утренних органов и сердечно-сосудистой системы большое внимание уделяют рассмотрению так называемых специальных клеток, которые обнаруживаются вокруг нервных окончаний (Лаврентьев, 1943, 1944; Плечкова, 1947; Григорьева, 1954; Лашков, 1963, и мн. др.). Указанные клетки вблизи барорецепторов сосудов описаны рядом авторов (Nonidez, 1935, 1937а, 1937Ь; Григорьева, 1954; Хайсман, 1966). Отсутствие или наличие такого рода специальных клеток являлось основанием для подразделения тканевых рецепторов на свободные и несвободные (Лаврентьев, 1943; Миславский, Маслов, 1958). Однако, как показывают данные электронной микроскопии, свободных нервных окончаний в организме, по-видимому, нет, ибо все они в той или иной степени окружены шванновскими клетками и основной мембраной (Санна, 1969).
Специальным клеткам вблизи тканевых рецепторов морфологи и гисто- х имики часто приписывают функциональную роль в трансформации энергии внешнего сигнала. По мнению отдельных исследователей, именно эти клетки обеспечивают преобразование механической энергии, а затем в свою очередь действуют па собственно нервные окончания и вызывают возникновение в них возбуждения (Миславский, Маслов, 1958). Указанная точка зрения неоднократно высказывалась морфологами и гистохимиками (Лаврентьев, 1943; Нортугалов, 1955; Koelle, 1961, 1962, 1963, и др.), однако можно утверждать, что никаких серьезных физиологических, равно как и морфологических данных в ее пользу в настоящее время в литературе не имеется. Напротив, имеются все основания считать, что практически у всех тканевых механорецепторов восприятие раздражения осуществляется именно в самих нервных окончаниях. Окружающие же их элементы выполняют вспомогательную, хотя, быть может, и очень важную роль в деятельности механорецепторов. Подробнее этот аспект проблемы будет рассмотрен далее (стр. 388).
В легких были описаны афферентные нервные окончания, расположенные среди гладких мышц, находящихся в стенках воздухоносных путей (Hayashi, 1937; Honjin, 1956; Лашков, 1963; Fisher, 1964; Spencer, Leof, 1964; Fillenz, Widdicombe, 1972). (’ большой долей вероятности их можно рассматривать как меха- порецепторные (легочные рецепторы растяжения), Первоначально нх даже именовали «гладкомышечные веретена» (Larsell, 1922), однако, по своему строению они сходны с другими механорецепторами, расположенными в дуге аорты, каротидном синусе, камерах сердца (Hayashi, 1937).
В других органах также можно обнаружить большое количество нервных окончаний, весьма похожих на рассмотренные выше механорецепторы. Однако их функция не может бытн сегодня установлена достаточно точно.
♦ ♦ ♦
Анализ морфологической литературы позволяет заключить, что в тканях внутренних органов и сердечно-сосудистой системы можно выделить два основных вида механорецепторов: инкапсулированные и неинкапсулированные. Типичными представителями этих рецепторов являются тельца Пачини и свободные нервные окончания. При этом в литературе нет достаточно убедительных морфологических фактов, отвечающих современным требованиям, которые позволили бы провести резкую грань между этими крайними вариантами. Напротив, между ними имеется большое количество промежуточных форм, различающихся между собой только лишь степенью развития вспомогательных структур (см., например: Quilliam, Armstrong, 1963).
Рецепторные образования опорно-двигательного аппарата
Нервные окончания опорно-двигательного аппарата очень разнообразны и весьма многочисленны. Они локализуются в мышцах, сухожилиях, сухожильных влагалищах, межкостных мембранах, фасциях, тканях суставов, надкостнице и т. д. Здесь можно обнаружить рецепторные образования, встречающиеся и в других частях тела (свободные нервные окончания, инкапсулированные рецепторы, такие как тельца Руффини, тельца Пачини или их разновидности), а также и специализированные сенсорные приборы, присущие только данным тканевым образованиям. Такими наиболее специфическими рецепторами опорнодвигательного аппарата являются сухожильные органы Гольджи и мышечные веретена.
Свободные нервные окончания. Наиболее распространенным видом рецепторов опорно-двигательного аппарата являются свободные нервные окончания. Так, например, даже в деэфференти- рованном и симпатэктомированном мышечном нерве (n. m. tibialis post.) после дегенерации перерезанных нервных проводников число немиелинизированных волокон, которые все заканчиваются свободными нервными окончаниями, в 2 раза превышает число мякотных волокон (Stacey, 1969). При этом следует учесть, что и тонкие миелинизированные волокна (диаметром менее 5—6 мкм) также в основном завершаются свободными нервными окончаниями. Последние сильно ветвятся так, что одиночное волокно может иннервировать весьма значительную площадь (до ЗООх Х1000 MKM)’ (Stacey, 1969).
Свободнее нервные окончания располагаются в самых различных структурах опорно-двигательного аппарата и функция их, по-видимому, может быть достаточно разнообразной. Несомненно, что части из но связана с симпатической нервной системой и со*
й
держит катехоламины. Другие же волокна, напротив, сохраняются после симпатэктомии.
Инкапсулированные рецепторы. Из типичных механорецепторов, встречающихся в коже и других тканях организма, наиболее часто в элементах опорно-двигательного аппарата наблюдаются тельца Пачини и пачиниподобные тельца. Пачиниподобные тельца в несколько раз меньше обычных телец, имеют более вытянутую форму и снабжены более тонкими мякотными волокнами (Barker, 1967; Stacey, 1969). При этом одно более толстое волокно (7— 10 мкм) может ветвиться и снабжать целую группу телец волокнами меньшего диаметра. Пачиниподобные тельца наиболее часто встречаются в местах мышечно-сухожильных соединений, в тканях суставов. При локализации в суставах их размер (280X Х120 мкм) больше, чем при расположении в мышцах (200 X хЗО мкм), и связаны они с нервными волокнами несколько больших диаметров (Freeman, Wyke, 1967). По некоторым данным (Skoglund, 1956), диаметр волокон, связанных с модифицированными тельцами Пачини сустава, не превышает 13 мкм.
В тканях суставов встречаются также инкапсулированные рецепторные образования, которые рассматриваются рядом авторов как тельца Руффини. Их строение напоминает структуры сухожильного органа Гольджи и аналогичные нервные окончания в коже (рис. 6), однако в последнем случае размеры телец Руффини меньше, особенно в длину (100 x 40 мкм).
Высказывалось мнение (Polacek, 1966), которое в целом разделяют и другие авторы (Skoglund, 1973), о том, что все суставные рецепторы на основе их структуры, филогенетического развития и функции следует разделить на три типа: 1) свободные нервные окончания; 2) кустиковидные окончания (spray type) и 3) инкапсулированные тельца (типа телец Пачини). К кустиковидным окончаниям относятся руффиниподобные тельца (в тканях суставов) и сухожильные органы Гольджи (в связках). Это подразделение по существу очень близко к другой классификации нервных окончаний в структурах пассивной локомоции (Stilwell, 1957а).
Изучение состава суставных нервов показало, что он сходен г таковыми у кожных проводников (мякотные волокна разных диаметров; немиелинизированные волокна, как симпатического происхождения, так и С-волокна дорзальных корешков). Что касается мякотных волокон, то максимум распределения их диаметров лежит в области 5—8 мкм, волокна же наибольших диаметров равны 17—18 мкм (Gardner, 1944; Skoglund, 1956; Burgess, (’dark, 1969a). Между отдельными животными имеются значительные индивидуальные различия.
Сухожильные органы Гольджи. Они располагаются в сухо- жилиях обычно на границе мышечной и сухожильной тканей, в опорных участках капсул суставов, в^наружных и внутренних суставных связках (Gol^i? 1880). Сухожцдьцьде органы представ* ляют собой основные рецепторные нервные окончания сухожилий, имеющие очень большое значение при осуществлении двигательных актов и поэтому они неоднократно исследовались большим числом исследователей, хотя, как замечает автор, впервые положивший начало электронномикроскопическому изучению этих структур (Merrillees, 1962), все принципиальные результаты с помощью световой микроскопии были осуществлены еще до 1900 г. Обзор исследований по иннервации сухожилий и суставов, а также подробное описание рецепторов в этих образованиях дан в ряде работ (Huber, DeWitt, 1900; Gardner, 1944, 1950; Boyd, 1954; Skoglund, 1956, 1973; Stilwell, 1957аД1957Ь; Ralston et a].,

Рис. 7. Сухожильный орган Гольджи (по: Ramon у Cajal, 1909).
а — сухожилия; б — большое афферентное волокно; в — его чувствительные терминали; г — мышечные волокна.
I960; Merrillees, 1962; Polacek, 1966; Barker, 1967; Freeman, Wyke, 1967; Bridgman, 1968).
Сухожильные органы описаны у всех групп позвоночных. Сложность их строения возрастает в ходе эволюции. Это выражается в развитии капсулы, которой снабжены рецепторы у птиц и млекопитающих, увеличении количества разветвлений основного нервного волокна, оплетающих элементы сухожильной ткани, и т. д.
Сухожильные органы представляют собой структуру вытянутой веретенообразной или иногда цилиндрической формы, размер которой в длину у человека может достигнуть 1 мм, у кошек он меньше — 0.1—0.5 мм (рис. 7). К рецептору подходит одиночное толстое мякотное волокно (8—15 мкм в диаметре), а также более тонкое миелинизированное волоконце, назначение которого неизвестно (Ruffini, 1897; Wohlfart, Henriksson, 1960). Некоторые исследователи полагают, что эти волоконца при участии свободных нервных окончаний, оплетающих рецептор, могут обеспечивать появление болевого ощущения при значительных деформациях (Weddell, Harpman, 1940). Однако возможно, что они представляют собой образования,{сходные по функции с аналогичными структурами телец Пачщш И других инкапсулированных рецец- торов, назначение которых, по-видимому, иное (стр. 21). Основное волокно распадается вблизи концевого органа на ряд вето- чек, которые в свою очередь образуют многочисленные кустиковидные разветвления, оплетающие сухожильные волокна. Сухожильные волокна, проникающие в капсулу рецептора, мельче и не так плотно упакованы в пучки, как снаружи. Нервные окончания обычно отделены от сухожильных волокон цитоплазмой шванновских клеток и материалом основной мембраны и лишь в отдельных местах прямо контактируют с ними. Капсула рецептора покрыта соединительной тканью и элементами оболочки нервного волокна. К рецептору подходят кровеносные сосуды.
К сухожильному органу часто примыкают свободные нервные окончания и различные инкапсулированные рецепторы: пачини- нодобные тельца, тельца Гольджи—Маццони. В ряде случаев сухожильные органы соединены последовательно с мышечными веретенами (Sherrington, 1894; Bridgman et al., 1969). При таком расположении сухожильные органы Гольджи должны включаться в работу при активации интрафузальных мышечных волокон.
Мышечные веретена. Они были описаны в середине прошлого столетия (см. обзор: Granit, 1955) и с тех пор привлекают к себе постоянное внимание. Под световым микроскопом мышечное веретено представляет собой вытянутую структуру, расширенную в середине за счет капсулы и напоминающую по форме веретено, что и обусловило ее название (Kiihne, 1863). Ввиду большой значимости в жизнедеятельности организмов, а также относительной доступности, мышечные веретена оказались к настоящему времени наиболее изученными механорецепторами. Библиография работ в этой области дана в специальной сводке (Eldred et al., 1967), а также в последних обзорах (Granit, 1970; Matthews, 1972).III
Морфология мышечных веретен и их иннервация наиболее подробно исследовались у амфибий (Е. G. Gray, 1957, 1958; Barker, Соре, 1962а; Robertson, 1960; Katz, 1961; Karlsson et al., 1966; Karlsson, 1972) и млекопитающих (Barker, 1948, 1962, 1967; Merrillees, 1960; Boyd, 1962; Cooper, Daniel, 1963; Adal, Barker, 1965, 1967; Jones, 1966; Corvaja et al., 1969; Barker et al., 1970; Ovalle, 1972; Swash, Fox, 1972, и др.). Менее детально изучались мышечные веретена рептилий и птиц (Saglam, 1968; Crowe, Ra- gab, 1970a, 1970b; Fukami, Hunt, 1970; Maier, Eldred, 1971; Ballot, Ridge, 1972, 1973). У рыб мышечные веретена отсутствуют, однако большое количество проприоцепторов располагается в плавниках. От этих плавниковых сухожильных рецепторов отходят толстые нервные волокна, аналогичные афферентным аксонам первичных окончаний мышечных веретен.
Хотя мышечные веретена впервые появляются у амфибий, наиболее просто они организованы у рептилий. Самое же сложное строение мышечных веретен наблюдается у млекопитающих. Недаром много работавший в этой области Руффини справедливо считал их после специализированных органов чувств самыми сложными рецепторными образованиями организма.
Мышечные веретена неравномерно распределены в теле животного. В отдельных мышцах, таких как экстраокулярные мышцы у кошки и кролика, они вообще отсутствуют (но встречаются в аналогичных мышцах человека). Всего 1—2 веретена обнаружено в мелких мышцах хвоста крысы. Напротив, в крупных мышцах человека их может быть до нескольких сот. Наибольшая плотность (оцениваемая по индексу Восса — числу рецепторов на единицу веса мышцы) обнаружена в небольших мышцах рук, совершающих тонкие, высококоординированные движения (Voss, 1937, 1956а, 1956Ь, 1959; Schulze, 1955; Cooper, 1966а, 1966b).
Как и в других рецепторах, в мышечном веретене имеются вспомогательные структуры, представленные специализированными поперечнополосатыми мышечными волокнами, получившими наименование интрафу зальных (в отличие от обычных, экстра- фу зальных волокон мышечной ткани), а также капсулой рецептора, заполненной жидкостью.
Расположение мышечных веретен таково, что они включены как бы параллельно основным экстрафузальным волокнам скелетной мышцы. При сокращениях экстрафузальных мышц интра- фузальные волокна с окружающими их нервными волокнами испытывают меньшую нагрузку, т. е. разгружаются. Сухожильные органы, напротив, расположены последовательно с экстрафу- зальными мышечными волокнами и при их сокращении испытывают возрастающую нагрузку. Указанное различие между этими типами мышечных рецепторных приборов (Fulton, Pi-Suner, 1928) широко использовалось физиологами при исследовании мышечных веретен и сухожильных рецепторов.
Интрафузальные волокна получают моторную иннервацию, благодаря которой они активно участвуют в деятельности мышечных рецепторов. Собственно рецептирующими элементами веретена являются окончания тонких нервных волоконец, оплетаю щих структуры интрафузальных мышц в их центральной зоне. За такой на первый взгляд обычной для механорецепторов картиной строения мышечных рецепторов стоит крайне сложная структура веретена, которая весьма разнится не только у животных разных классов, но и у представителей одного класса. Хотя изучению строения мышечных веретен посвящено огромное число работ, полной ясности в этом вопросе нет до настоящего времени. Основные сведения по структуре веретен были получены в экспериментах на лягушке, кролике и кошке.
Основу веретена составляют пучки параллельно расположенных интрафузальных волокон. Число волокон, так же как их размер, могут сильно варьировать. Так, у рептилий имеется всего одно волокно, у земноводных (лягушки) — 3—12, у млекопитающих (кошки) — 2—13 (в среднем 6). Диаметр] волокон может колебаться от 6 до 28 мкм, при этом в составе даже одного пучка могут оказаться как толстые, так и тонкие волокна. Длина волокон может достигать нескольких миллиметров. Волокна, входящие в состав веретена, могут начинаться от разных экстра- фузальных волокон, но сходятся они у дистального конца.
У лягушки каждое интрафузальное волокно состоит из двух плотных поперечноисчерченных сократимых и полярно расположенных частей (около 150 мкм каждая), а также центральной, специализированной области (рис. 8). Исчерченность волокон отсутствует в центральной зоне на протяжении 50—100 мкм. В этой зоне число миофиламентов резко уменьшается (на 85%), вместо них появляется сложная сеть тонких соединительнотканных волокон. Поэтому данная область является обычно несократимой. Центральная область получила наименование ретикулярной зоны, а соседние с ней участки — компактных зон (Katz, 1961). С функциональной точки зрения, существование различных областей означает наличие в волокне более упругих (ретикулярных) и более вязких (компактных) участков.
Ретикулярная область может рассматриваться как область с «малой надежностью» в отношении распространения мышечного спайка вдоль волокна. В тонких же интрафузальных волокнах амфибий, где нет ретикулярной области, мышечный потенциал при его распространении из одного конца волокна в другой не блокируется (R. S. Smith, 1964а, 1964Ъ).
В отношении строения интрафузальных волокон млекопитающих вплоть до 1956 г. сохранялась классическая схема. Согласно этому представлению (см. обзорную работу: Barker, 1948), в центре интрафузального волокна находится несократимый участок, получивший наименование области ядерной сумки. С обеих сторон к этой зоне примыкают участки, получившие наименование областей миотрубки, которые отделяют область ядерной сумки от обычных мышечных структур интрафузального волокна. В областях миотрубки под световым микроскопом было видно постепенное исчезновение (в направлении к центру) поперечной исчер- ченности волокна. Именно в областях ядерной сумки и миотрубки и локализуются сенсорные нервные окончания веретена.
В 50-х годах на человеческом материале (Cooper, Daniel, 1956) и при исследовании кошек (Boyd, 1956) было впервые обнаружено существование двух типов веретен. В дальнейшем эти данные были подтверждены другими работами (Walker, 1958; Swett, Eldred, 1960b; Barker, Gidumal, 1961; Boyd, 1962; Cooper, Daniel, 1963). Обнаруженные различия между веретенами относились к строению экваториальной области интрафузальных волокон.
В полярных областях йнтрафузальных волокон хорошо видны ядра, рассеянные в основном по периферии волокна, как это имеет место в обычных экстрафузальных мышечных волокнах. Ближе к центральной области, где волокно покрывается капсулой, в нем появляются дополнительные ядра, располагающиеся преимущественно в центре, аналогично тому, как это бывает в сердечной или в развивающейся скелетной мышце. Концентрация ядер достигает максимума в центре волокна. Это и послужило в свое время основанием называть центральную зону волокна областью ядерной сумки. (Сразу же следует заметить, что никакой специальной сумки для ядер, иной, чем сарколемма самого волокна, нет). Волокна такого типа получили наименование волокон с ядерной сумкой — ЯС-волокон.
У волокон другого типа, которые в большинстве случаев являются более тонкими и короткими, концентрация ядер в экваториальной области меньше, и они располагаются в ряд, образуя цепочки. Такие волокна получили наименование волокон с ядерной цепочкой — ЯЦ-волокон. У некоторых животных, например кроликов, ЯЦ-во- локна отсутствуют.
МОб МОБ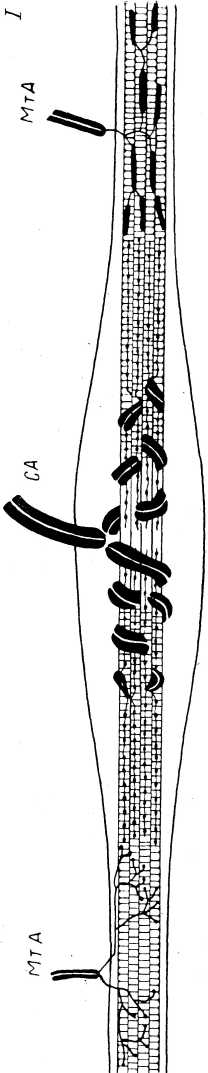

МОб*~~^~ СОб ~
100 мкм
й
о СП ел
-й
ей
05
(Л
я
о (Я
00
«о
о
О§
а! ой* • о 2 я я 5 я <n s а
И
1
S я£и
к
I
Iss
я
й а о &о
Q
S Я М
§ о §11 33£ « ё?4
f>»
О.» Я ей Д Я
й
05
Я
ей к ей £=§£ „Й S Й.Я '-'б'И и8§
g S к £ I« и2«
и**! S* «°8 :§! ^Ii 1|§ О Я о 1
§i-
•-UH Ен Н о Q • -я й 8&ё
£я5
Ф
й о «II ® S 8 о я ° Й1
8§§
в“° <h | gi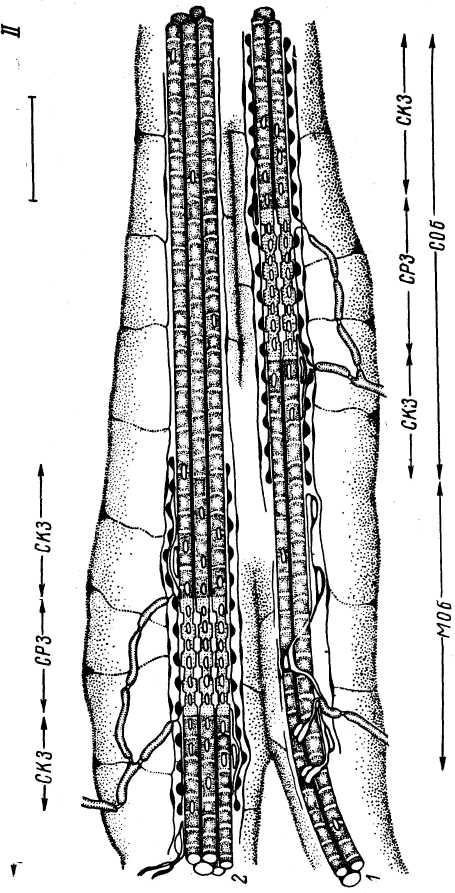
Hunt, 1964). Какой-либо резкой границы между миотрубкой и обычными частями интрафу зального волокна нет.
У волокон обоих типов центральная ядерная область невелика, достигая всего лишь 300 мкм (ядерные сумки у человека имеют длину 100—250 мкм), а при растяжении — 500 мкм. В обоих случаях миофибриллы в центральной области истончаются и образуют очень тонкий слой. В ЯЦ-волокнах миофибриллы содержатся в большем количестве, чем в ЯС-волокнах (Landon, 1966; Adal, 1969; Corvaja et al., 1969). Вблизи экваториальной области отмечается наличие эластической ткани. Описано распределение в ЯС- и ЯЦ-волокнах эластической ткани, играющей важную роль в формировании механических свойств веретен (Cooper, Daniel, 1963, 1967).
К настоящему времени накопилось много данных, говорящих о существовании значительных различий между ЯЦ- и ЯС-волок- нами. Как показали электронномикроскопические наблюдения, ЯЦ-волокна содержат большее число митохондрий, чем ЯС-ВО’ локна. Это позволяет, применяя гистохимические методы, основанные на реакции с митохондриальной АТФазой и сукцинатдегидрогеназой. довольно избирательно окрашивать ЯЦ-волокна (Henneman, Olson, 1965; Barker et al., 1970). ЯЦ-волокна содержат также больше миофибриллярной АТФазы (Nystrom, 1967; Spiro, Beilin, 1969а). ЯС-волокна, в свою очередь, имеют большее количество миогемоглобина (James, 1968).
Число ЯЦ- и ЯС-волокон сильно разнится в разных мышцах и у разных животных. Например, в крупных веретенах человека, где общее число волокон может достигать 14 и более, 3—4 волокна относятся к ЯС-волокнам, а остальные к ЯЦ-волокнам (Cooper, Daniel, 1963). ЯЦ-волокна чаще встречаются у животных, обладающих более тонкой координацией движений.
В пределах одного веретена интрафузальные ЯЦ-волокна могут иногда делиться. Длинные и достаточно толстые ЯС-волокна могут несколько раз образовывать ядерные сумки, входить в состав последовательно соединенных веретен, которые получили наименование тандемных (Е. G. Gray, 1957; Barker, Соре, 1962а; Boyd, 1962; Cooper, Daniel, 1963). Волокна такого типа довольно часто встречаются у лягушек.
Центральная область обычных волокон, входящих в состав одного веретена, окружена капсулой. Величина ее может достигать у человека 5 мм и более (Cooper, Daniel, 1963). ЯС-волокна и длинные ЯЦ-волокна могут простираться далеко за пределы капсулы. В этих случаях она может покрывать лишь х/3 общей длины волокон. Более же короткие ЯЦ-волокна могут оканчиваться вблизи концов капсулы, прикрепляясь либо к ее стенкам, либо к ЯС-волокнам (Boyd et al., 1962; Bridgman et al., 1969). При изолированном сокращении ЯЦ-волокон, закрепленных'на капсуле, область соседних ЯС-волокон веретена будет испытывать меньшую нагрузку.
Наибольшего размера (80—200 мкм) капсула достигает в средней части. Упругость капсулы, так же как и интрафузальных волокон, увеличивается за счет эластических волокон (Cooper, Daniel, 1963, 1967). Ультраструктура капсулы как у млекопитающих, так и у амфибий рассматривалась многими авторами (Merrillees, 1960; Katz, 1961; Karlsson et al., 1966; Landon, 1966; Corvaja et al., 1969).
У лягушки капсула веретена состоит из двух слоев: наружного и внутреннего, которые соединены между собой рядом поперечно направленных пучков. В ретикулярной области каждое интрафузальное волокно окружено индивидуальной внутренней капсулой. Временами (обычно несколько от- тупя от экваториальной области) 2—3 интрафузальных волокна объединяются в пучок и оказываются окруженными единой внутренней капсулой. Наружная капсула может заключать в себе несколько таких пучков. Пример частичного объединения наружной капсулой двух пучков волокон у лягушки представлен на рис. 8, II, Внутренняя капсула имеет меньшую протяженность по сравнению с наружной. Она ограничивается преимущественно ретикулярной и компактными зонами и, постепенно уменьшаясь в толщине, исчезает, не соединяясь с наружной капсулой.
Наружная капсула образована несколькими (9—12) слоями уплощенных клеток, плотно перекрывающих друг друга и связанных межклеточными соединениями. Никаких специальных пор в капсуле нет, помимо щелей вдоль входящих в веретено сенсорных и моторных нервных волокон. Ближе к полярным частям структуры наружной капсулы истончаются. Появляются ходы, соединяющие внутреннее и наружное пространство. В клеточных структурах наружной капсулы видны митохондрии и различные включения. Капсула оплетена коллагеновыми волокнами, идущими в разных направлениях. Внутренняя оболочка состоит из одного слоя клеток, получивших наименование эндомизиалъных (Merrillees, 1960). Они похожи на клетки наружной капсулы, но содержат меньше цитоплазмы. Количество везикул в них ограничено (Landon, 1966; Corvaja et al., 1969).
Капилляры, обильно снабжающие веретена, как правило, обнаруживаются снаружи капсулы или между слоями наружной капсулы. В отдельных случаях ближе к полярным частям капсулы они могут встречаться и на внутренней поверхности наружной капсулы (Barker, 1948; Cooper, Daniel, 1963; Corvaja et al., 1969).
В центральной области веретена, где капсула отходит от интрафузальных волокон, имеется периаксиалъное пространство, заполненное жидкостью. По мнению части исследователей (Sherrington, 1894; Jahn, 1959), это пространство связано с лимфатической системой, и его поэтому можно назвать лимфатическим. Однако во внутрикапсулярной жидкости обнаружено большое количество кислых мукополисахаридов (Brzezinski, 1961а). Такое отличие в составе лимфы и жидкости капсулы заставило автора предположить, что гиалуроновые кислоты образуются оболочкой веретена. Таким образом, можно полагать, что между лимфатическим пространством и внутрикапсулярным пространством имеется диффузионный барьер. В пользу такого представления свидетельствуют и данные авторов, отмечавших затруднения в проникновении веществ через капсулу веретен (Calma, Kidd, 1962; Kidd, 1964). Об участии клеток капсулы в формировании состава внутрикап- сулярной жидкости говорят и наблюдения морфологов, обнаруживших обилие везикул различных размеров в элементах наружной капсулы (Corvaja et al., 1969). Существование такого рода структур, по-видимому, является результатом интенсивного транспорта веществ через мембраны клеток (Bennett, 1956).
Как далеко простирается существование известных различий в составе между внутрикапсулярной жидкостью веретена и лимфой, сегодня еще неясно. Можно пока только сослаться на результаты экспериментов, проведенных на других тканевых высокоспециализированных механорецепторах — тельцах Пачини (Ильинский, Красникова, 1972). Как показали прямые измерения, в жидкости телец Пачини имеется повышенная концентрация ионов калия и пониженное содержание белка по сравнению с плазмой крови. Необычный состав среды, окружающей различные механорецепторы, может играть важную роль в механизмах работы этих сенсорных структур (глава 7).
Гиалуроновые кислоты благодаря своей вязкости, по-видимому, могут существенно влиять на механические свойства капсулы, заполненной жидкостью. Предполагается, что внутрикап- сулярная жидкость играет роль в распределении давления внутри веретена (Bridgman, Eldred, 1964).
К мышечному веретену подходит большое количество нервных волокон, разветвляющихся в его разных частях. Основная масса разветвлений находится в области, покрытой капсулой рецептора.
Исследованиями многочисленных авторов (Sherrington, 1894; Weiss, Dutil, 1896; Ruffini, 1897; Hinsey, 1927-1928; Hines, Tower, 1928; Barker, 1948, и мн. др.) было установлено существование в интрафузальных мышцах млекопитающих трех основных типов нервных окончаний. Из них два типа следует отнести к сенсорным окончаниям, а один — к моторным. Сенсорные окончания одного типа получили наименование первичных, или аннулоспиральных, а другого типа — вторичных, или гроздевидных. Пластинчатые же окончания в мышцах оказались двигательной природы. Термины аннулоспиральные и гроздевидные окончания, хотя и по сей день встречаются в литературе, по-видимому, менее удачны, чем первичные и вторичные, так как в ряде случаев, например у кролика и человека, на основании морфологических критериев разделить сенсорные окончания на типы не удается. Однако они разнятся по месту своего расположения в веретене (Barker, 1948). Первичные окончания локализуются в экваториальной части волокон, т. е. в середине центрального участка, в области ядерной сумки; вторичные же окончания располагаются в области миотрубки, в зоне, примыкающей к этому экваториальному участку. Двигательные окончания рассеяны по остальным частям интрафузальных волокон. У лягушки веретена обладают всего лишь одним типом сенсорных нервных окончаний. Они локализуются в центральной сенсорной области.
Йе все веретена у млекопитающих содержат сенсорные окончания обоих типов. Часть из них обладает лишь первичными окончаниями (так называемые простые веретена); другие имеют как первичные, 1ак и вторичные окончания (промежуточные); третьи наряду с первичными содержат большое количество разнообразных вторичных окончаний (сложные веретена). Простые веретена часто имеют короткие интрафу зальные волокна. Их можно видеть в шейных мышцах. Напротив, сложные веретена имеют более длинные мышечные волокна и встречаются в мышцах конечностей (Ruffini, 1897; Barker, 1959, 1962; Eldred et al., 1962; Cooper, Daniel, 1963).
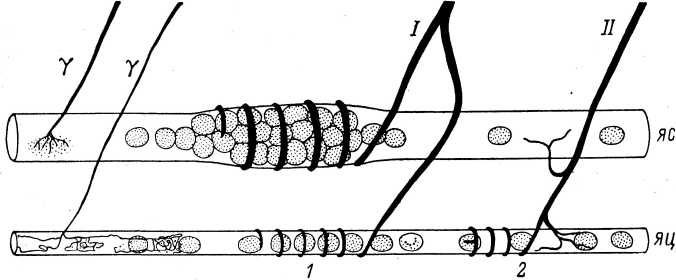
Рис. 9. Иннервация двух типов (ЯС и ЯЦ) интрафузальных мышечных волокон веретена млекопитающих (по: Matthews, 1964).
I, II — афферентные волокна (группы I и II); 1, 2 — первичные и вторичные окончания; Т — гамма-моторные волокна.
Первичные окончания у млекопитающих встречаются как в ЯС-, так и в ЯЦ-волокнах, и имеют вид спиралей и колец (более мелких у ЯЦ-волокон), обвивающих мышечные волокна в их экваториальной области. В центре ядерной зоны область разветвлений первичных окончаний составляет в длину, примерно, 300 мкм (Boyd, 1962; Cooper, Daniel, 1963; Barker, 1967). Нервные окончания образуются разветвлениями миелинизированных волокон, которые являются наиболее толстыми нервными проводниками млекопитающих. В нервном стволе их диаметр достигает 20 мкм. По современной классификации их относят к волокнам группы la (Lloyd, 1943; Bradley, Eccles, 1953; см. также о нервных волокнах мышц в обзоре: Matthews, 1972).
Вторичные окончания разветвляются в основном в структурах ЯЦ-волокон и значительно реже — в ЯС-волокнах (рис. 9). Образуются они разветвлениями волокон группы II. Веретено может иметь до 5 вторичных окончаний (Boyd, 1962). Если волокна
группы la не дают разветвлений и иннервируют лишь одно веретено, то волокна группы II делятся, и их веточки могут подходить к вторичным окончаниям в других веретенах. В мышцах кошки на два первичных окончания приходится в среднем три вторичных (Boyd, 1962).
В сложных веретенах, имеющих вторичные окончания, волокна первичных окончаний имеют больший диаметр, чем в простых веретенах. Вторичные окончания, лежащие ближе к экваториальной области, связаны с более толстыми волокнами, чем те окончания, которые локализуются дальше от центральной зоны волокна. Более центрально расположенные вторичные окончания кошек имеют чаще вид колец и спиралей (аннулоспиральный вид), а локализованные более периферически — гроздевидную форму. Между областями разветвлений первичных и вторичных окончаний на ЯЦ-волокнах кошки имеется участок, свободный от сенсорных окончаний (Corvaja et al., 1969).
Нервные волокна, образующие чувствительные окончания, подходят к веретену в его центральной части, проникают наискось через наружную капсулу и затем уже в самом внутрикапсу- лярном пространстве теряют миелин, но остаются окруженными шванновскими клетками, которые затем исчезают в зоне контакта терминалей с мышечными волокнами. На рис. 8, II видно, что миелин теряется в переходном участке между сенсорной ретикулярной и сенсорной компактной зонами. Как видно на этом рисунке, внутри капсулы нервное волокно имеет один или несколько перехватов Ранвье.
Нервное волокно сильно ветвится. У лягушки, например, обнаружено 6 ступеней дихотомических разветвлений одиночного сенсорного волокна, так что общее число концевых колб может достигать 10 000. При этом сенсорные терминали образуются тонкими цилиндрическими трубочками (около 0.15 мкм в диаметре), соединяющими между собой варикозные расширения нервных волокон, которые тесно связаны с мышечными элементами с помощью соединительнотканных нитей (Katz, 1961). У кошки в экваториальной области ЯС-волокон сенсорные окончания имеют на поперечном сечении диаметр 0.5—3 мкм. В области миотрубки размеры терминалей нервных волокон меньше — не более 1 мкм (Merrillees, 1960; Corvaja et al., 1969).
Сенсорные окончания тесно примыкают к поверхности мышечных волокон. У лягушки в ретикулярной зоне сенсорные колбы находятся в тесном контакте с регулярно расположенными мышечными отростками, заключающими в себе один саркомер контрактильного материала (Katz, 1961; Karlsson et al., 1966). У кошки нервные окончания лежат в неглубоких канавках, расположенных на поверхности интрафузальных мышечных волокон со всех их сторон. Иногда они посылают пальцеобразные отростки, которые могут достигать области ядерной мембраны (Corvaja et al., 1969). Мембраны нервных окончаний и мышечных волокон разделены щелью в 150—200 А. Базальная мембрана покрывает окончания, однако, между нервными элементами и мышечной тканью она отсутствует. Нервные окончания содержат типичные нейрофиламенты, большое количество митохондрий, везикул, микротрубочек. Число митохондрий, которые обычно имеют небольшие размеры, иногда столь велико, что они как бы полностью забивают окончание. Размер везикул составляет 200— 1000 А, микротрубочек — 200—270 А. Тонкая структура первичных и вторичных окончаний одинакова.
В мышечных веретенах как амфибий, так и млекопитающих было обнаружено большое количество особых клеток, которые получили наименование клеток-сателлитпов (Katz, 1961; Karlsson et al., 1966; Landon, 1966; Corvaja et al., 1969). У лягушек основная масса клеток-сателлитов расположена в сенсорной компактной зоне и отчасти в ретикулярной области. Они тесно связаны как с интрафузальными мышечными волокнами, так и с сенсорными окончаниями. У кошки эти клетки, имеющие небольшие размеры, локализуются как в ЯС-, так и ЯЦ-волокнах в их полярных частях, будучи вытянутыми вдоль длинной оси волокон. Тонкий слой цитоплазмы клеток содержит рибосомы, митохондрии, разнообразные пиноцитозные везикулы. В отличие от нервных окончаний число митохондрий в клетках незначительно. Функция клеток-сателлитов пока не ясна.
Существование двигательной иннервации веретён было отмечено давно (Kerschner, 1883, 1893; Weiss, Dutil, 1896; Cipollone, 1897-1898; Huber, DeWitt, 1897; Hinsey, 1927-1928; Hines, Tower, 1928, и др.). Известно, что эфферентные волокна, направляющиеся в мышцу, состоят из двух основных групп: толстых мякотных волокон {алъфа-группа) и тонких волокон {гамма- группа) (Eccles, Sherrington, 1930). Первоначально считалось, что двигательные волокна интрафузальных мышц представлены почти исключительно гамма-волокнами, которые заканчиваются пластинчатыми окончаниями, аналогичными тем, которые существуют в экстрафузальной мускулатуре и которые являются концевыми органами толстых альфа-волокон (Barker, 1948). Последующее усовершенствование импрегнационных методик, применение новых гистохимических методов равно как и использование электронномикроскопической техники позволило существенно дополнить и углубить эту первоначальную схему двигательной иннервации.
Согласно существующим в настоящее время представлениям, в интрафузальной мускулатуре млекопитающих могут находиться три вида окончаний двигательных {фузимоторных) (Hunt, Pain- tal, 1958) нервных волокон (рис. 10), а именно: 1) гамма-кустиковидные диффузные окончания гроздевидной извитой формы (gamma trails); 2) гамма-пластинки (gamma plates), именуемые еще окончаниями р2, или окончаниями II типа; 3) пластинчатые окончания р1? или окончания I типа (Coers, Durand, 1956; Cooper,
Daniel, 1956; Hess, 1961; Boyd, 1962; Barker, Cope, 1962b; Coers, 1962;. Barker, 1967; Corvaja et al., 1969; Hennig, 1969; Barker et al., 1970; см. также труды симпозиума: Granit (ed.), 1966; и обзоры: Granit, 1970; Matthews, 1972).
Гамма-кустиковидные окончания являются концевыми органами волокон гамма-группы и локализуются наиболее центрально из всех двигательных окончаний. Область их распространения может достигать 1 мм в каждую сторону от экваториальной зоны. Гамма-пластинки (р2, окончания II типа) также являются концевыми структурами гамма-фузимоторных волокон. Помимо своего строения, они отличаются от окончаний первой группы и более периферической локализацией на интрафузальном волокне. Пластинчатые окончания (I типа) практически ничем не отличаются от аналогичных структур обычной мускулатуры. Они связаны с бета-волокнами (или медленными альфа-волокнами), которые могут быть веточками обычных экстрафузальных, двигательных нервов. Локализуются они, как правило, у концов интрафузаль- ных волокон в областях их полюсов.
Некоторые авторы считают, что ЯС-волокна содержат преимущественно гамма-пластинки, а ЯЦ-волокна — гамма-кустиковидные окончания (Boyd, Davey, 1966). Последнее утверждение не носит, по-видимому, всеобщего характера, так как известны веретена, содержащие оба вида нервных окончаний (например, у кролика, который имеет лишь волокна ЯС-типа). Можно еще отметить, что удалось наблюдать веретено у крысы, к которому подходило два моторных нервных волокна. Одно из них иннервировало исключительно ЯС-, а другое — ЯЦ-волокна. При этом на обоих типах интрафузальных волокон удалось обнаружить разного вида моторные окончания (Poryako, Smith, 1968). Таким образом, оказалось, что одно и то же нервное волокно может образовывать различные терминали на мышечных структурах, и вид окончаний может не зависеть от типа интрафузального волокна.
Количественная оценка встречаемости различных видов окончаний показала, что 91 % веретен имеет гамма-кустиковидные окончания, 78% — гамма-пластинки (р2) и 58% — пластинчатые окончания (Pi) (Barker, Ip, 1966).
Рис. 10. Моторная иннервация ЯС- и ЯЦ-волокон мышечных веретен кошки (по: Barker et al., 1970).
ЭМВ — экстрафузальные мышечные волокна; pt — пластинчатые окончания (ЯС=75%, ЯЦ=25%); р2 — гамма-пластинки (ЯС = 90%, ЯЦ=10%); рхВ и р2В — волокна, заканчивающиеся р, и р2; Мтпа и Мшр — моторные альфа- и бета-волокна; РОб — область разветвления кустиковидных окончаний; СВ — сосудистое вздутие аксона; МВ и НмВ — миелинизированные и немиелинизированные волокна кустиковидных окончаний; ВПА — возвратные претерминальные аксоны (стрелка) кустиковидных окончаний; Фа — быстрые фу зимоторные альфа-волокна. Справа — схемы электр оннрмикроскопическЪй картины трех типов фузимоторных мионевральных соединений,

У лягушки двигательная иннервация веретена осуществляется большим (5—12 мкм) и малым (2—3.5 мкм) нервными волокнами. Более толстое волокно образует пластинчатый (en plaques) тип нервных окончаний, а более тонкое — кустиковидный (en grappes) (Е. G. Gray, 1957). Моторные волокна веретена берут начало от обычных интрафузальных двигательных нервов. Волокна большого диаметра оканчиваются на фазных, быстрых мышцах, а волокна малого диаметра на тонических, медленных мышечных структурах.
К веретену обычно подходит несколько двигательных волокон (7—23), зоны иннервации которых перекрывают друг друга (Hagbarth, Wohlfart, 1952; Boyd, 1962). Волокна, подходящие к одному веретену, могут быть связаны даже с 2—3 разными (смежными) сегментами спинного мозга (Guajunco, 1932). В свою очередь одно и то же гамма-волокно может давать веточки к различным веретенам (Hunt, Kuffler, 1951а; Steg, 1964; Boyd, Davey, 1966, 1968). Однако могут встречаться и веретена, полностью лишенные двигательной иннервации (Jones, 1966; Barker, 1967).
На рис. 10 представлена схема моторной иннервации ЯС- и ЯЦ- волокон мышечных веретен mm. peroneus et soleus кошки. Видно, что немиелинизированные волокна кустиковидных окончаний не всегда участвуют в иннервации: в m. peroneus они наблюдаются в дополнение к миелиновым волокнам кустиковидных окончаний несколько более, чем в 1/3 веретен. Менее 5% веретен обладают возвратными претерминальными аксонами кустиковидных окончаний, покидающими их. Некоторые р± волокна могут быть идентифицированы как коллатерали бета-волокон, которые также иннервируют экстрафузальные мышечные волокна.
Таким образом, картина двигательной иннервации веретен может быть чрезвычайно разнообразной, что говорит о широких функциональных возможностях фузимоторной системы.
У млекопитающих моторные нервные волокна имеют различный диаметр. Он варьирует у разных животных в широких пределах: например, диаметр волокон нерва m. soleus кошки равен 1-—18 мкм, а т. soleus кролика — 2—14 мкм (Barker, 1966). Волокна теряют миелин вскоре после, а иногда даже до проникновения в пространство капсулы веретена. Они многократно делятся и, постепенно истончаясь, образуют окончания на различных частях интрафузальных мышечных волокон. Претерминальные аксоны окружены тонкими цитоплазматическими структурами шванновских клеток, которые отделены от аксона щелью порядка 150 А. Терминальные волокна лежат на поверхности интрафузальных мышечных волокон. Они содержат различные число митохондрий и большое количество маленьких везикул (у кошки 350—450А в диаметре — Gorvaja et al,, 1969).
Тонкое строение моторных окончаний напоминает структуру аналогичных окончаний в экстрафузальной мускулатуре (Merril- 44
loos, 1960; Gruner, 1961; Katz, 1961; Karlsson et al., 1966; Landon, 1966; Adal, Barker, 1967; Gorvaja et al., 1969; сравн.: Palade, 1954; Kobertson, 1956; Reger, 1958; Hess, 1961, и др.).
Кустиковидные окончания представляют собой луковичные структуры, покрытые шванновскими клетками, которые изолируют их от окружающих тканей. Расстояние между мембраной нервного окончания и поверхностью мышечного волокна составляет примерно 500 А. Складчатость постсинаптической мембраны отсутствует. Нервные окончания содержат большое количество везикул (200—1000 А); встречаются и более крупные (свыше 1000 А) везикулы. В, ряде случаев можно видеть, как содержимое везикул изливается в синаптическую щель.
Пластинчатые окончания в отличие от кустиковидных глубоко проникают в интрафузальное волокно. Однако во всех случаях между нервной и мышечной тканью располагается базальная мембрана. Наблюдается складчатость плазматической и базальной мембраны интрафузального волокна в местах контакта с нервным окончанием, хотя и не в такой степени выраженная, как в случае экстрафузальной мускулатуры. Как и в случае кустиковидных окончаний, нервное волокно в местах контактов содержит скопление митохондрий. Нервные окончания заполнены большим количеством везикул.
Собственно веретено, по-видимому, не получает симпатической иннервации (Boyd, 1962). Все тонкие немиелинизированные волокна, снабжающие кустиковидные окончания, несомненно, относятся к соматическим нервам. Однако сосуды веретена получают иннервацию симпатического происхождения (Hinsey, 1927— 1928). В последнее время, правда, появились сведения, говорящие о возможности проникновения в веретена симпатических волокон, не связанных с сосудами (Banker, Girvin, 1971; Santini, Ibata, 1971).
« * *
Как видно из изложенного, в опорно-двигательном аппарате находятся механорецешоры самого различного строения. Это различие заключается прежде всего в особенностях структуры вспомогательного аппарата, достигающего чрезвычайной сложности в мышечных веретенах. Это выражается и в особенностях самих окончаний афферентных волокон в тканях, и в диаметре нервных волокон. Однако несмотря на эту разницу в строении, между всеми рассмотренными механорецепторами имеется и сходство. Ни у одного из них в структуре не удается обнаружить элементов иных, чем сами окончания афферентных нервных волокон, которые могли бы выполнять специфическую функцию рецепции механического раздражения. Это означает, что все рассмотренные рецепторы на основании их строения могут быть отнесены к группе первичных сенсорных приборов.
Механорецепторы органов чувстй
Значительная часть органов чувств животных специально предназначена для восприятия различного вида механических воздействий среды. У позвоночных — это органы боковой линии, вестибулярный аппарат, орган слуха. Все эти органы чувств могут рассматриваться как части единой акустико-латеральной системы (Mayser, 1882; Herrick, 1897; Шмальгаузен, 1938; Lowenstein, 1950, 1967b; Bergeijk, 1967). В силу генетического родства они имеют много сходных черт как в морфологическом строении, так и в функциональной организации своих периферических и центральных частей. Ниже мы остановимся именно на том, что явля- етсясходным в воспринимающих частях данных сенсорных приборов. Вопросы же более специальные, касающиеся специфических особенностей строения, равно как и функционирования каждого из перечисленных органов чувств, мы рассматривать здесь не можем и отсылаем читателя к соответствующим обобщающим работам (по боковой линии: Дислер, 1960; Dijkgraaf, 1963; Винников, 1966; Cahn, 1967; Flock, 1965а, 1971b; по вестибулярному аппарату: Ger- nandt, 1959, Rasmussen, Windle, 1960; Wersall et al., 1965; Винников, 1966; Титова, 1968; Винников и др., 1971; Lowenstein, 1971, 1972; Wersall, 1972; по органу слуха: Davis, 1959, 1961; Bekesy, 1960; Schwartzkopf, 1960, 1962; Винников, Тиюва, 1961; Wersall et al., 1965; Винников, 1966; Spoendlin, 1966, 1968, 1973; Титова, 1968; Angelborg, Engstrom, 1973).
Все органы чувств, предназначенные для восприятия механических стимулов, устроены по одинаковому принципу. Они состоят из вспомогательного аппарата (наиболее простого в органах боковой линии и наиболее сложного в органе слуха), рецепторов (представленных волосковыми клетками) и, наконец, афферентных и эфферентных нервных волокон, контактирующих с волосковыми клетками.
Собственно рецепторные структуры
Собственно рецепторной структурой во всех органах чувств акустико-латеральной системы является волосковая клетка (рис. 11). Как показали многочисленные морфологические исследования, волосковые сенсорные клетки органов боковой линии (см., например: Flock, 1965а; Hama, 1965; Аронова, 1967; Yamada, Hama, 1972; Yamada, 1973) и лабиринта различных животных (например: С. A. Smith, 1956, 1957; lurato, 1961,1967; Roberts, Ryan, 1971; Miller, 1973; см. также обзоры: Wersall, 1956, 1960,1972; Engstrom, Wersall, 1958a, 1959b; Винников и др., 1971; Flock, 1971a; Angelborg, Engstrom, 1973), несмотря на существование некоторых особенностей в их ультраструктуре, имеют в целом сходное строение. Они представляют собой обычно небольшие (20—40 мкм в длину и 5— 10 мкм в ширину) клетки цилиндрической формы. В вестибулярных органах птиц и млекопитающих наряду с этими клетками (тип II) имеются еще колбообразные клетки (тип I) (рис. 12).
В цитоплазме клеток содержится большое количество митохондрий. На своей вершине клетки несут пучок волокон, из-за наличия которых и получили свое наименование.
Волоски бывают двух типов. К одному типу (стереоцилии) относятся неподвижные регулярно расположенные и весьма многочисленные (порядка 30—100 на одной клетке) волоски. Второй тип (киноцилия) представлен, как правило, всего лишь одним более
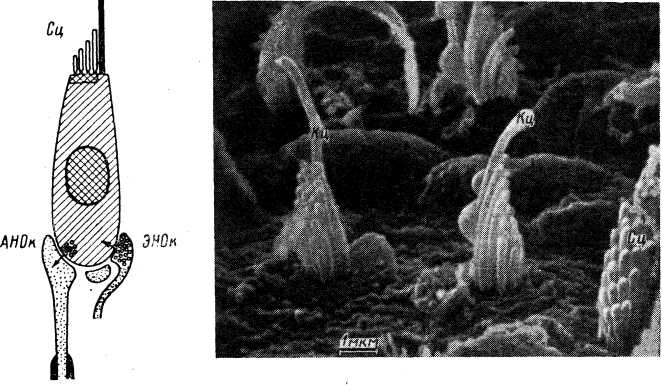
Рис. И. Волосковая клетка (I — по: Flock, 1971а; II — по: Flock, 1971b).
I схема строения волосковой клетки; II — общий вид волосковых механорецепторов
боковой линии рыбы (Lota lota) в электронном сканирующем микроскопе. Кц — киноцилия; Сц — стереоцилия; АНОк — афферентное нервное окончание; ЭНОк — эфферентное нервное окончание.
длинным и подвижным волоском, расположенным на поверхности клетки ассиметрично по отношению к пучку стереоцилий. У млекопитающих волосковые клетки органа слуха утратили киноци- лию. В некоторых случаях (macula utriculi) в одной и той же клетке могут встречаться две киноцилии, находящиеся с противоположных сторон от группы стереоцилий.
Каждая стереоцилия состоит из цитоплазматической сердцевины, окруженной мембраной, которая у основания волоска переходит в клеточную мембрану. В сердцевине стереоцилии располагается пучок многочисленных и очень тонких фибрилл. У основания стереоцилии они сливаются, углубляясь в клетку, и оканчиваются в гранулярной кутикулярной пластине (рис. 13). Размер
стереоцилий в пучке различен. По мере приближения к киноци лии их длина возрастает (рис. 11).
В отличие от стереоцилий, сердцевина киноцилии является более дифференцированной. Она содержит 9 пар волосков, распо-

Рис. 12. Афферентная и эфферентная регуляция двух (Z и II) типов волосковых клеток млекопитающих (по: Wersall, 1967).
Кц — киноцилия; Сц — стереоцилия; АНОк — афферентное нервное окончание; ЭНОк — эфферентное нервное окончание; Син. — синапс; ОКл — опорная клетка; БМ — базальная мембрана.
ложенных равномерно по окружности вокруг одной центральной пары фибрилл. Центральные фибриллы ориентированы определенным обрдзом по отношению к стереоцилиям: линия, соединяющая
эту пару фибрилл, перпендикулярна рядам стереоцилий. Центральные фибриллы оканчиваются в области поверхности клетки, в то время как периферические волокна проникают в глубину и находятся в связи с так называемым базальным тельцем, расположенным у основания киноцилии (рис. 13). В волосковых клетках органа слуха млекопитающих, которые лишены киноцилии, имеется эквивалент базального тельца — центриоля, расположенная также на периферии клетки (см. обзоры: Flock, 1965b, 1971а).
Рис.
13. Схема ультраструктуры стереоцилии
(Сц)
и киноцилии (Ки)
(по: block, 1965а).
Кор.
Сц —
корешок стереоцилии; БТ
—
базальное тельце. Стрелка
—
направление возбуждающего воздействия.
Справа
и слева
—
изображения поперечных сечений на
соответствующих уровнях стерео- и
киноцилии.

По своему строению киноцилия сходна с различными жгутиками и ресничками, встречающимися у различных представителей животного царства (см. обзоры: Винников, 1966, 1971).
Органы боковой линии. Как выяснилось в ходе исследований, проведенных в последние 10—15 лет, сенсорные образования органов боковой линии следует подразделять на два основных типа: так называемые «обычные» (невромасты) и «особые» {«специализированные»') (Dijkgraaf, 1963; Szabo, 1965). Последние еще иногда называют ампулированными, что, по-видимому, не очень точно. Рецепторной структурой «обычных» органов боковой линии является волосковая клетка, аналогичная клеткам органа слуха и вестибулярного аппарата. «Обычные» органы боковой линии выполняют
49
Невромасты встречаются у круглоротых, рыб, амфибий, а «особые» структуры боковой линии обнаружены до настоящего времени только у некоторых рыб (см. обзоры: Акоев и др., 1971; Протасов, 1972). Невромасты расположены в различных местах на коже головы и тела животных. У круглоротых * многих костистых рыб ц у всех водных амфибий имеются свободные невромасты. У большинства же рыб невромасты заключены в специальные каналы, сообщающиеся с внешней средой. Каналы заполнены жидкостью, физико-химические свойства и ионный состав которой могут отличаться от окружающей водной среды и жидкостей тела животного (Ильинский, Красникова, 1971). Каждый невромаст содержит группу волосковых клеток, расположенных среди опорных клеток. Волоски рецепторных клеток погружены в желатинообразный материал, образующий купулу, которая покрывает каждый невромаст (рис. 14). Таким образом, у таких невромастов вся вспомогательная структура рецепторного аппарата ограничена купулой. У невромастов, погруженных в каналы, между внешним стимулом и ре- цептирующей волосковой клеткой могут находиться не только вещество купулы, но еще и структуры, образующие стенку канала, а также жидкость, заполняющая этот канал.
«Особые» рецепторные структуры боковой линии были выявлены у рыб, как обладающих электрическими органами, так и у не имеющих их (например, у акулообразных рыб). В первом случае величина разряда, генерируемого электрическими органами, оказалась различной у разных животных, что позволило разделить всех рыб, обладающих электрическими органами, на сильно- и слабоэлектрические. Поэтому в литературе различают рыб сильноэлектрических, слабоэлектрических, неэлектрических, обладающих электрорецепторами, а также не имеющих электрорецепторов. Соответственно выявлена различная чувствительность разных рыб к действию электрического тока. Законченной и общепринятой классификации «особых» сенсорных образований системы боковой линии пока нет, Подробнее других этот вопрос рассмат-
ривал Сабо (Szabo, 1965), классификация которого может быть принята за основу.
Согласно этой классификации, «особые» образования могут быть подразделены прежде всего на ампулированные и бугорковые
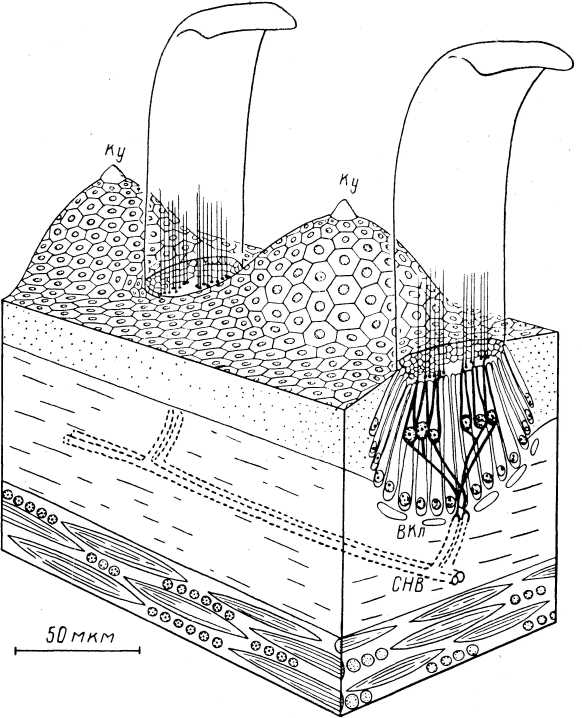
Рис. 14. Примитивные эпидермальные органы боковой линии жабы (Xenopus laevis) (по: Gorner, 1963).
Ку — купулы, обращенные непосредственно в окружающую водную среду; ВКл — волосковые клетки; СНВ — сенсорное волокно.
(тип I и тип II). Среди ампулированных структур можно различить такие, у которых наличие ампул очень хорошо выражено. Здесь прежде всего следует указать на ампулы Лоренцини, которые имеют длинный, расположенный под кожей канал (он может достигать даже десятков сантиметров у крупных скатов). Такой ка
нал открывается с одной стороны в виде поры на поверхность кожи, а с другой стороны оканчивается специальным расширением — ампулой (рис. 15). Весь канал, имеющий большой диаметр (от 200 до 1500 мкм у животных разных видов), заполнен желеобразным веществом особого состава (Murray, Potts, 1961; Ильинский, Красникова, 1971, 1974). Чувствительные элементы локализованы в стенке ампулы (стр. 56, рис. 17).
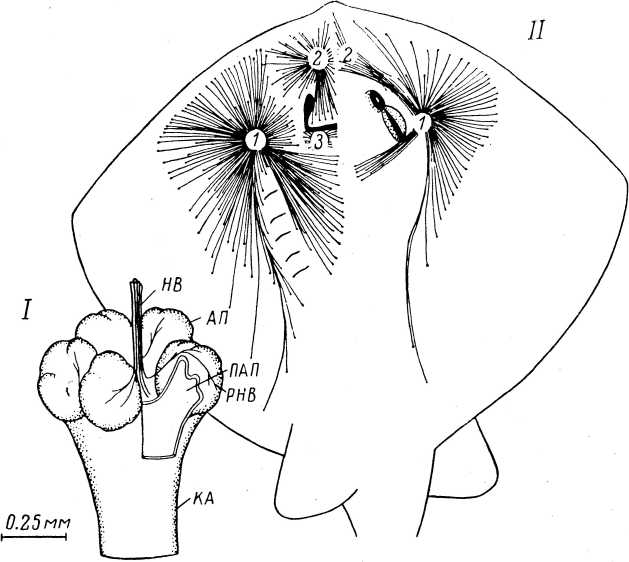
Рис. 15. Ампулы Лоренцини.
— строение ампулы; II — топография ампулярного аппарата Лоренцини на вентральной (слева) и дорзальной (справа) поверхностях ската Dasyatis pastinaca. 1 — гиоидная,
— ростральная из — гио-мандибулярная группы ампул Лоренцини. НВ — нервные волокна; РНВ — разветвления нервных волокон; АП — ампулярные пузырьки; ПАП — полость ампулярного пузырька; КА — канал ампулы.
К ампулированным образованиям относятся также ампулоподобные органы гимнотид и органы типа А мормирид и гимнар- хуса (представители слабоэлектрических рыб). Они характеризуются каналом, проникающим через всю толщу эпидермиса, и ампуловидным расширением на конце канала. Канал и ампула заполнены студенистым веществом.
К ампулярным органам относят также (Mullinger, 1964) и ям- новый орган (small pit organ), описанныйТерриком (Herrick, 1901)
у карликового сомика [Amiurus (letalurus) nebulosus].*Это животное является неэлектрическим. По мнению Сабо (Szabo, 1965), ямковый орган благодаря особенностям своего строения и иннервации может рассматриваться как переходная форма между ампулярными органами слабоэлектрических рыб и ампулами Лорен- цини, которые встречаются у сильноэлектрических, слабоэлектрических и неэлектрических рыб.
Бугорковые органы (tuberous organs) обнаружены у многих слабоэлектрических рыб (гимнотид, мормирид и др.). Они не имеют ни ампулы, ни протоков, заполненных желеобразным веществом. По данным Сабо (Szabo, 1965), у гимнотид бугорковый орган залегает, как и орган I типа, в инвагинации базальной мембраны. Бугорок поддерживающих клеток выступает из самой глубокой части этой инвагинации. Тонкие чувствительные клетки прикрепляются своим основанием к верхушке поддерживающих клеток, а их тело свободно лежит в полости, ограниченной как этими поддерживающими, так и особыми эпителиальными клетками.
В отдельную группу Сабо относит «особые» органы боковой линии, описанные у мормирид (органы типа В, мормиромаст типа II: Cordier, 1938; Szabo, Barets, 1963). У этих структур чувствительные клетки группируются вокруг основания интраэпидермальной полости, заполненной прозрачным содержимым, отличным от содержимого ампулярных органов. Так же как у бугорковых органов, полость не имеет никакого выхода на поверхность тела животного. Чувствительные клетки по своему строению делятся на два вида (Barets, Szabo, 1964). Детали строения, иннервации и расположения на теле животных различных «особых» органов боковой линии можно найти в многочисленных специальных работах (Dijkgraaf, Kalmijn, 1963; Mullinger, 1964; Murray, 1965a; Szabo, 1965; Bennett, 1967; Lissmann, Mullinger, 1968).
Как было выяснено при исследовании функциональных характеристик «особых» образований боковой линии, чувствительностью к механическим действиям обладали некоторые бугорковые и многочисленные ампулированные образования и прежде всего те из них, воспринимающим элементом которых являлась рецепти- рующая клетка, снабженная волоском. Такими структурами являются ампулы Лоренцини. Строение их исследовалось весьма подробно (Barets, Szabo, 1962; Waltman, 1966; Derbin, 1970; Szabo, 1972; Ильинский и др., 1974),
Стенка канала ампулы Лоренцини состоит из нескольких слоев эпителиальных клеток и обладает благодаря этому очень высоким сопротивлением (до 6 мом на 1 см2). Дно ампулы выстлано двумя типами клеток: опорными и рецепторными (стр. 56, рис. 17). Лишь незначительная поверхность соприкасается с содержимым ампулы. Волосок рецептирующих клеток содержит 8 1 или 9. 0 фибрилл.
В отличие от ампул Лоренцини, рецепторы яйковых оргайоЁ не имеют цилиарного аппарата (Mullinger, 1964). Он отсутствует также у рецепторов ампул Gymnotidae (Lissmann, Mullinger, 1968), мормиромастов Gnathonemus (Barets, Szabo, 1964), бугорковых органов разных рыб (Szabo, 1965) и в ряде других случаев.
Вестибулярный аппарат. В вестибулярном аппарате- рецепторные структуры сгруппированы всего лишь в некоторых местах:
Кость
Вестибулярная
лестница (перилимфа)
полоска
Улитковый
канал
(эндолимфа)
Тенториальная
Волосковые
клетки мембрана
наружные
внутренние
каемка
Сосудистая
Ретикулярная
Клетки
Ген- Uq
el | |
зенаиКлауЛ |а
t
I
диуса
7
11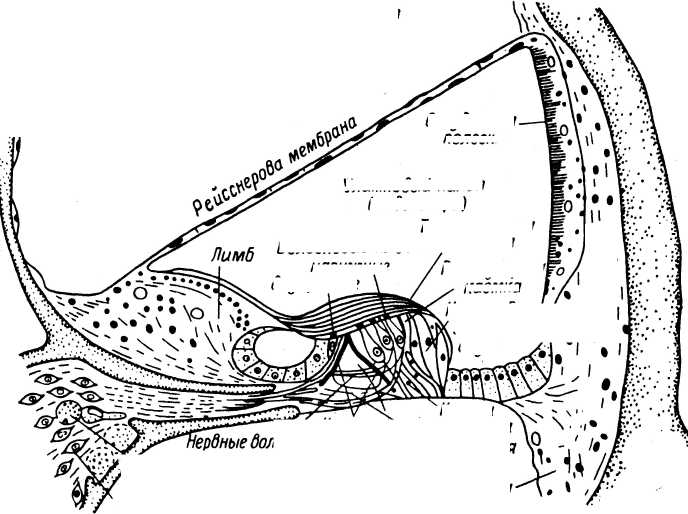











Клетки- Дейтерса стол^ы ' Базилярная
™Внутриганглионарные >Уннель мембрана
пугки t Спиральная
Спиральный ганглии связка
Барабанная лестница (перилимфа)
Рис. 16. Поперечный срез улиткового канала у морской свинки (по: Davis, 1957).
Схема среза на уровне второго (нижнего) завитка улитки.
в слуховых гребешках (кристах) полукружных каналов, в слуховых пятнах (макулах) саккулюса, утрикулюса и лагены. Волоски рецепторных клеток крист также, каки в органах боковой линии, покрыты веществом купу л. Последние, однако, более значительных размеров и более сложной формы. Макулы имеют отолитовую мембрану, содержащую кристаллы карбоната кальция. Купулы и отолиты погружены в эндолимфу — весьма специфическую жидкость (см. стр. 317), заполняющую полукружные каналы и мешочки преддверия. В целом следует признать, что вспомогательные структуры рецепторного аппарата вестибулярной системы имеют много сходства с аналогичными образованиями органов боковой линии. Орган слуха. В органе слуха вспомогательный аппарат организован значительно сложнее. К нему у высших позвоночных относятся: ушная раковина, которая у многих животных имеет специальную мускулатуру; наружный слуховой проход; барабанная перепонка; система косточек среднего уха и связанные с ней мышцы; наконец, многочисленные образования и жидкости внутреннего уха (основная мембрана, тпекториалъная мембрана, эндолимфа, перилимфа и др.). Подробное описание всех этих элементов и их свойств даны в обзорных работах (Bekesy, 1960; Винников, Титова, 1961; Винников, 1971; Молчанов, Радионова, 1972).
Волосковые рецепторы органа слуха локализованы в специальном образовании — органе Корти (рис. 16). Волоски рецепторных, клеток погружены в вещество тенториальной мембраны, которая омывается эндолимфой. Последняя характеризуется очень необычным ионным составом и рядом интересных физико-химических особенностей (Bekesy, 1952; С. A. Smith et al., 1954, 1958; Tasaki et al., 1954; см. также обзоры: Tasaki, 1957; Bekesy, 1960; Maggio, 1966; Bosher, Warren, 1968), на которых мы остановимся далее (см. гл. 7).
Афферентная и эфферентная иннервация сенсорных волосковых клеток
Иннервация органов чувств акустико-латеральной системы изучалась последние 10—15 лет довольно интенсивно (Rasmussen, 1946, 1953, 1960, 1964; Wersall, 1956, 1968; Engstrom, 1958, 1961; lurato, 1962, 1967; C. A. Smith, Rasmussen, 1963, 1968; Spoendlin, Gacek, 1963; Wersall et al., 1965; Винников, 1966, 1971; Лебедева, 1966; Spoendlin, 1966, 1968, 1973; Flock, 1967; G. A. Smith, 1967, 1968; Ross, 1971, и мн. др.). Хотя полной ясности в этом вопросе еще нет (например, не совсем понятна роль тонких немиелинизированных волокон, существование которых отмечалось рядом авторов, не до конца выяснена иннервация различных областей вестибулярного аппарата и органа слуха и т. д.), тем не менее уже сегодня можно представить себе общую картину связи волосковых механорецепторов с нервными волокнами.
Иннервация волосковых механорецепторных клеток несколько отличается у разных животных, а также в различных органах чувств. В кувшинообразных или колбообразных волосковых клетках I типа, встречающихся только в лабиринте (у пресмыкающихся и особенно у птиц и млекопитающих), имеется одно чашеобразное афферентное окончание, которое заключает в себе весь рецептор, за исключением его апикальной части (рис. 12). В ряде случаев в одной синаптической чаше могут находиться несколько рецепторных клеток. Цилиндрические рецепторные клетки Ц типа, которые имеются у всех позвоночных, на своей поверхности несут более многочисленные и более мелкие синапсы афферентных волокон. Эфферентные волокна, подходя к клеткам I типа, образуют синаптические окончания только на афферентных волокнах, с клетками же II типа эфферентные волокна контактируют непосредственно (рис. 12) (Wersall, 1956, 1968; Wersall et al., 1965).
Органы боковой линии. Механорецепторные структуры боковой линии могут снабжаться как мякотными, так и безмякотными волокнами (Murray, 1955; Flock, 1967; Harris, Flock, 1967; Ильинский и др., 1971). Толстые мякотные волокна, несущие афферентную функцию, при подходе к рецепторам теряют миелин и разветвляются, так что одно волокно иннервирует группу клеток (рис. 14). В органах боковой линии наряду с афферентными (негранулированными) синаптическими окончаниями на волосковых механорецепторах обнаружены гранулированные, заполненные большим количеством везикул окончания, которые, можно думать, принадлежат эфферентным нервным волокнам (Hama, 1962, 1965; Flock, 1965а). Эфферентные синапсы обнаружены только на рецепторных клетках. Они не встречаются на афферентных волокнах. Предполагается, что эфферентную функцию выполняют тонкие мякотные (Gorner, 1965; Russell, 1968) и не- миелинизированные (Harris, Flock, 1967) волокна. В отдельных случаях эфферентная иннервация волосковых клеток боковой линии может отсутствовать (Hama, 1962, 1965).
У ампулированных структур органов боковой линии имеются некоторые особенности. Каждая ампула Лоренцини иннервируется несколькими миелиновыми волокнами (до 30 у сильноэлектрического ската Torpedo). Синаптический аппарат сенсорных клеток имеет характерный вид лентообразной полоски (рис. 16, III). Один рецептор может иметь несколько синаптических контактов, а окончания одного нервного волокна могут разветвляться на нескольких рецептирующих клетках. Миелиновые волокна, подходящие к ампулам Лоренцини, имеют довольно большой диаметр; например, у слабоэлектрического ската Raja clavata максимум распределения диаметров лежит в области 12 мкм (Ильинский и др., 1971). Существенно то обстоятельство, что ампулярные каналы, берущие начало в разных участках тела, сходятся в определенных местах в краниальной части тела. Группа ампул окружена общей капсулой, куда и входит афферентный нерв. Таким образом, путь от рецепторов до центров у всех ампул данной группы оказывается одинаковым, что способствует возникновению высокосинхронизованного ответа в центральном отделе данной сенсорной системы.
Существенной морфологической особенностью рецепторов «особых» органов боковой линии является отсутствие у них эфферентной иннервации. Физиологический смысл этого обстоятельства в настоящее время еще не выяснен.
Вестибулярный аппарат. Основную группу волокон, идущих
да
о О И й и 5 « О ЕГ R![]()
![]()
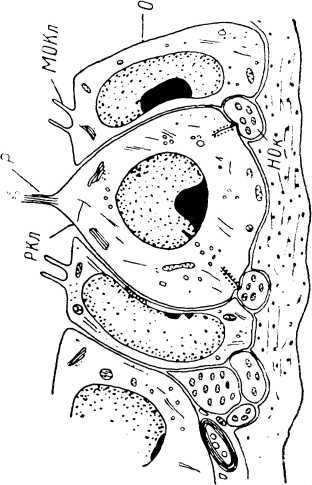
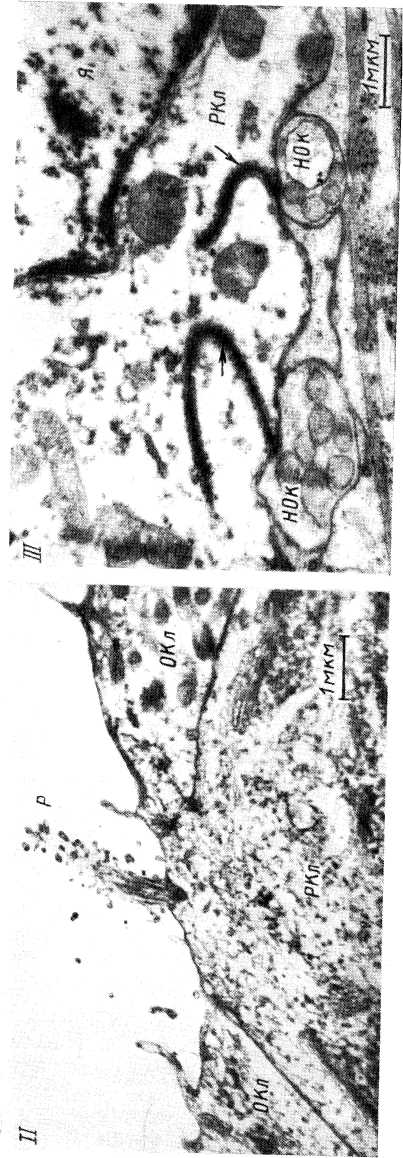
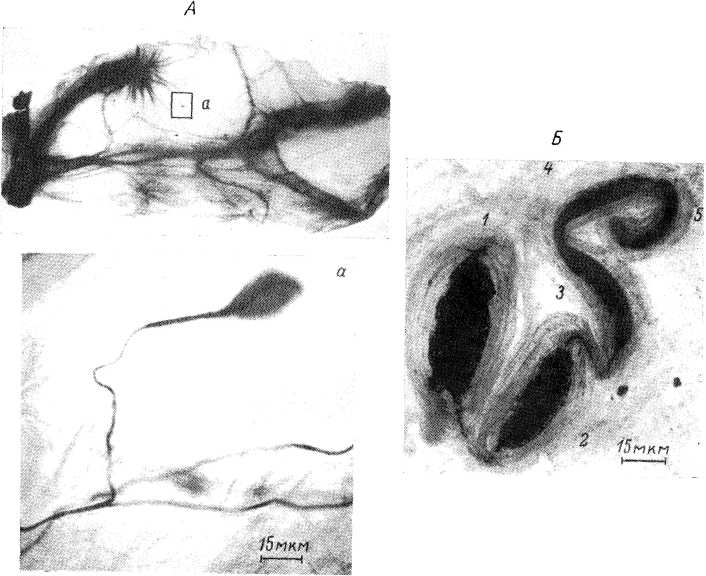
Рис. 108. Регенерация телец Пачини после реиннервации брыжейки толстого кишечника афферентным кожным нервом (и. saphenus) (Чалисова, Ильинский, 1974).
А — звездообразная неврома на конце пересаженного нерва, многочисленные вновь образованные тельца Пачини (форму развивающегося тельца Пачини см. на а); Б — пять (I—5) новообразованных рецепторов на одном нервном волокне. Импрегнация по Бильшовскому—Грос.
к вестибулярному аппарату, составляют мякотные волокна диаметром 1—15 мкм, хотя отмечается существование и более тонких (0.3—1 мкм) немиелинизированных волокон (Engstrom, Rexed, 1940; Rasmussen, 1940; Wersall, 1956; Блинков, Глезер, 1964). У рыб афферентные вестибулярные волокна теряют миелин вблизи самих синаптических контактов. У остальных позвоночных миелиновая оболочка исчезает раньше, и обнаженные аксоны идут между опорных клеток эпителия, которые выполняют функцию шванновских клеток (Wersall, 1956; Wersall et al., 1965). Клетки I и II типов вестибулярного аппарата могут иннервироваться как раздельными группами волокон, так и одним и тем же волокном. Одно и то же афферентное волокно может оканчиваться на нескольких рецепторных клетках, и на одном рецепторе может оканчиваться несколько различных волокон (Wersall, 1956). У вестибулярных клеюк млекопитающих обнаружены интересные особенности. Оказалось, чю между пре- и постсинаптическими мембранами этих клеток могут существовать очень тесные взаимосвязи, которые позволяют предположить существование электрической, эфаптической передачи (Spoendlin, 1966; Flock, 1971а).
Наличие эфферентных волокон, направляющихся к рецепторам вестибулярного аппарата, было показано рядом авторов (Petroff, 1955; Dohlman et al., 1958; Rasmussen, Gacek, 1958; Gacek, 1960; Altman, Carpenter, 1961; Ireland, Farkashidy, 1961; Gacek et al., 1965; Hillman, 1969; Gribenski, 1970; см. также обзоры: Кисляков и др., 1972; lurato et al., 1972). Согласно Росси (Rossi, Cortesina, 1963, 1965; Rossi, 1968), существуют прямой дорзальный и прямой вентральный эфферентные вестибулярные пучки, которые иннервируют исключительно вестибулярные рецепторы, а также прямой ретикуло-кохлеарно-вестибулярный пучок, волокна которого связаны с рецепторами как органа слуха, так и вестибулярного аппарата. В вестибулярном нерве эфферентные волокна образуют компактный пучок, лежащий рядом и дорзальнее эфферентного кохлеарного пучка (Gacek, 1968).
Существование эфферентных синапсов (рис. 12) на волосковых механорецепторах (телах клеток II типа) и на отходящих (от клеток I типа) афферентных волокнах (аксо-дендритные синапсы) было продемонстрировано с помощью электронного микроскопа (Wersall, 1956, 1967, 1968; Engstrom, 1958, 1961; Engstrom, Wersall, 1958a, 1958b; Wersall et al., 1965).
Количество эфферентных волокон в вестибулярном аппарате сравнительно невелико: порядка 400 при общем числе волокон вестибулярного нерва у кошки до 12 000 (Gacek et al., 1965); диаметр их у кошки близок к размеру афферентных волокон и равен 2—4 мкм (Gacek, 1960). Пресинаптические же их разветвления достаточно тонки (0.18—0.3 мкм) и образуют своего рода горизонтальное нервное сплетение (С. A. Smith, Rasmussen, 1968). Небольшое число эфферентных волокон, по-видимому, компенсируется их значительным разветвлением на пути к рецепторам. В итоге Одно волокно может $ыть в контакте с несколькими афферентным# волокнами и волосковыми клетками. Однако это не означает, что все вестибулярные рецепторы имеют эфферентную иннервацию.
Орган cijyxa. Нервные волокна органа слуха довольно многочисленны (порядка 50 000 у кошки) и относительно небольшого диаметра: 1—8 мкм (Gacek, Rasmussen, 1961; Лебедева, 1966). У кошки основная масса афферентных волокон идет к внутренним волосковым клеткам и только около 5000 или даже менее того (всего лишь до 5% от общего числа) идет к наружным волосковым рецепторам (Spoendlin, 1969, 1971). Если учесть, что у кошки количество внутренних сенсорных клеток составляет примерно 2600, а наружных — примерно 9900 (Retzius, 1884; Engstrom, Wersall, 1958а, 1958b; Винников, Титова, 1961), то понятно, что на каждом внутреннем рецепторном элементе оканчивается большое число (10—20) афферентных нервных волокон. Следует отметить, что афферентные волокна, иннервирующие внутренние клетки, практически совсем не ветвятся и имеют окончания на одном (самое большее 2—3) рецепторе. Напротив, афферентные волокна, связанные с наружными волосковыми клетками, сильно разветвляются (каждое волокно дает по своему ходу коллатерали приблизительно на протяжении 0.6 мм). В итоге каждое волокно оканчивается на большом числе рецепторов (Spoendlin, 1966, 1968, 1973). Структура синаптических контактов афферентных волокон с рецепторами органа слуха принципиально сходна с аналогичными образованиями других органов чувств акустико-латеральной системы (Wersall et al., 1965).
Несомненно, что эфферентная иннервация рецепторных элементов органа слуха является наиболее сложно организованной по сравнению с другими органами чувств. Общее количество эфферентных волокон у кошки сравнительно невелико — порядка 500. Однако они сильно ветвятся, благодаря чему общее число эфферентных сигналов лишь несколько меньше количества афферентных окончаний. Поверхность же их контакта с рецепторами может быть даже несколько больше, чем у афферентных структур (С. A. Smith, Rasmussen, 1963; Rasmussen, 1964; Spoendlin, 1966, 1973).
В области внутренних волосковых клеток эфферентные волокна образуют контакты почти исключительно с отходящими афферентными волокнами, число их синаптических связей незначительно, а размеры синаптических бляшек невелики. Напротив, в области наружных волосковых клеток эфферентные волокна контактируют непосредственно с рецепторными элементами, а размеры и число синаптических контактов значительны. При этом наиболее многочисленные и большие синаптические окончания эфферентные нервы образуют на наружных волосковых клетках базального витка улитки. Вверх по улитке число и размеры эфферентных синапсов уменьшаются (Wersall et al., 1965; Spoendlin, 1968).
♦ ♦ ♦
Таким образом, видно, что у всех механорецепторных образований органов чувств собственно рецептирующим элементом является специальная волосковая чувствительная клетка, которая имеет синаптические контакты с афферентными нервными волокнами сенсорных нейронов. Следовательно, механорецепторы органов чувств должны быть отнесены к вторичным сенсорным структурам. Основное различие между разными механорецепторными приборами органов чувств заключается в строении их вспомогательного аппарата.