2.3.2. Регионы соматосенсорной коры
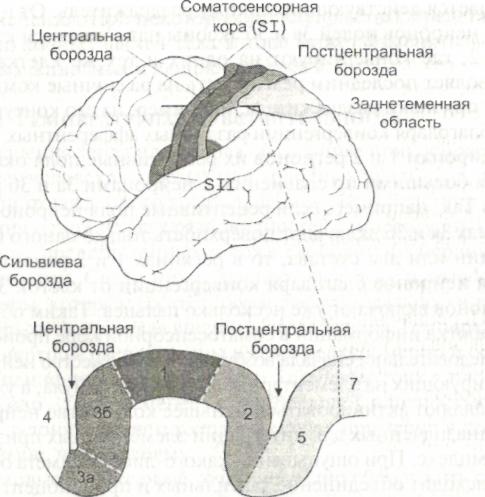
Первоначальная схема Пенфилда была уточнена в ходе исследований коры с помощью точечных микроэлектродов, которые позволяли регистрировать активность отдельных нейронов при раздражении ограниченных участков поверхности тела или стимуляции афферентных нейронов, передающих информацию от проприоцепторов. Применение такой техники исследований привело к представлению о существовании функционально различающихся регионов соматосенсорной коры. Эти регионы соответствуют За, 36, 1 и 2 полям Бродмана (рис. 2.6). В поле За поступает информация преимущественно от проприоцепторов мышечных веретен, сухожилий и суставных сумок, в поле 36 — от механорецеп-торов кожи, причем и в том и в другом случае она содержит только элементарные характеристики действующих стимулов. Дальнейшая переработка информации, доставленной от рецепторов кожи, происходит в поле 1, а в поле 2 переработанные сигналы комбинируются с информацией, поступившей от проприоцепторов. Таким образом, если в полях За и 36 формируются только элементарные ощущения действующих раздражителей, то в полях 1 и 2 создаются комплексные представления.
Все четыре поля получают информацию от одной и той же поверхности тела, но в каждом из полей одно ощущение доминирует над остальными. В поле За преобладает вход от интрафузальных рецепторов, в поле 36 — от поверхностных 1рецепторов кожи, в поле 2 — от рецепторов, реагирующих на сильное давление, а в поле 1 — от быстро адаптирующихся рецепторов кожи. В полях За и 36 отсутствуют нейроны, воспринимающие информацию о направлении действия стимула; они содержатся в полях 1 и 2, где определяется трехмерная форма предмета и направление, в котором пере мешается действующий на кожу раздражитель. От некоторых нейронов полей За и 36 аксоны направляются к полям 1 и 2, где конвергируют на одних и тех же клетках, что позволяет последним реагировать на различные комплексные признаки раздражителя, например, на его контур.
Благодаря конвергенции различных афферентных входов к нейронам 1 и 2 регионов их рецептивные поля оказываются большими по сравнению с нейронами За и 36 регионов. Так, например, если рецептивные поля нейронов в регионах За и 36 включают поверхность только одного пальца и один или два сустава, то в регионах 1 и 2 рецептивные поля нейронов благодаря конвергенции от клеток За и 36 регионов включают уже несколько пальцев. Таким образом, обработка информации в соматосенсорной коре происходит последовательно: сначала возбуждается множество нейронов, реагирующих на элементарные признаки стимула, а уже они заставляют активироваться меньшее количество нейронов, предназначенных для интеграции элементарных признаков в комплекс. При ощупывании какого-либо предмета обычно происходит объединение тактильных и проприоцептивных ощущений, то есть сведений о характере поверхности и ее трехмерной форме. Если ощупываемый предмет хорошо знаком человеку (например, монета, расческа, часы), то испытуемый его называет, основываясь на сопоставлении сенсорных ощущений с имеющимся у него опытом. Способность создавать комплексное представление о предмете основана на т.н. стереогностическом чувстве.
К
Рис. 2.6. Регионы соматосенсорной коры в области представительства руки
Опознание предметов на ощупь основано на использовании афферентной информации от тактильных рецепторов (поступает в регион 36) и рецепторов интрафузальных волокон (поступает в регион За). В этих регионах происходит элементарная обработка информации, в которой участвуют простые нейроны коры. Следующий этап переработки информации осуществляется в регионах 1 и 2, где имеются комплексные нейроны, получающие информацию из регионов За и 36 и способные объединять несколько элементарных признаков. Целостное восприятие действующего стимула происходит при участии регионов 5 и 7 заднетеменной области.
летки коры, специализирующиеся на переработке однородной информации, объединены в кортикальные колонки, диаметр которых составляет приблизительно 0,2—0,5 мм. Аксоны таламических нейронов, передающих сигналы к коре, в основном заканчиваются в пределах одной колонки, что позволяет ей получать обособленную часть информации и играть роль элементарной функциональной единицы. Принципиальная схема соединений между нейронами разных уровней, обеспечивающая соматопическое представительство в коре того или иного участка тела, определяется генетически, однако индивидуальный сенсорный опыт, в особенности приобретаемый в раннем возрасте, способен вносить в ее изменения. При некоторых обстоятельствах представи-ельство разных частей тела в коре может изменяться и у зрослых животных и людей.