

2) Распад пуриновых оснований
Наиболее активно катаболизм пуринов идет в печени, тонком кишечнике (пище-вые пурины) и почках. Распад пуринов начинается с отщепления 5’-фосфатной груп-пы от АМФ или ГМФ. Это осуществляет фермент 5’-нкуклеотидаза, локализованный на цитоплазматической мембране. Образующиеся продукты (аденозин и гуанозин) включаются в ряд простых реакций, результатом которых является образование об-щего промежуточного продукта ксантина (рис.8). И далее ксантин под воздействием ксантиноксидазы превращается в конечный продукт катаболизма пуринов – мочевую кислоту. Следует оговориться, что конечным продуктом мочевая кислота является только у человека, приматов, птиц и большинства млекопитающих. У остальных жи-вых организмов мочевая кислота превращается в аллантоин.
Около 18-20% мочевой кислоты удаляется с желчью через кишечник, где она де-градируется микрофлорой до CO2 и воды. Остальная часть удаляется через почки.
3) Регуляция синтеза белка
Работа генов в любом организме – прокариотическом, эукариотическом, одноклеточном или многоклеточном – контролируется и координируется.
Различные гены обладают неодинаковой временной активностью. Одни из них характеризуются постоянной активностью. Такие гены отвечают за синтез белков, необходимых клетке или организму на протяжении всей жизни, например, гены, продукты которых участвуют в синтезе АТФ. Большинство же генов обладает непостоянной активностью, они работают только в определенные моменты, когда появляется необходимость в их продуктах – белках. Гены различаются и по уровням своей активности (низкий или высокий).
Белки клетки классифицируются как регуляторные и структурные. Регуляторные белки синтезируются на регуляторных генах и контролируют работу структурных генов. Структурные гены кодируют структурные белки, выполняющие структурную, ферментативную, транспортную и другие функции (кроме регуляторной!).
Регуляция синтеза белка осуществляется на всех этапах этого процесса: транскрипции, трансляции и посттрансляционной модификации либо путем индукции, либо путем репрессии.
Регуляция активности генов эукариотических организмов намного сложнее регуляции экспрессии генов прокариот, что определяется сложностью организации эукариотического организма и особенно многоклеточного. В 1961 г. французские ученые Ф. Жакоб, Ж. Моно и А. Львов сформулировали модель генетического контроля синтеза белков, катализирующих усвоение клеткой лактозы – концепцию оперона.
Опероном называют группу генов, работа которых контролируется одним геном-регулятором.
Ген-регулятор – это ген, обладающий постоянной низкой активностью, на нем синтезируется белок-репрессор – регуляторный белок, который может соединяться с оператором, инактивируя его.
Оператор – точка начала считывания генетической информации, он управляет работой структурных генов.
В структурных генах лактозного оперона содержится информация о ферментах, участвующих в метаболизме лактозы. Поэтому лактоза будет служить индуктором – агентом, инициирующим работу oпepoна.
Промотор – место прикрепления РНК-полимеразы.
Терминатор – место окончания синтеза иРНК.
При отсутствии индуктора система не функционирует, поскольку "свободный" от индуктора – лактозы – репрессор соединен с оператором. В этом случае фермент РНК-полимераза не может катализировать процесс синтеза иРНК. Если в клетке оказывается лактоза (индуктор), она, взаимодействуя с репрессором, изменяет его структуру, в результате чего репрессор освобождает оператор. РНК-полимераза соединяется с промотором, начинается синтез иРНК (транскрипция структурных генов). Затем на рибосомах формируются белки по программе иРНК-лактозного оперона. У прокариотических организмов на одну молекулу иРНК переписывается информация со всех структурных генов оперона, т.е. оперон – это единица транскрипции. Транскрипция продолжается до тех пор, пока в цитоплазме клетки сохраняются молекулы лактозы. Как только все молекулы будут клеткой переработаны, репрессор закрывает оператор, синтез иРНК прекращается.
Таким образом, синтез иРНК и, соответственно, синтез белка должны строго регулироваться, поскольку у клетки недостаточно ресурсов для одновременной транскрипции и трансляции всех структурных генов. И про-, и эукариоты постоянно синтезируют только те иРНК, которые необходимы для выполнения основных клеточных функций Экспрессия остальных структурных генов осуществляется под строгим контролем регуляторных систем, запускающих транскрипцию только в том случае, когда возникает потребность в определенном белке (белках).
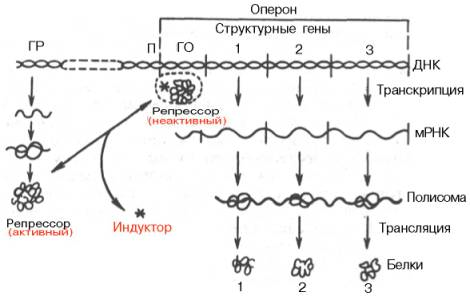 индукция
индукция
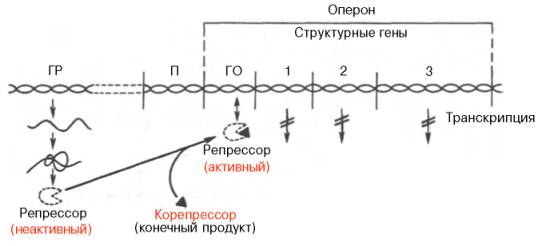 репрессия
репрессия