

Строение[
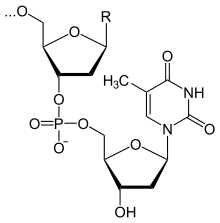
Фрагмент полимерной цепочки ДНК
Полимерные формы нуклеиновых кислот называют полинуклеотидами. Цепочки из нуклеотидов соединяются через остаток фосфорной кислоты (фосфодиэфирная связь). Поскольку в нуклеотидах существует только два типа гетероциклических молекул, рибоза и дезоксирибоза, то и имеется лишь два вида нуклеиновых кислот — дезоксирибонуклеиновая (ДНК) ирибонуклеиновая (РНК).
Мономерные формы также встречаются в клетках и играют важную роль в процессах передачи сигналов или запасании энергии. Наиболее известный мономер РНК — АТФ, аденозинтрифосфорная кислота, важнейший аккумулятор энергии в клетке.
ДНК и РНК
ДНК (дезоксирибонуклеиновая кислота). Сахар — дезоксирибоза, азотистые основания:пуриновые — гуанин (G), аденин (A), пиримидиновые — тимин (T) и цитозин (C). ДНК часто состоит из двух полинуклеотидных цепей, направленных антипараллельно.
РНК (рибонуклеиновая кислота). Сахар — рибоза, азотистые основания: пуриновые — гуанин (G), аденин (A), пиримидиновыеурацил (U) и цитозин (C). Структура полинуклеотидной цепочки аналогична таковой в ДНК. Из-за особенностей рибозы молекулы РНК часто имеют различные вторичные и третичные структуры, образуя комплементарные участки между разными цепями.
Типы рнк
Ма́тричная рибонуклеи́новая кислота́ (мРНК, синоним — информацио́нная РНК, иРНК) — РНК, содержащая информацию опервичной структуре (аминокислотной последовательности) белков[8]. мРНК синтезируется на основе ДНК в ходе транскрипции, после чего, в свою очередь, используется в ходе трансляции как матрица для синтеза белков. Тем самым мРНК играет важную роль в «проявлении» (экспрессии) генов.Рибосо́мные рибонуклеи́новые кисло́ты (рРНК) — несколько молекул РНК, составляющих основу рибосомы. Основной функцией рРНК является осуществление процесса трансляции — считывания информации с мРНК при помощи адапторных молекул тРНК и катализ образования пептидных связей между присоединёнными к тРНК аминокислотами.
Транспортная РНК, тРНК — рибонуклеиновая кислота, функцией которой является транспортировка аминокислот к месту синтеза белка. Имеет типичную длину от 73 до 93 нуклеотидов и размеры около 5 нм. тРНК также принимают непосредственное участие в наращивании полипептидной цепи, присоединяясь — будучи в комплексе с аминокислотой — к кодону мРНК и обеспечивая необходимую для образования новой пептидной связи конформацию комплекса.
Для каждой аминокислоты существует своя тРНК.тРНК является одноцепочечной РНК, однако в функциональной форме имеет конформацию «клеверного листа». Аминокислота ковалентно присоединяется к 3'-концу молекулы с помощью специфичного для каждого типа тРНК фермента аминоацил-тРНК-синтетазы. На участке C находится антикодон, соответствующий аминокислоте.
12. Хлоропласты. Строение. Химический состав, Функции.
. Весь процесс фотосинтеза протекает в зеленых пластидах - хлоропластах. Различают три вида пластид: лейкопласты - бесцветные, хромопласты - оранжевые, хлоропласты - зеленые. Именно в хлоропластах сосредоточен зеленый пигмент хлорофилл. Незеленые растения, например грибы, лишены пластид. Эти растения не обладают способностью к фотосинтезу. В процессе эволюции дифференциация пластид произошла очень рано. Правда, у фотосинтезирующих бактерий и сине-зеленых водорослей пластид еще нет, их роль выполняет окрашенная часть протоплазмы, прилегающая к оболочке. Это наиболее примитивная организация фотосинтетического аппарата. Однако уже у водорослей имеются специальные образования (хроматофоры), в которых сосредоточены пигменты, они разнообразны по форме (спиральные, ленточные, в виде пластинок или звезд). Высшие растения характеризуются вполне сформировавшимся типом пластид в форме диска или двояковыпуклой линзы. Приняв форму диска, хлоропласты становятся универсальным аппаратом фотосинтеза.
Химический состав хлоропластов достаточно сложен и характеризуется высоким (75 %) содержанием воды. Около 75—80 % общего количества сухих веществ приходится на долю различных органических соединений, 20—25 % — на долю минеральных веществ. Структурной основой хлоропластов являются белки, содержание которых достигает 50—55 % сухой массы, примерно половина из них водорастворимые. Такое высокое содержание белков объясняется их многообразными функциями в составе хлоропластов. Это структурные белки, являющиеся основой мембран, белки-ферменты, транспортные белки, поддерживающие определенный ионный состав, отличающийся от цитозоля, сократительные белки, подобные актомиозину мышц, которые обеспечивают двигательную активность хлоропластов. Белки выполняют также рецепторную функцию, принимая участие в регуляции интенсивности фотосинтеза в меняющихся условиях внутренней и внешней среды.
Важнейшей составной частью хлоропластов являются липиды, содержание которых колеблется от 30 до 40 % сухой массы. Липиды хлоропластов представлены тремя группами соединений.
Углеводы не являются конституционными веществами хлоропласта. В очень небольших количествах фосфорные эфиры сахаров участвуют в восстановительном цикле углерода, в основном же это продукты фотосинтеза. Поэтому содержание углеводов в хлоропластах колеблется значительно (от 5 до 50 %). В активно функционирующих хлоропластах углеводы обычно не накапливаются, происходит их быстрый отток. При уменьшении потребности в продуктах фотосинтеза в хлоропластах образуются крупные крахмальные зерна. В этом случае содержание крахмала может возрасти до 50 % сухой массы и активность хлоропластов снизится.
В хлоропластах высокое содержание минеральных веществ. Сами хлоропласты составляют 25—30 % массы листа, но в них сосредоточено до 80 % железа, 70—72 % — магния и цинка, около 50 % — меди, 60 % кальция, содержащихся в тканях листа. Эти данные хорошо согласуются с высокой и разнообразной ферментативной активностью хлоропластов. Минеральные элементы выступают в роли простетических групп и кофакторов деятельности ферментов. Магний входит в состав хлорофилла. Важная роль кальция заключается в стабилизации мембранных структур хлоропластов.
Строение хлоропласта
Строение хлоропласта, наблюдаемое с помощью электронного микроскопа, весьма сложное. Подобно ядру и митохондриям хлоропласт окружен оболочкой, состоящей из двух липопротеидных мембран. Внутреннюю среду представляет относительно однородная субстанция — матрикс, илистрома, которую пронизывают мембраны — ламеллы. Ламеллы, соединенные друг с другом, образуют пузырьки — тилакоиды. Плотно прилегая друг к другу, тилакоиды образуют граны,которые различают даже под световым микроскопом. В свою очередь, граны в одном или нескольких местах объединены друг с другом с помощью межгранных тяжей — тилакоидов стромы. Пигменты хлоропласта, участвующие в улавливании световой энергии, а также ферменты, необходимые для световой фазы фотосинтеза, вмонтированы в мембраны тилакоидов.
 Рис.1.
Строение хлоропласта
Рис.1.
Строение хлоропласта
1 — внешняя мембрана; 2 — внутренняя мембрана; 3 — крахмальное зерно; 4 — ДНК; 5 — тилакоиды стромы (фреты); 6 — тилакоид граны; 7 — матрикс (строма)
Строение зрелых хлоропластов одинаково у всех высших растений, как и в клетках разных органов одного растения (листьях, зеленеющих корнях, коре, плодах). В зависимости от функциональной нагрузки клеток, физиологического состояния хлоропластов, их возраста различают степень их внутренней структурированности: размеры, количество гран, связь между ними. Так, в замыкающих клетках устьиц основная функция хлоропластов — фоторегуляция устьичных движений. Этот процесс обеспечивается энергией высокоструктурированными митохондриями. Хлоропласты содержат крупные крахмальные зерна, набухшие тилакоиды, липофильные глобулы, что свидетельствует об их низкой энергетической нагрузке.
С возрастом строение хлоропластов существенно меняется. Молодые хлоропласты характеризуются ламеллярной структурой, в таком состоянии хлоропласты способны размножаться делением. В зрелых хлоропластах хорошо выражена система гран. В стареющих хлоропластах происходит разрыв тилакоидов стромы, связь между гранами уменьшается, в дальнейшем наблюдаются распад хлорофилла и деструкция гран. В осенней листве деградация хлоропластов приводит к образованию хромопластов, в которых каротиноиды сосредоточены в пластоглобулах.
Физиологические особенности хлоропластов
Важным свойством хлоропластов является их способность к движению. Хлоропласты передвигаются не только вместе с цитоплазмой, но способны и самопроизвольно изменять свое положение в клетке. Скорость движения хлоролластов составляет около 0,12 мкм/с. Хлоропласты могут быть распределены в клетке равномерно, однако чаще они скапливаются около ядра и вблизи клеточных стенок. Большое значение для расположения хлоропластов в клетке имеют направление и интенсивность освещения. При малой интенсивности освещения Хлоропласты становятся перпендикулярно к падающим лучам, что является приспособлением к лучшему их улавливанию. При высокой освещенности хлоропласты передвигаются к боковым стенкам и поворачиваются ребром к падающим лучам. В зависимости от освещения может также меняться и форма хлоропластов. При более высокой интенсивности света их форма становится ближе к сферической.
Основная функция хлоропластов - это процесс фотосинтеза. В 1955 г. Д. Арнон показал, что в изолированных хлоронпластах может быть осуществлен весь процесс фотосинтеза. Важно отметить, что хлоропласты имеются не только в клетках листа. Они встречаются в клетках не специализирующихся на фотосинтезе органов: в стеблях, колосковых чешуйках и остях колосьев, корнеплодах, клубнях картофеля и т. д. В ряде случаев зеленые пластиды обнаруживаются в тканях, расположенных не в наружных, освещенных частях растений, а в слоях, удаленных от света, в тканях центрального цилиндра стебля, в средней части луковицы лилейных, а также в клетках зародыша семени многих покрытосеменных растений. Последнее явление (хлорофиллоносность зародыша) привлекает внимание систематиков растений. Имеются предложения разделить все покрытосеменные растения на две большие группы: хлороомбриофиты и лейкоэмбриофиты, т. е. содержащие и не содержащие хлоропласты в зародыше (Яковлев). Исследования показали, что структура хлоропластов, расположенных в других органах растения, так же как и состав пигментов, сходны с хлоропластами листа. Это дает основания считать, что они способны к фотосинтезу.
В том случае, если они подвергаются освещению, по-видимому, в них действительно происходит фотосинтез. Так, фотосинтез хлоропластов, расположенных в остях колоса, может составлять около 30% от общего фотосинтеза растения. Позеленевшие на свету корни способны к фотосинтезу. В хлоропластах, находящихся, в кожуре плода до определенного этапа его развития, также может идти фотосинтез. Согласно предположению А. Л. Курсанова, хлоропласты, расположенные вблизи проводящих путей, выделяя кислород, способствуют повышению интенсивности обмена веществ ситовидных трубок. Вместе с тем роль хлоропластов не ограничивается их способностью к фотосинтезу. В определенных случаях они могут служить источником питательных веществ (Е. Р. Гюббенет). Хлоропласты содержат большее количествовитаминов, ферментов и даже фитогормонов (в частности, гиббереллина). В условиях, при которых ассимиляция исключена, зеленые пластиды могут играть активную роль в процессах обмена веществ.