

Agadzhanyan T
.1.pdfслабление. Источником энергии в медленных волокнах является аэробное дыхание. При недостатке кислорода мышцы продолжают работать за счет анаэробного гликолиза, в этом случае образуется молочная кислота и создается кислородная задолженность. По мере окисления дыхательного субстрата мобилизуются резервные углеводы и жиры. Медленные волокна обеспечивают длительное сокращение мышцы, поддержание позы.
Быстрые (фазические, белые) мышечные волокна располагаются в поверхностных слоях мышц конечностей. Они белого цвета, так как миоглобина и цитохромовых пигментов в них мало. Клетки быстрых мышечных волокон содержат мало митохондрий, саркоплазматический ретикулум хорошо развит, имеется множество гликогеновых гранул. Быстрые мышечные волокна иннервируются толстыми нервными волокнами 10–20 мкм в диаметре. Обычно на одном мышечном волокне одна или две концевых пластинки. Скорость проведения импульса составляет 8–40 м/с. Наблюдается быстрое сокращение (в 3 раза быстрее, чем у медленных волокон) и довольно быстрое утомление. Источником АТФ служат анаэробные процессы (гликолиз), быстро создается кислородная задолженность. В качестве энергетического субстрата интенсивно используется гликоген.
Другой способ классификации волокон основан на том, что они различаются по своим сократительным и метаболическим особенностям: в зависимости от скорости укорочения – на быстрые и медленные; в зависимости от главного пути образования АТФ – на оксидативные и гликолитические.
На основании этих характеристик можно выделить
3типа волокон:
1)медленные оксидативные (тип I) – низкая актив-
ность миозиновой АТФ-азы и высокая окислительная способность;
2)быстрые оксидативные (тип IIа) – высокая актив-
ность миозиновой АТФ-азы и высокая окислительная способность;
69

3) быстрые гликолитические (тип IIб) – высокая ак-
тивность миозиновой АТФ-азы и высокая гликолитическая способность.
Четвертый возможный вариант – медленные гликолитические волокна в настоящее время еще не обнаружены.
Рис. 2.22. Двигательная единица. Строение нервной клетки (мотонейрона):
1 – ядро; 2 – ядрышко; 3 – сателлит ядрышка; 4 – дендрит; 5 – эндоплазматическая щель; 6 – синаптическое окончание; 7 – ножка астроцита; 8 – гранулы ДНК; 9 – липофусцин; 10 – аппарат Гольджи; 11 – митохондрия; 12 – аксонный холмик; 13 – нейрофибриллы; 14 – аксон; 15 – миелиновая оболочка; 16 – перехват Ранвье; 17 – ядро шванновской клетки; 18 – шванновская клетка в области нервно-мышечного синапса; 19 – ядро
мышечной клетки; 20 – нервно-мышечное соединение; 21 – мышца (Дж. Шаде, Д. Форд, 1976)
70
Иннервация скелетных мышц. Двигательные нейроны
(мотонейроны) – передают импульсы от центральной нервной системы к эффекторам. К каждой мышце подходит много двигательных нейронов, которые ветвятся и иннервируют группы мышечных волокон. Каждая такая группа вместе с иннервирующим ее мотонейроном называется двигательной единицей (ДЕ). При стимуляции мотонейрона все мышечные волокна, к которым подходят веточки его аксона, сокращаются одновременно. Число волокон в двигательной единице зависит от того, насколько сложным должно быть управление данной мышцей. Например, в глазодвигательной мышце оно составляет около 10, а бицепсе – более 1000. В месте контакта двигательного аксона с мышечным волокном формируется нервно-мышечное соединение, или двигательная концевая пластинка (рис. 2.22). Здесь аксон не имеет миелиновой оболочки, а его ветвящиеся окончания погружены в бороздки концевой пластинки.
Механика мышечного сокращения
Скелетные мышцы передают усилие частям скелета посредством сухожилий. Во время развития силы у мышцы есть тенденция укоротиться, т.е. растянуть упругие структуры, прикрепляющие ее к скелету. Сила, развиваемая мышцей, соответствует сумме сил отдельных волокон. Чем толще мышца и больше «физиологическая» площадь ее поперечного сечения (сумма площадей поперечных сечений отдельных волокон), тем она сильнее. При мышечной гипертрофии ее сила и толщина волокон возрастают в одинаковой степени.
Количество поперечных мостиков, связывающих актиновые и миозиновые филаменты (следовательно, и развиваемая сила), зависит от степени перекрывания толстых и тонких нитей, а значит и от длины саркомера или мышцы.
Типы и виды сокращения мышц. Различают следую-
щие типы (режимы) сокращения скелетной мышцы: изометрический (изменяется только тонус), изотонический (изме-
71
няется только длина) и ауксотонический (изменяются длина и тонус).
Изометрический режим проявляется в том, что в мышце во время ее активности нарастает напряжение (генерируется сила), но она не может укорачиваться (например, при попытке поднять слишком тяжелый предмет). При изотоническом режиме мышца укорачивается при неизменном внутреннем напряжении (например, при небольшой величине поднимаемого груза), развивая силу, способную поднять данный груз, она укорачивается (уменьшает свою длину), сохраняя напряжение, равное весу поднимаемого груза. Так как при изотоническом сокращении есть элементы изометрического сокращения (в самом начале сокращения мышцы), а при изометрическом сокращении наблюдаются элементы смещения (укорочения), то, в сущности, любое сокращение является смешанным (ауксотоническим), в котором преобладает либо изотонический, либо изометрический элемент.
Различают два основных вида мышечных сокращений: одиночные и тетанические сокращения. При непосредственном раздражении мышцы (прямое раздражение) или опосредованно через иннервирующий ее двигательный нерв (непрямое раздражение) одиночным стимулом возникает одиночное мышечное сокращение, в котором выделяют три фазы: латентный период – время от начала действия раздражителя до начала ответной реакции; фаза сокращения (фаза укорочения); фаза расслабления.
Одиночное сокращение возникает при действии на мышцу порогового раздражения. Реакция мышцы (генерация ПД и сокращение) начинается не сразу, а через 2–2,5 мс – латентный период (время от нанесения раздражения до начала генерации ПД). ПД скелетной мышцы длится 3–5 мс, и на фоне его развития начинается мышечное сокращение. Одиночный двигательный акт (одиночное сокращение) состоит из фазы сокращения, которая продолжается около 50 мс, и фазы расслабления, которая длится более 50 мс. Возбудимость мышцы во время одиночного сокращения изменяется. В начале сокращения (во время развития ПД) возникает фаза
72

абсолютной рефрактерности (длительность 3–5 мс), затем фаза относительной рефрактерности, а после нее фаза повышенной возбудимости (рис. 2.23). Длительность этих периодов зависит от функционального состояния мышцы.
Рис. 2.23. Миограмма одиночного сокращения мышцы лягушки и суммация сокращений (запись на фаль-аппарате)
Для А: а – одиночное сокращение (1 – отметка раздражения, 2 – латентный период, 3 – фаза сокращения, 4 – фаза расслабления);
б – отметка времени 0,01 с.
Для Б – суммация сокращений: I – неполная и II – полная суммация: 1 – одиночное мышечное сокращение при воздействии первого стимула; 2 – одиночное мышечное сокращение при воздействии второго стимула; 3 – кривая суммации – сокращения, возникающего в ответ на одновременное раздражение двумя стимулами
Суммация сокращений возникает в том случае, если на мышцу наносится два и более пороговых раздражений с периодом меньше длительности одиночного сокращения (около 100 мс). Если второе раздражение наносится на мышцу в
73
период, когда она не полностью расслаблена, то наблюдается слияние двух сокращений (суммация), однако в этом случае не происходит полного слияния первого и второго сокращений (не полная суммация) и на миограмме регистрируются две вершины (см. рис. 2.23). Мышца суммирует следующие друг за другом процессы, лежащие в основе последовательных сократительных актов (свойство суперпозиции). Если интервал между стимулами меньше 50 мс, то второе раздражение воздействует на мышцу в конечный период фазы сокращения и происходит полное слияние двух сокращений (полная суммация), а на миограмме регистрируется кривая с одной вершиной. В обоих случаях (неполная и полная суммация) амплитуда сокращения на второе раздражение будет больше, чем на первое, хотя величина первого и второго стимулов одинакова. Это связано с тем, что в ответ на пороговый стимул сокращаются не все мышечные волокна данной мышцы, а та их часть, которая имеет минимальный порог возбуждения (в них происходит генерация ПД). В остальных волокнах возникают лишь локальные ответы, если второй стимул дается через короткий интервал времени, то локальные ответы на первый и второй стимул суммируются,
иПД возникают в ранее не возбужденных волокнах.
Вестественных условиях к скелетной мышце из ЦНС поступают не одиночные импульсы, а серия импульсов, следующих друг за другом с определенными интервалами, на которые мышца отвечает длительным сокращением. Такое длительное сокращение мышцы, возникающее в ответ на ритмическое раздражение, называется тетаническим сокращением или тетанусом.
Тетанус – длительное сокращение (напряжение) мышцы, возникающее в ответ на ритмическое раздражение двигательного нерва или мышцы. При этом происходит слияние (суммация) и наложение (суперпозиция) одиночных сокращений (напряжений). Если каждый последующий импульс возбуждения поступает к мышце в тот период, когда она находится в фазе укорочения, то возникает гладкий тетанус, а если в фазу расслабления – зубчатый тетанус (рис. 2.24).
74
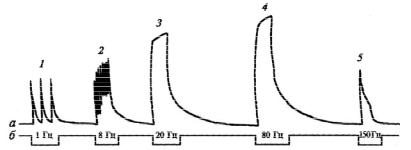
Рис. 2.24. Виды тетануса. Запись сокращений икроножной мышцы лягушки (а) при различной частоте раздражений (б):
1 – одиночные сокращения; 2 – зубчатый тетанус; 3 – гладкий тетанус; 4 – оптимум; 5 – пессимум сокращений
Виды тетануса: зубчатый и гладкий. Зубчатый (не-
полный) тетанус возникает при редкой стимуляции (около 10–15 стим./с), когда каждый последующий импульс поступает к мышце в фазу расслабления. Гладкий (сплошной) тетанус возникает при более частой (выше 20–25 стим./с) стимуляции, когда каждый последующий импульс поступает в фазу сокращения.
Амплитуда тетанического сокращения превышает амплитуду одиночного мышечного сокращения. Н.Е. Введенский объяснил это фазными изменениями возбудимости мышцы, введя понятие об оптимуме и пессимуме частоты раздражения.
Оптимум – частота раздражения, когда каждое последующее раздражение осуществляется в фазу повышенной возбудимости, в результате возникает гладкий тетанус максимальной амплитуды. Общая величина укорочения (и напряжения) в экспериментальных условиях растет тем больше, чем чаще стимуляция. Это продолжается до известных пределов, так как при чрезмерной частоте нервных импульсов развивается частичный блок проведения возбуждения в синапсе, так называемый пессимум (по Н.Е. Введенскому). Пессимум – частота раздражения, когда каждое последую-
75
щее раздражение приходит в фазу пониженной возбудимости и регистрируется тетанус с минимальной амплитудой.
Обычный режим естественного сокращения мышечных волокон близок к зубчатому тетанусу. Однако такой режим функционирования единиц, составляющих ДЕ, не сказывается на сокращении целой мышцы; оно напоминает гладкий тетанус. Причина этого – асинхронность работы мотонейронов и обусловленная этим асинхронность сократительной реакции отдельных мышечных волокон.
Сила мышцы и ее работа
Мышцы способны развивать силу только при укорочении, поэтому чтобы сместить кость и затем вернуть ее в исходное положение, необходимы, по крайней мере, две мышцы или две группы мышц. Пары мышц, действующие таким образом, называются антагонистами.
Сила – это способность преодолевать внешнее сопротивление или противодействовать ему мышечным напряжением. За счет силы производится работа (А):
А = FS,
где F – сила, S – путь, тогда – F = A/S (при равномерном движении).
Механическая работа мышцы (А) по подъему груза Р на высоту h вычисляется по формуле:
А = Ph,
где A – работа; Р – масса груза; h – высота, на которую он поднят.
Поэтому силу мышцы можно определить по величине максимального груза, который она способна поднять. Максимальная работа производится при средних величинах нагрузок. Лабильность мышцы равна 200–300 Гц.
В движениях с ускорением сила (F), сообщающая телу ускорение, равна произведению массы (m) на ускорение (a) (II закон Ньютона):
F = ma.
76
Сила измеряется в ньютонах (Н): 1Н = 1кг/м/с2. Таким образом, существует два проявления силы: в статике – F = P (кг), в динамике – F = ma (Н).
Поскольку каждое волокно подчиняется закону «все или ничего», сила, развиваемая двигательной единицей при одиночном сокращении, варьирует слабо; либо все ее волокна возбуждаются и сокращаются, либо они расслаблены. Развиваемая сила зависит от частоты стимуляции, во время гладкого тетануса она примерно вдвое больше, чем при зубчатом тетанусе. При повышении частоты импульсации мотонейрона от 5 до 50 Гц одиночные сокращения или зубчатый тетанус двигательных единиц переходят в гладкий тетанус, в результате сила сокращения увеличивается. Сила и скорость сокращения мышцы увеличиваются также по мере активации (вовлечения) все большего количества двигательных единиц.
Мощность мышцы равна произведению развиваемой ею силы на скорость укорочения. Например, максимальная мощность (200 Вт) мышцы руки достигается при скорости сокращения 2,5 м/с. Мощность выше при умеренных нагрузках и скоростях сокращения. Максимальная скорость укорочения саркомера определяется максимальной скоростью скольжения актиновых и миозиновых нитей относительно друг друга. Чем быстрее поперечные мостики расщепляют АТФ и взаимодействуют с актином, тем выше скорость этого скольжения. В медленных волокнах, например в позных мышцах, у миозина низкая АТФ-азная активность, и по составу он отличается от миозина с высокой АТФ-азной активностью, который содержится в быстрых волокнах, обеспечивающих движения. Поскольку саркомеры располагаются в миофибриллах последовательно, их укорочения суммируются, так что при одной и той же скорости укорочения саркомера длинная мышца будет сокращаться быстрее, чем короткая.
Скорость сокращения снижается с увеличением нагрузки; если величина нагрузки совпадает с максимальной силой, развиваемой в изометрических условиях, то мышца вообще
77
не укорачивается, а при еще большей нагрузке растягивается, например, при ходьбе под гору. Быстро укорачиваясь, мышца развивает меньшую силу, чем при медленном укорочении. Поэтому очень быстрые движения возможны, только если не требуется большая их сила, т.е. когда мышцы не нагружены,
инаоборот, максимальная мышечная сила требует медленных движений, например, если необходимо передвинуть крупный предмет.
Сила мышц, их способность к работе, скорость сокращения и расслабления, выносливость определяются структурными, функциональными и центральными факторами. Структурные факторы: число и длина мышечных волокон, строение (степень наклона мышечных волокон к оси движения), соотношение типов волокон, из которых она состоит. При преобладании в мышце быстрых волокон – выше скорость сокращения и максимальная сила, развиваемая мышцей при быстром сокращении. Если больше медленных волокон, мышца выносливее, т.е. обладает способностью производить длительную работу. Функциональные факторы обусловлены количественным содержанием в мышцах сократительных белков, АТФ, креатинфосфата, гликогена.
Центральная нервная система определяет внутримышечную координацию (число ДЕ, вовлекаемых в данный момент; степень синхронности работы ДЕ; степень активации каждой ДЕ (определяется частотой нервных импульсов)
имежмышечную координацию (мобилизацию агонистов, торможение антагонистов), а также трофические влияния симпатической нервной системы на мышцы.
Рефлекторный тонус. Благодаря периодической низкочастотной рефлекторной активации небольшого числа двигательных единиц некоторые мышцы (прежде всего мышцы, определяющие позу тела) могут находиться в состоянии длительного непроизвольного напряжения, которое обусловлено асинхронной работой их ДЕ. Такой нейроген-
ный «тонус» модулируется системой γ-волокон мышечных веретен; во время умственного напряжения или эмоционального возбуждения он непроизвольно усиливается, а в состоя-
78