

filo_ontogenez
.pdfне приводить до порушень дихання і ковтання, оскільки ні трахея, ні
стравохід, не є стиснутими.
Подвійна аортальна дуга виникає тоді, коли між місцем відходження сьомої міжсегментної артерії і місцем її з’єднання з лівою спинною аортою зберігається права спинна аорта. Судинне кільце оточує
трахею і стравохід, стискуючи їх, ускладнюючи дихання і ковтання. Права аортальна дуга виникає тоді, коли ліва четверта дуга і
ліва спинна аорта повністю облітеруються і заміщаються відповідними судинами правої сторони. В деяких випадках, коли артеріальна зав'язка розташовується зліва і проходить позаду стравоходу, спо-
стерігається ускладнене ковтання.
Перервана аортальна дуга обумовлюється облітерацією четвертої аортальної дуги зліва. Цей дефект часто об'єднується з аномальним відходженням правої підключичної артерії. Артеріальна протока залишається відкритою і низхідна аорта з підключичними артеріями забезпечуються кров'ю з пониженим вмістом кисню. Аортальний стовбур забезпечує дві загальні сонні артерії.
Аномалії розвитку венозної системи
Складність розвитку порожнистих вен приводить до високої
частоти відхилень від нормальної схеми.
Подвійна нижня порожниста вена розвивається тоді, коли ліва
сакрокардинальна вена не втрачає з’єднання з лівою субкардинальною.
Відсутність нижньої порожнистої вени спостерігається тоді, коли субкардинальна вена не сполучається з печінкою і скидає кров прямо в праву субкардинальну вену; згодом кров з каудальної частини тіла досягає серця через непарну і верхню порожнисту вени. Печінкова вена входить в праве передсердя в тому місці, де впадала б нижня порож-
ниста вена. Звичайно це аномалія пов'язана з іншими дефектами серця. Верхня ліва порожниста вена — це аномалія, викликана збере-
женням лівої передньої кардинальної вени і облітерацією загальної кардинальної і проксимальної частини передньої кардинальної вен справа. У такому випадку кров справа скидається вліво з допомогою плечеголовної вени. Ліва верхня порожниста вена впадає в праве пе-
редсердя крізь лівосинусовий ріг, тобто крізь коронарний синус. Для подвійної верхньої порожнистої вени характерні збере-
ження лівої передньої кардинальної вени і нарушене формування лівої плечеголовної вени. Персистуюча ліва передня кардинальна вена, яка називається лівою верхньою порожнистою веною, впадає в праве передсердя через коронарний синус.
231

2.8. РОЗВИТОК ЕНДОКРИННИХ ЗАЛОЗ
Онтогенетична класифікація ендокринних залоз:
1.Ентодермально-бранхіогенні залози (що походять з глотки
ізябрових кишень зародка) — щитоподібна, паращитоподібні
івилочкова залози.
2.Ентодермально-середньокишкові залози (походять з кишкової трубки) — острівки підшлункової залози.
3.Мезодермально-міжниркові залози (похідні інтерреналової системи) — кіркова речовина наднирників.
4.Мезодермально-мезенхімні залози (похідні мезенхіми) — ендокринні інтерстиціальні елементи статевих залоз (яєчок, яєчників).
5.Ектодермально-неврогенні залози (що походять з проміжного мозку) — епіфіз і гіпофіз.
6.Ектодермально-ротові залози (похідні епітелію кишені Ратке — епітелію ротової бухти) — аденогіпофіз.
Оскільки ендокринні залози мають різне походження, розвиток і будо-
ву і об'єднуються лише за функціональною ознакою (внутрішня секреція), то правильно вважати, що вони складають не систему, а ендокринний апарат.
Морфофункціональна класифікація ендокринних залоз:
1.Центральні (ектодермально-неврогенні): •• гіпофіз; •• епіфіз.
2.Периферичні:
••ектодермально-бранхіогенні — щитоподібна залоза, паращитоподібні залози, тимус;
••адреналові — мозкова речовина наднирників;
••мезодермальні — статеві, кіркова речовина наднирників;
••ентодермальні залози кишкової трубки — острівки Лангерганса.
Розвиток центральних ендокринних залоз
Розвиток гіпоталамо-гіпофізарного комплексу
Філогенез
У круглоротих гіпофіз має примітивну будову. Дно III шлуночка формує нейрогіпофіз, який твердою і м'якою мозковою оболонками відокремлений від аденогіпофіза. Останній розділяється на три частини, які лежать в одній площині.
232

Уриб гіпофіз також складається з нейро- і аденогіпофіза. Позаду аденогіпофіза формується особливий виступ — судинний мішок, який розглядається як гомолог задньої частини гіпофіза інших хребетних. У хрящових риб в гіпофізі є порожнина, у кісткових риб — він суцільний. Починаючи з риб виникає задня частина гіпофіза як самостійний орган.
Уземноводних добре розвинена дистальна частина. Проміжна частина нагадує поперечну смугу.
Урептилій проміжна частина досягає значних розмірів. Дистальна частина вигинається в дорзальному напрямі, проміжна частина товщає і спереду оточує дистальну.
Уптахів дистальна частина округлої або овальної форми ділиться на дві зони — головну (ростральну) і каудальну. Проміжна частина повністю відсутня, а дистальна відокремлена від нейрогіпофізу широкою сполучнотканинною перегородкою.
Уссавців гіпофіз містить всі три частини.
Онтогенез
Гіпоталамічна область мозку розвивається з базальної частини нейроектодермального епітелію, розташованої у вигляді потовщення на вентральній стінці проміжного мозкового міхура. Вона випинається назовні у вигляді лійкоподібного виросту.
Назустріч цьому виросту аж до тісного зіткнення з ним із зачатка ротової порожнини, що примикає до передньої кишки, яка
сліпо закінчується, росте порожнисте пальцеподібне вип’ячування (гіпофизарна кишеня, або кишеня Ратке). Гістологічно ці процеси
виявляються у ембріона людини завдовжки 6,5 мм. На подальших етапах розвитку (у ембріонів завдовжки 12—20 мм) між ними розростається мезенхіма, яка бере участь в структурних перетвореннях кишені Ратке: його гирло починає звужуватися у вигляді трубчастого утворення, яке потім відшнуровується і перетворюється на рудиментарну структуру, що отримала назву глоткового гіпофіза. Тим часом краї проксимальної ділянки кишені Ратке, що сліпо закінчується, змикаються, і вона перетворюється на порожнисту структуру, задня стінка якої тісно прилежить до воронкоподібного виросту діенцефального міхура. Передня стінка кишені Ратке, що замкнулася, сильно розростається і, товщаючи, здавлює порожнину кишені до розмірів вузької щілини. Піддається перетворенням і нервовий зародок: його проксимальна ділянка зберігає форму лійки, що про-
233
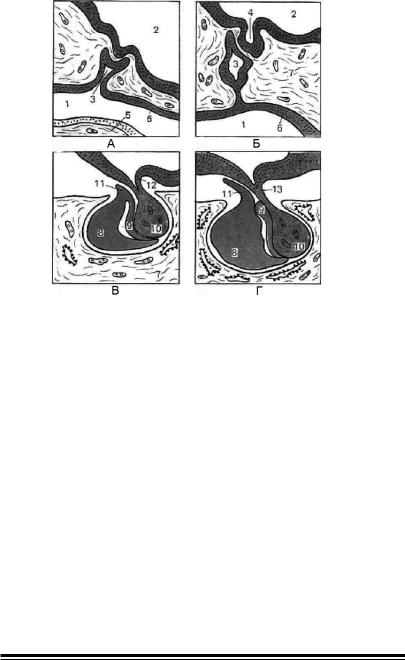
Мал. 16. Стадії розвитку гіпофіза (схема) (за О.В. Волковою, зі змінами,
з Афанасьева Ю. І., 1983).
А — ембріональні зачатки; Б — з’єднання ектодермального і нейрального ембріональних зачатків; В, Г — формування основних частин адено- і нейрогіпофізу; 1 — ротова порожнина; 2 — порожнина шлуночку мозку; 3 — гіпофизарна кишеня Ратке; 4 — дивертикул проміжного мозку; 5 — язик; 6 — ектодермальний епітелій ротової порожнини; 7 —мезенхіма; 8 — передня стінка кишені Ратке (передня частка гіпофіза); 9 — задня стінка кишені Ратке (проміжна частка гіпофіза); 10 — задня частка гіпофіза; 11 — туберальна частина; 12 — епендіма; 13 —гіпофізарна ніжка.
довжується в порожнисте стебло; дистальний кінець стебла втрачає порожнину і починає товщати, оточуючись з усіх боків тонкою задньою стінкою кишені Ратке. Від переднього потовщення стінки кишені Ратке росте язикоподібне вип’ячування, що оточує лійку і проксимальну частину стебла.
Таким чином, в процесі ембріонального розвитку з нервового і епітеліального зачатків формуються дві частки гіпофіза — нерво-
ва, або нейрогіпофіз, і залозиста, або аденогіпофіз. В межах кожної з них виділяють: у нейрогіпофізі — лійкоподібну проксимальну частину — серединне піднесення нейрогіпофіза, стеблевидну — ін-
фундибулярну частину нейрогіпофіза і потовщену дистальну час-
тину, що сліпо закінчується, — головну частину нейрогіпофіза. В аденогіпофізі виділяють: передню головну частину, що розвиваєть-
234

ся з потовщеної передньої стінки кишені Ратке, туберальну части-
ну — її язикоподібний виріст, що оточує серединне піднесення і проксимальну частину стебла; нарешті, проміжну частину, яка розви-
вається з тонкої задньої стінки кишені Ратке і оточує задню головну частину нейрогіпофіза.
В межах гіпоталамо-гіпофізарного комплексу виділяють дві системи, що структурно і функціонально розрізняються: гіпоталамонейрогіпофізарну і гіпоталамо-аденогіпофізарну. Перша повністю розвивається з нервового зачатка і її можна представити як систему крупноклітинних нейронів переднього гіпоталамуса, що походять з його супраоптичного і паравентрикулярного ядер і які закінчуються великою кількістю нервових закінчень у задній головній частині нейрогіпофіза.
Як уже згадувалося, первинною закладкою гіпоталамуса у ембріонів є базальна частина нейроектодермального епітелію, розта-
шована у вигляді потовщення на вентральній стінці проміжного міхура, в якій розрізняють дорсальну, середню і вентральну долі.
Найраніше в ембріогенезі з'являються крупноклітинні ядра гіпоталамуса, що розвиваються з тієї частини зачатка, яка знаходиться на межі середньої і вентральної доль. Пізніше розвиваються супрахіазмальні ядра, які виконують роль центрального координатора механізмів, регулюючих біологічні ритми секреції гормонів. До цього ж періоду відноситься формування інших дрібноклітинних ядер, серед яких найважливішим в регуляції ендокринних функцій, а також репродукції, статевої і харчової поведінки є паравентрикулярне, аркуатне, переднє гіпоталамічне, вентро- і дорсомедіальне ядра. Серед них заслуговує особливої уваги аркуатне ядро, при формуванні якого спостерігається взаємодія з епендімою, що підстилає його. Нейроепітеліальні клітини після формування рострального і каудального компонентів цього витягнутого по горизонтальній осі і зігнутого ядра трансформуються в епендімні клітини, вистилаючі нижньобокові стінки III шлуночка. На рівні рострального компоненту ядра клітини зберігають вид звичайної війчастої епендіми, тоді як на каудальному рівні вони набувають структури, харак-
терної для спеціалізованої епендіми. Ці клітини отримали назву таніцитів від грецьких слів таніс (довгий) і цитос (клітина). Тані-
цити забезпечують двосторонній транспорт біологічно активних речовин (гормонів, нейрогормонів, нейротрансмітерів) з ліквору III шлуночку в портальну кров передньої головної частини аденогіпофіза і навпаки.
235
Розвиток епіфізу
Філогенез
Епіфіз (пінеальна або шишкоподібна залоза) є нейроендокринним органом, присутнім у всіх хребетних тварин. В ході еволюції епіфіз поступово втрачав функцію фоторецептора і перетворювався на нейроендокринну залозу, продукуючи нейрогормони відповідно до інформації про освітленість навколишнього середовища і граючу важливу роль в регуляції процесів росту і розмноження. При цьому спочатку трубчаста структура пінеальних органів замінювалася альвеолярною і паренхіматозно-часточковою.
Укруглоротих шишкоподібне тіло разом з парапінеальним органом формує пінеальній комплекс. У ньому міститься рудиментарний орган зору, який складається з сітківки і кришталика.
Уриб шишкоподібне тіло має залозисту структуру, воно продукує секрет, при цьому зберігає функцію фоторецептора. Тому для епіфізу риб характерна незавершеність трансформації зі світлочутливого органа в ендокринну залозу.
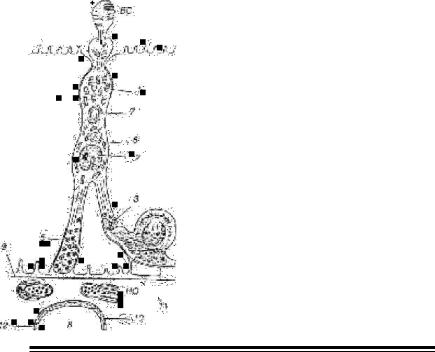
Мал. 17. Пінеалоцити у риб і земноводних (а), плазунів і птахів
(б) (за Коваленко Р. І, Полено-
вим А. Л., 2001).
ВС - зовнішній фоточутливий сегмент; І До – інтерстиціальна клітина; Н - нейрон; АЛЕ - нервове волокно і еготерміналь; 1 - мітохоідрія; 2 - ядро; 3 - синаптичні стрічки; 4 - параболоїд; 5 – секреторні гранули; 6 - гранулярна ендоплазматична мережа; 7 - комплекс Гольджі; 8 - капіляр; 9 - зовнішня базальна пластина; 10 – внутрішня базальна пластина; 11 - перикапілярний простір; 12 – ендотелій фенестрованого типу.
236

Уамфібій епіфіз великий, має трубчасту структуру, добре кровопостачання. Структура відповідає ендокринній залозі з активною секрецією.
Урептилій пінеальна залоза досягає значних розмірів, при цьому зберігає функцію фоторецептора.
Уптахів епіфіз — високоактивний нейроендокринний орган. Епіфіз ссавців — гомолог пінеального органу у Anamnia. Втрачає
фоторецепторну функцію, набувають значно ширших повноважень клітинні елементи.
Онтогенез
Епіфізом більшості ссавців є компактне формування головного мозку, розташоване в борозні між передніми горбами покривної пластинки. Входячи до складу епіталамусу, орган зв'язаний за допомогою двох комісур з габенулярним ядром і субкомісуральним органом, що лежить в задній стінці III шлуночка. У людини епіфіз виникає на 2-му місяці внутріутробного життя з каудальної частини проміжного мозку. У новонароджених вага органу складає 70—90 міліграм, у дорослих людей — 150—200 міліграм. Спочатку епіфіз на сагітальних зрізах має вид трубчастої структури, Причому епіфізарна кишеня сполучається з порожниною III шлуночку. У склепінні епіфізарної кишені поступово формується жолобок, в якому можна бачити судини, що сполучаються з венозними синусами мозкової оболонки. Інтенсивна облітерація епіфізарної кишені, що розповсюджується від дистальних відділів органу до проксимальних, приводить до формування численних складок. Одночасно відбувається проліферація не тільки клітин епітелію, але і «вільних» клітин, наслідком чого є потовщення середніх і, особливо, дистальних відділів органу. На пізніших термінах ембріонального розвитку епіфіз має компактну структуру. Поверхня епіфізу злегка горбиста, колір — сірувато-рожевий. Він оточений сполучнотканинною оболонкою, від якої всередину відходять прошарки, що розділяють паренхіму органу на окремі нечітко виражені часточки. У проксимальній частині залози поблизу від пінеальної бухти III шлуночку розташовуються альвеолярні структури (залишки ембріональної кишені).
Кровопостачання органу здійснюється судинною мережею, утвореною вторинними гілками середньої і задньої мозкових артерій. Увійшовши до сполучнотканинної капсули органу, судини розпадаються на безліч капілярів з формуванням мережі, що характеризується великою кількістю анастомозів. Кров від епіфізу від-
237
водиться частково в систему великої мозкової вени Галена, а деяка кількість її поступає у вени судинного сплетення III шлуночку. Кров в епіфізі протікає з великою швидкістю, поступаючись тільки швидкості кровотоку в нирці. Особливістю будови капілярів епіфізу ссавців є те, що у представників одних отрядів (гризуни) в ендотеліоцитах є фенестри, а у інших (хижі, примати) такі формування відсутні. Ці структурні відмінності співпадають з функціональними властивостями капілярів, які виражаються в першому випадку у відсутності гематоенцефалічного бар'єру, а в другому — в його наявності. Фенестрація ендотеліоцитів капілярів, на думку більшості авторів, забезпечує проникнення високомолекулярних речовин в порожнину судини.
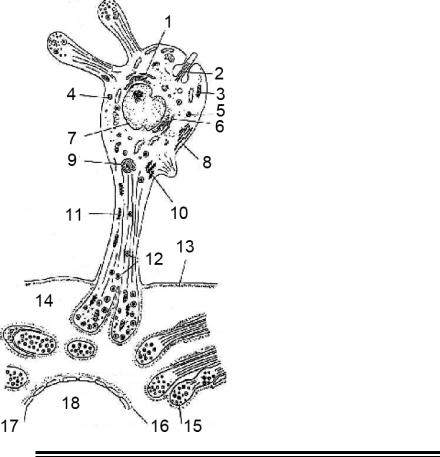
Мал. 18. Схема будови пінеалоцита у ссавців (за Коваленко Р. І., Поленовим А.
Л., 2001).
1-гранулярнаендоплазматична мережа; 2 - вія; 3 - мітохондрія; 4 - мультивезикулярне тільце; 5 - лизосома; 6 - комплекс Гольджі; 7 - ядро; 8 - агранулярна ендоплазматична мережа; 9 - мультиламеллярне тільце; 10 - синаптичні стрічки; 11 - синаптичні стрічки із спустошеними везикулами; 12 - секреторні гранули; 13 - зовнішня базальна пластина; 14 - перикапиллярний простір; 15 - нервове волокно і його терміналь; 16 - внутрішня базальна пластина; 17 - фенестрирований ендотеліоцит; 18 - капіляр.
238

Основними клітинними елементами пінеальних органів y Anamnia є клітини фоторецепторів (сприймають світловий сигнал і трансформують його в електричний стимул), нейрони (сприймають електричний стимул і передають його в інші відділи головного мозку) і опорні, або інтерстиціальні клітини (виконують, можливо, також нейроендокринну функцію). У пінеальних органах плазунів і в альвеолярних структурах епіфізу птахів виявлені особливі рудиментарні клітини фоторецепторів, в тілі і відростках яких виявляються секреторні гранули. Останні зустрічаються і в деяких інтерстиціальних клітинах.
Переважаючими клітинами епіфізу ссавців є пінеалоцити (ПЦ). Довгі відростки ПЦ, що гілкуються, як правило, закінчуються на базальній пластині перикапілярного простору. Розрізняють більші «світлі» ПЦ з гомогенною цитоплазмою і меншого розміру «темні» ПЦ з ацидофільними або базофільними гранулами у цитоплазмі. Невідомо, чи є ці два різновиди ПЦ самостійними клітинними типами, або є функціональними і віковими різновидами клітин епіфізу. Цікаво, що у деяких видів кажанів, що ведуть виключно нічний спосіб життя, виявляється одна єдина популяція ПЦ світлого типу. З використанням стереометричного аналізу виявлено, що світлі ПЦ мають округліші ядра і містять меншу кількість гетерохроматину. У темних ПЦ виявляються мітохондрії з численними кристами, цистерни ГЕС коротші і численніші, КГ має велику щільність. Деякі автори наводять дані про те, що у молодих тварин відносний вміст світлих ПЦ вищий, ніж у дорослих, а інші повідомляють протилежні дані. Л. А. Колеснікова (1996) виділяє в епіфізі хижих ссавців не два, а чо-
тири різні типи ПЦ: світлі, темні, проміжні і гранулярні.
Вікові особливості. Деяка кількість пінеалоцитів зазнає атрофії,
а строма розростається і в ній збільшується відкладення фосфатних і карбонатних солей у вигляді накопичень, названих мозковим піском.
Розвиток периферичних ендокринних залоз
Розвиток щитоподібної залози
Філогенез
Щитоподібна залоза є у всіх класів хребетних, хоча у костистих риб її паренхіма не об'єднана в компактний орган, а є залозистою тканиною, розкиданою уздовж аорти. У земноводних і птахів залоза складається з двох незв'язаних доль, що лежать на значній відстані одна від одної. У рептилій і ссавців долі зближуються, розташовуються по обох сторонах трахеї і, як правило, зв'язані перешийком.
239

Онтогенез
У людини перші ознаки формування щитоподібної залози спостерігаються у 22—26-добового ембріона (довжина тіла 2—4 мм), коли відбувається проліферація і випинання клітинного матеріалу дна
глотки між I і II парами глоткових кишень в області майбутнього сліпого отвору язика. Цей виріст, що іменується медіальним зачатком,
майже із самого початку має дводольчату будову і зміщується у каудальному напрямі, втрачаючи зв'язок з ембріональною глоткою. У окремих випадках закладка щитоподібної залози не формується (аплазія органу).
Епітелій, що походить з медіального зачатка і формуючий основну масу щитоподібної залози дорослого організму, формально розвивається з головної кишки, проте він є дериватом прехордальної пластинки, у зв'язку з чим має епідермальну (по Н. Г. Хлопіну 1946) детермінацію. У епітеліальну закладку органу вростають сполучна тканина і судини. Об'єм часток, що складаються з анастомозуючих тяжів пролифера епітеліальних клітин, прогресивно збіль-
шується. Ця стадія розвитку щитоподібної залози називається передфолікулярною і відповідає 6—9-у тижню внутріутробного
розвитку. Надалі (на 10—11-му тижні) у клітинах починають накопичуватися ШИК-позитивні краплі, які зливаються у екстрацелюлярному просторі з формуванням перших мікрофолікулів (11—12-ий тижні). Практично в цей же період з'являється характерна для щитоподібної залози здатність до накопичення йоду і (за деякими даними, дещо пізніше) до синтезу тиреоїдних гормонів, а також чутливість до дії стимулюючих чинників. Таким чином, гістогенез щитоподібної залози тісно поєднується з її функціональним дозріванням. Морфогенез фолікулів пов'язаний з поляризацією тироцитів і формуванням між ними комплексу контактів, які обумовлюють формування просвіту фолікула. В ході ембріонального розвитку фолікули безперервно збільшуються у розмірах, одночасно відбувається їх новоформування. Частина епітеліальних клітин — похідних медіального зачатка — залишається у вигляді компактних інтерфолікулярних скупчень (острівців), вміст яких у міру розвитку плоду прогресивно знижується.
Як показують морфометричні дослідження, інтенсивний фолікулогенез і накопичення колоїду характерні для періоду 10—18 тижнів; з 19-го по 29-ий тижні відбувається деяка стабілізація розмірів фолікулів і співвідношення об'ємів епітелію і колоїду в залозі. Після
240