

Промежуточный обмен липидов
Синтез и распад липидов происходит в течение 1-2 суток. Адипоциты жировых тканей играют основную роль в обмене триацилглицеринов, которые в них синтезируются, откладываются про запас и по мере необходимости расходуются. Липолиз – расщепление жира в жировой ткани осуществляется ферментами моно-, ди-, три-глицеридлипазами, которые расщепляют триацилглицериды на глицерин и жирные кислоты. Жирные кислоты и глицерин в клетках организма окисляются до углекислоты и воды, а энергия, образующаяся при этом аккумулируется в молекулах АТФ.
Окисление глицерина начинается с фосфорилирования с участием АТФ. В результате этой реакции образуется фосфоглицериновая кислота, которая при участии НАД подвергается дегидрированию с образованием 3-фосфоглицеринового альдегида:
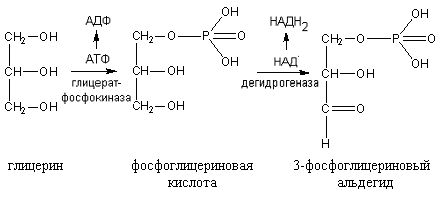
3-фосфоглицериновый альдегид далее окисляется как при гликолизе. Часть фосфоглицериновой кислоты используется для синтеза фосфатидов.
Окисление жирных кислот происходит по β-углеродному атому в матриксе митохондрий. Жирные кислоты с короткой углеродной цепью (4-10 атомов) проникают в митохондрии; а с более длинной цепью проникают в виде ацилкарнитинов:
ж ирная
кислота + АТФ + карнитин ацилкарнитин
ирная
кислота + АТФ + карнитин ацилкарнитин
При β-окислении жирных кислот в каждом цикле отделяется по 2 С в виде активированной уксусной кислоты (ацетил-КоА) – CH3-CO-SKoA. Например, при окислении пальмитиновой кислоты (C16) образуется 8 молекул CH3-CO-SKoA:
![]()
Последовательность реакции окисления жирной кислоты выглядит следующим образом: в начале жирная кислота активируется с участием HS-коэнзима А, в результате образуется ацил-КоА. В митохондриях ацил-КоА подвергается дегидрированию с участием фермента ацил-КоА-дегидрогеназы, при этом происходит восстановление ФАД в ФАДН2, образуется ненасыщенная активированная жирная кислота:
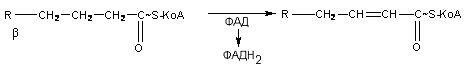
На следующем этапе происходит присоединение молекулы воды с участием фермента еноилгидратазы, в результате образуется β-гидроксиацил-КоА:
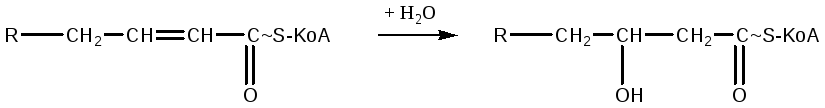
Далее происходит образование кетокислоты в результате дегидрирования β-гидроксиацил-КоА: реакция происходит с участием кофермента НАД, который восстанавливается в НАДН2:
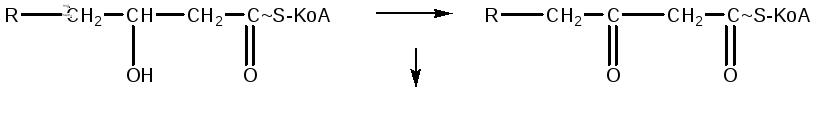
Молекула кетоформы ацил-КоА под влиянием фермента и при наличии одной молекулы HS-KoA разрывается на 2 части: ацетил-КоА и ацил-КоА, который имеет на два атома углерода меньше исходной кислоты:
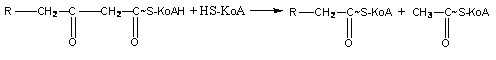 ацил-КоА
ацетил-Коа
ацил-КоА
ацетил-Коа
Ацетил-КоА подвергается окислению в цикле Кребса, ацил-KoA - снова проходит путь β-окисления, при этом каждый раз цепь укорачивается на 2 атома углерода.
Окисление жирной кислоты, имеющий 2n атомов углерода происходит за n-1 циклов, так как, окисление бутирил-КоА (4C) завершается за один цикл.
Энергетический баланс β-окисления жирных кислот
При каждом цикле образуется ФАДН2, НАДН2 которые в дыхательной цепи дают:
ФАДН2 – 2 молекулы АТФ
НАДН2 – 3 молекулы АТФ.
Окисление 1 молекулы CH3-CO-KoA в цикле Кребса дает 12 молекул АТФ.
При полном окислении жирной кислоты (n/2)-1 циклов β-окисления, возникает n/2 молекул ацетил-КоА, следовательно, при окислении, например, пальмитиновой кислоты (C16) образуется 130 молекула АТФ.