

Вопрос 51



Спинной мозг, его строение и функции.
Вопрос 52
Спинной мозг – наиболее древнее образование ЦНС. Характерная особенность строения – сегментарность.Нейроны спинного мозга образуют его серое вещество в виде передних и задних рогов. Они выполняют рефлекторную функцию спинного мозга.Задние рога содержат нейроны (интернейроны), которые передают импульсы в вышележащие центры, в симметричные структуры противоположной стороны, к передним рогам спинного мозга. Задние рога содержат афферентные нейроны, которые реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения.Передние рога содержат нейроны (мотонейроны), дающие аксоны к мышцам, они являются эфферентными. Все нисходящие пути ЦНС двигательных реакций заканчиваются в передних рогах.В боковых рогах шейных и двух поясничных сегментов располагаются нейроны симпатического отдела вегетативной нервной системы, во втором—четвертом сегментах – парасимпатического.В составе спинного мозга имеется множество вставочных нейронов, которые обеспечивают связь с сегментами и с вышележащими отделами ЦНС, на их долю приходится 97 % от общего числа нейронов спинного мозга. В их состав входят ассоциативные нейроны – нейроны собственного аппарата спинного мозга, они устанавливают связи внутри и между сегментами.Белое вещество спинного мозга образовано миелиновыми волокнами (короткими и длинными) и выполняет проводниковую роль.Короткие волокна связывают нейроны одного или разных сегментов спинного мозга.Длинные волокна (проекционные) образуют проводящие пути спинного мозга. Они формируют восходящие пути, идущие к головному мозгу, и нисходящие пути, идущие от головного мозга.Спинной мозг выполняет рефлекторную и проводниковую функции.Рефлекторная функция позволяет реализовать все двигательные рефлексы тела, рефлексы внутренних органов, терморегуляции и т. д. Рефлекторные реакции зависят от места, силы раздражителя, площади рефлексогенной зоны, скорости проведения импульса по волокнам, от влияния головного мозга.
Рефлексы делятся на:1) экстероцептивные (возникают при раздражении агентами внешней среды сенсорных раздражителей);2) интероцептивные (возникают при раздражении прессо-, механо-, хемо-, терморецепторов): висцеро-висцеральные – рефлексы с одного внутреннего органа на другой, висцеро-мышечные – рефлексы с внутренних органов на скелетную мускулатуру;3) проприоцептивные (собственные) рефлексы с самой мышцы и связанных с ней образований. Они имеют моносинаптическую рефлекторную дугу. Проприоцептивные рефлексы регулируют двигательную активность за счет сухожильных и позотонических рефлексов. Сухожильные рефлексы (коленный, ахиллов, с трехглавой мышцы плеча и т. д.) возникают при растяжении мышц и вызывают расслабление или сокращение мышцы, возникают при каждом мышечном движении;4) позотонические рефлексы (возникают при возбуждении вестибулярных рецепторов при изменении скорости движения и положения головы по отношению к туловищу, что приводит к перераспределению тонуса мышц (повышению тонуса разгибателей и уменьшению сгибателей) и обеспечивает равновесие тела)Исследование проприоцептивных рефлексов производится для определения возбудимости и степени поражения ЦНС.Проводниковая функция обеспечивает связь нейронов спинного мозга друг с другом или с вышележащими отделами ЦНС.
Нейроны спинного мозга, их функции.
Нейроны спинного мозга образуют его серое вещество в виде симметрично расположенных двух передних и двух задних рогов в шейном, поясничном и крестцовом отделах. Серое вещество распределено на ядра, вытянутые по длине спинного мозга, и на поперечном разрезе располагается в форме буквы Н. В грудном отделе спинной мозг имеет, помимо названных, еще и боковые рога (рис. 4.9).
Задние рога выполняют главным образом сенсорные функции и содержат нейроны, передающие сигналы в вышележащие центры, в симметричные структуры противоположной стороны либо к передним рогам спинного мозга.
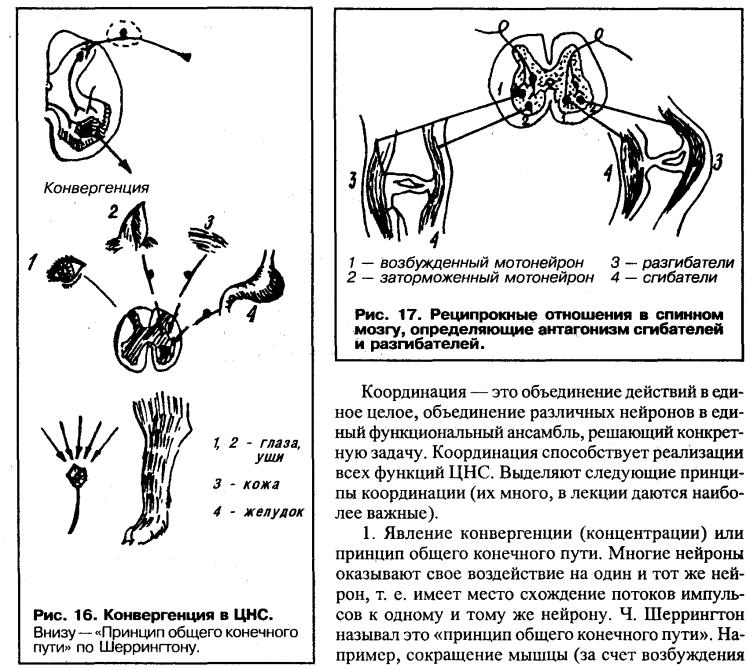
В передних рогах находятся нейроны, дающие свои аксоны к мышцам. Все нисходящие пути центральной нервной системы, вызывающие двигательные реакции, заканчиваются на нейронах передних рогов. В связи с этим Шеррингтон назвал их «общим конечным путем».
Начиная с I грудного сегмента спинного мозга и до первых поясничных сегментов, в боковых рогах серого вещества располагаются нейроны симпатического, а в крестцовых — парасимпатического отдела автономной (вегетативной) нервной системы.
Спинной мозг человека содержит около 13 млн. нейронов, из них 3% — мотонейроны, а 97% — вставочные. Функционально нейроны спинного мозга можно разделить на 4 основные группы:
1) мотонейроны, или двигательные, — клетки передних рогов, аксоны которых образуют передние корешки;
2) интернейроны — нейроны, получающие информацию от спинальных ганглиев и располагающиеся в задних рогах. Эти нейроны реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения;
3) симпатические, парасимпатические нейроны расположены преимущественно в боковых рогах. Аксоны этих нейронов выходят из спинного мозга в составе передних корешков;
4) ассоциативные клетки — нейроны собственного аппарата спинного мозга, устанавливающие связи внутри и между сегментами.
В средней зоне серого вещества (между задним и передним рогами) спинного мозга имеется промежуточное ядро (ядро Кахаля) с клетками, аксоны которых идут вверх или вниз на 1—2 сегмента и дают коллатерали на нейроны ипси- и контралатеральной стороны, образуя сеть. Подобная сеть имеется и на верхушке заднего рога спинного мозга — эта сеть образует так называемое студенистое вещество (желатинозная субстанция Роланда) и выполняет функции ретикулярной формации спинного мозга.
Средняя часть серого вещества спинного мозга содержит преимущественно короткоаксонные веретенообразные клетки (промежуточные нейроны), выполняющие связующую функцию между симметричными отделами сегмента, между клетками его передних и задних рогов.
Мотонейроны. Аксон мотонейрона своими терминалами иннервирует сотни мышечных волокон, образуя мотонейронную единицу. Чем меньше мышечных волокон иннервирует один аксон (т. е. чем меньше количественно мотонейронная единица), тем более дифференцированные, точные движения выполняет мышца (см. раздел 2.4).
Несколько мотонейронов могут иннервировать одну мышцу, в этом случае они образуют так называемый мотонейронный пул. Возбудимость мотонейронов одного пула различна, поэтому при разной интенсивности раздражения в сокращение вовлекается разное количество волокон одной мышцы. При оптимальной силе раздражения сокращаются все волокна данной мышцы; в этом случае развивается максимальное сокращение мышцы.
Мотонейроны спинного мозга функционально делят на α- и γ-нейроны.
α-Мотонейроны образуют прямые связи с чувствительными путями, идущими от экстрафузальных волокон мышечного веретена, имеют до 20 000 синапсов на своих дендритах и характеризуются низкой частотой импульсации (10—20 в секунду), γ-Мотонейроны, иннервирующие интрафузальные мышечные волокна мышечного веретена, получают информацию о его состоянии через промежуточные нейроны. Сокращение интрафузального мышечного волокна не приводит к сокращению мышцы, но повышает частоту разрядов импульсов, идущих от рецепторов волокна в спинной мозг. Эти нейроны обладают высокой частотой импульсации (до 200 в секунду).
Интернейроны. Эти промежуточные нейроны, генерирующие импульсы с частотй до 1000 в секунду, являются фоновоактивными и имеют на своих дендритах до 500 синапсов. Функция интернейронов заключается в организации связей между структурами спинного мозга и обеспечении влияния восходящих и нисходящих путей на клетки отдельных сегментов спинного мозга. Очень важной функцией интернейронов является торможение активности нейронов, что обеспечивает сохранение направленности пути возбуждения. Возбуждение интернейронов, связанных с моторными клетками, оказывает тормозящее влияние на мышцы-антагонисты.
Нейроны симпатического отдела автономной системы. Расположены в боковых рогах сегментов грудного отдела спинного мозга. Эти нейроны являются фоновоактивными, но имеют редкую частоту импульсации (3—5 в секунду).
Нейроны парасимпатического отдела автономной системы. Локализуются в сакральном отделе спинного мозга и являются фоновоактивными.
В случаях раздражения и поражения задних корешков спинного мозга наблюдаются «стреляющие», опоясывающие боли на уровне метамера пораженного сегмента, снижение чувствительности всех видов, утрата или снижение рефлексов, вызываемых с метамера тела, который передает информацию в пораженный корешок.
В случаях изолированного поражения заднего рога утрачивается болевая и температурная чувствительность на стороне повреждения, а тактильная и проприоцептивная сохраняется, так как из заднего корешка аксоны температурной и болевой чувствительности идут в задний рог, а аксоны тактильной и проприоцептивной — прямо в задний столб и по проводящим путям поднимаются вверх.
Вследствие того, что аксоны вторых нейронов болевой и температурной чувствительности идут на противоположную сторону через переднюю серую спайку спинного мозга, при повреждении этой спайки на теле симметрично утрачивается болевая и температурная чувствительность.
Поражение переднего рога и переднего корешка спинного мозга приводит к параличу мышц, которые теряют тонус, атрофируются, при этом исчезают рефлексы, связанные с пораженным сегментом.
В случае поражения боковых рогов спинного мозга исчезают кожные сосудистые рефлексы, нарушается потоотделение, наблюдаются трофические изменения кожи, ногтей. При одностороннем поражении парасимпатического отдела автономной нервной системы на уровне крестцовых отделов спинного мозга нарушений дефекации и мочеиспускания не наблюдается, так как корковая иннервация этих центров является двусторонней.
Проводящие пути спинного мозга, их функции.
Восходящие проводящие пути. Несут импульсы от рецепторов, воспринимающих информацию из внешнего мира и внутренней среды организма. В зависимости от вида чувствительности, которую они проводят, их делят на пути экстеро—, проприо— и интероцептивной чувствительности. Нисходящие пути передают импульсы от структур головного мозга к двигательным ядрам, осуществляющим ответные реакции на внешние и внутренние раздражения.
Основными восходящими путями спинного мозга являются тонкий пучок, клиновидный пучок, латеральный и вентральный спинно—таламические пути, дорсальный и вентральный спинно—мозжечковые пути (рис. 3.19).
Тонкий пучок (Голля) и клиновидный пучок (Бурдаха) составляют задние канатики спинного мозга. Эти пучки волокон являются отростками чувствительных клеток спинальных ганглиев, проводящих возбуждение от проприорецепторов мышц, сухожилий, частично тактильных рецепторов кожи, висцерорецепторов. Волокна тонкого и клиновидного пучков — миелинизированные, они проводят возбуждение со скоростью 60—100 м/с. Короткие аксоны обоих пучков устанавливают синаптические связи с мотонейронами и интернейронами своего сегмента, длинные же направляются в продолговатый мозг. По пути они отдают большое число ветвей к нейронам вышележащих сегментов спинного мозга, образуя, таким образом, межсегментарные связи.
По волокнам тонкого пучка проводится возбуждение от каудальной части тела и тазовых конечностей, по волокнам клиновидного пучка — от краниальной части тела и грудных конечностей. В спинном мозгу оба этих пути идут, не прерываясь и не перекрещиваясь, и оканчиваются в продолговатом мозгу у одноименных ядер, где образуют синаптическое переключение на второй нейрон. Отростки второго нейрона направляются к специфическим ядрам таламуса противоположной стороны, образуя тем самым своеобразный перекрест. Здесь они переключаются уже на третий нейрон, аксоны которого достигают нейронов IV слоя коры больших полушарий.
Считают, что по этой системе проводится информация тонко дифференцированной чувствительности, позволяющая определить локализацию, контур периферического раздражения, а также его изменения во времени.
По латеральному спинно—таламическому пути проводится болевая и температурная чувствительность, по вентральному спинно—таламическому —тактильная. Существуют сведения, что по этим путям возможна также передача возбуждения от проприо— и висцероцепторов. Скорость проведения возбуждения в волокнах составляет 1—30 м/с. Спинно—таламические пути прерываются и перекрещиваются либо на уровне сегмента, в который они только что вступили, либо вначале проходят несколько сегментов по своей стороне, а затем переходят на противоположную. Отсюда идут волокна, оканчивающиеся в таламусе. Там они образуют синапсы на нервных клетках, аксоны которых направляются в кору больших полушарий.
Полагают, что по системе волокон этих путей в основном передается информация о качественной природе раздражителей.
Дорсальный спинно—мозжечковый путь, или пучок Флексига — филогенетически это наиболее древний чувствительный путь спинного мозга. Местом расположения нервных клеток, аксоны которых образуют волокна этого пути, является основание дорсального рога спинного мозга. Не перекрещиваясь, путь достигает мозжечка, где каждое волокно занимает определенную область. Скорость проведения по волокнам спинно—мозжечкового пути около 110 м/с. По ним проводится информация от рецепторов мышц и связок конечностей. Наибольшего развития этот путь достигает у копытных животных.
Вентральный спинно—мозжечковый путь, или пучок Говерса, также образуется аксонами интернейронов противоположной стороны спинного мозга. Через продолговатый мозг и ножки мозжечка волокна направляются к коре мозжечка, где занимают обширные площади. Импульсы со скоростью проведения до 120 м/с идут от сухожильных, кожных и висцерорецепторов. Они участвуют в поддержании тонуса мышц для выполнения движений и сохранения позы.
Нисходящие проводящие пути. Эти пути (рис. 3.20) связывают высшие отделы ЦНС с эффекторными нейронами спинного мозга. Основными из них являются пирамидный, красноядерно—спинномозговой и ретикулярно—спинномозговой пути.
Пирамидный путь образован аксонами клеток двигательной зоны коры больших полушарий. Направляясь к продолговатому мозгу, эти аксоны отдают большое число коллатералей структурам промежуточного, среднего, продолговатого мозга и ретикулярной формации. В нижней части продолговатого мозга большая часть волокон пирамидного пути переходит на противоположную сторону (перекрест пирамид), образуя латеральный пирамидный путь. В спинном
мозгу он располагается в боковом канатике. Другая часть волокон идет, не перекрещиваясь, до спинного мозга и только на уровне сегмента, в котором оканчивается, переходит на противоположную сторону. Это прямой вентральный пирамидный путь. Оба заканчиваются на мотонейронах передних рогов серого вещества спинного мозга. Состав волокон этого пути неоднороден, в нем представлены миелинизированные и немиелинизированные волокна разного диаметра со скоростями проведения возбуждения от 1 до 100 м/с.
Основной функцией пирамидных путей является передача импульсов для выполнения произвольных движений. Надежность в осуществлении этой функции повышается благодаря дублированию связи головного мозга со спинным посредством двух путей — перекрещенного и прямого. В эволюционном ряду пирамидный тракт развивался параллельно с развитием коры больших полушарий и достиг наибольшего совершенства у человека.
Красноядерно—спинномозговой путь (Монакова) образован аксонами клеток красного ядра среднего мозга. Выйдя из ядра, волокна полностью переходят на противоположную сторону. Часть из них направляется в мозжечок и ретикулярную формацию, другие — в спинной мозг. В спинном мозгу волокна располагаются в боковых столбах перед перекрещенным пирамидным путем и оканчиваются на интернейронах соответствующих сегментов. Красноядерно—спинномозговой путь несет импульсы от мозжечка, ядра вестибулярного нерва, полосатого тела.
Основное назначение красноядерно—спинномозгового пути — управление тонусом мышц и непроизвольной координацией движений. В процессе эволюции этот путь возник рано. Большое значение он имеет у животных, слабее развит у человека.
Преддверно—спинномозговой путь образован волокнами, которые являются отростками клеток латерального пред дверного ядра (ядра Дейтерса), лежащего в продолговатом мозгу. Этот тракт имеет наиболее древнее эволюционное происхождение. По нему передаются импульсы от вестибулярного аппарата и мозжечка к мотонейронам вентральных рогов спинного мозга, регулирующие тонус мускулатуры, согласованность движений, равновесие. При нарушении целостности этого пути наблюдаются расстройства координации движений и ориентации в пространстве.
В спинном мозгу помимо основных длинных имеются и короткие нисходящие пути, соединяющие между собой его отдельные сегменты.
Рефлексы спинного мозга, методы их исследования.
Роль спинного мозга в организации и регуляции движений.