

ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ
ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ «КУРСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ» МИНИСТЕРСТВА ЗДРАВООХРАНЕИЯ И СОЦИАЛЬНОГО РАЗВИТИЯ
РОССИЙСКОЙ ФЕДЕРАЦИИ (ГБОУ ВПО КГМУ МИНЗДРАВСОЦРАЗВИТИЯ РОССИИ)
КАФЕДРА
, НОРМАЛЬНОЙ ФИЗИОЛОГИИ
КОНТРОЛЬНЫЕ ВОПРОСЫ К ЭКЗАМЕНУ
по дисциплине нейрофизиология Факультет: клинической психологии Курс: I Форма обучения: очная
Цели и задачи нейрофизиологии.
Нейрофизиология — раздел физиологии животных и человека, изучающий функции нервной системы и её основных структурных единиц — нейронов. Она тесно связана с нейробиологией, психологией, неврологией, клинической нейрофизиологией, электрофизиологией, этологией, нейроанатомией и другими науками, занимающимися изучением мозга.
Мозг представляет наиболее сложно организованную материю, каждый элемент которой постоянно взаимодействует с информацией, поступающей как из окружающей среды, так и из внутренней среды организма. Анализ этой полимодальной информации необходим в адаптации организма к постоянно меняющимся условиям обитания, его выживаемости и поддержании гомеостаза. Физиология изучает законы, управляющие этими процессами. Нервная система, наряду с иммунной и эндокринной системами, интегрирует различные ткани и органы в единое целое, поэтому знание механизмов этого влияния являются первостепенно важными для понимания работы мозга.
В курсе рассматривается вклад нейробиологии в понимание психической деятельности, физиологические закономерности эмбрионального и постнатального развития. Специальные разделы курса посвящены рассмотрению фундаментальных процессов возбуждения и торможения в ЦНС, физиологии и нейрохимии нейронов и ганглий, функционированию соматической и вегетативной нервных систем.
Задача курса – сформировать у студентов представление о связи психических явлений с особенностями функционирования центральной нервной системы, об интеграции вегетативных, нейроэндокринных и центральных регуляций при осуществлении поведения на базе основных биологических мотиваций, рассмотреть нервные структуры, нейро-гормональные механизмы в регуляции питьевого, пищевого, полового поведения. В результате изучения курса студент должен иметь представление:
об основных направлениях и методах изучения функций нейрона и центральной нервной системы;
о механизмах кодирования информации в нервной системе;
об общих и специфических особенностях функционирования центральной и периферической (вегетативной) нервной системы.
Должен знать:
основные методы исследования в физиологии человека
вклад отечественных и зарубежных ученых в развитие взглядов на физиологию ЦНС;
механизмы нейро-гормональной регуляции функций органов и систем организма;
физиологические закономерности функций пищеварения, дыхания, кровообращения и репродукции;
Понятие о физиологической функции.
Физиологическая функция — это проявление жизнедеятельности, имеющее приспособительное значение.
под фуекцией понимают специфическую деятельность системы или органа.Например,функция дыхания обмен кислорода и углекислого газа Организм может существовать только в том случае, если он отвечает приспособительными реакциями на изменения, происходящие в окружающей среде. Существование организма было бы, например, невозможно, если бы он не реагировал различным образом на пищевые вещества и на вредоносные агенты, если бы недостаток кислорода или воды в окружающей среде не вызывал ответных приспособительных реакций со стороны организма, и т. д. Лишь благодаря разнообразным реакциям на изменения, происходящие в окружающей среде и в нем самом, «как часть природы каждый животный организм представляет собой сложную обособленную систему, внутренние силы которой каждый момент, покуда она существует как таковая, уравновешиваются с внешними силами окружающей среды... Таким образом вся жизнь от простейших до сложнейших организмов... есть длинный ряд все усложняющихся до высочайшей степени уравновешиваний внешней среды» •> Эти бесчисленные уравновешивания организма выявляются в его разнообразных реакциях в ответ на действие агентов окружающего мира. Физиологические процессы, обеспечивающие приспособление организма к окружающей его среде, относятся к явлениям регуляции. Явления .регуляции основаны на связи всех органов животного организма между собой. Лишь при наличии такой связи в организме различные внешние и внутренние агенты могут вызвать согласованные ответные реакции в ряде его органов (сокращение мышц, изменение кровообращения и дыхания и т. д.). Химическая связь, осуществляемая благодаря переносу от одних органов к другим различных химических веществ жидкостями тела (т.. е. гуморально), не может приводить к быстро сменяющимся и диференцированным реакциям, соответствующим огромному разнообразию действующих агентов. Поэтому у животных развивается и приобретает основное значение в регуляции функций организма нервная система, связанная с особыми образованиями, воспринимающими раздражения из внешней и внутренней среды, — рецепторами. Нервная регуляция настолько важна для понимания всех процессов в организме, что с основными данными, ее характеризующими, необходимо ознакомиться в начале изучения физиологии. Процесс нервного возбуждения Нервная система построена из огромного количества разнообразных нервных клеток, обычно имеющих один более длинный отросток — аксон и несколько обычно коротких отростков — дендритов. Нервная клетка со всеми ее отростками называется невроном. Аксоны нервных клеток, выходящие из центральной нервной системы (или вступающие в нее от рецепторов), представляют собой нервное волокно, обычно покрытое шванновской и миэлиновой оболочками. Эти волокна собираются в более или менее толстые стволы, называемые нервами. Каждый нерв содержит огромную массу разнообразных нервных волокон, каждое из которых является отростком какой-либо одной нервной клетки. В нервной ткани под влиянием раздражения возникает возбуждение, которое характеризуется тем, что оно, во-первых, очень быстро распространяется по всему неврону и, во-вторых, воздействует на клетки, к которым подходит аксон возбужденного неврона. Приведем несколько примеров. Классический объект для исследования нервного проведения — так называемый нервно-мышечный препарат лягушки, т. е. извлеченная из тела мышца лягушки с подходящим к ней нервом. Если мы будем в какой-либо точке раздражать нерв, нанося механическое, тепловое или электрическое раздражение, то произойдет сокращение мышцы, в которую входят волокна раздражаемого нерва. То же самое легко показать на человеке: если через кожу раздражать ударами индукционного тока или толчками постоянного электрического тока нервные стволы, идущие близ поверхности тела, то наступает сокращение мышц, иннерви-руемых этими нервами. Рассмотрим еще один пример. Мы производим, например, у собаки или у кошки электрическое раздражение проходящего на шее блуждающего нерва (среди волокон этого нерва есть такие, которые заканчиваются в сердце, а есть и такие, которые заканчиваются в желудке и стенке кишок). Оказывается, что при этом сокращения сердца замедляются или прекращаются, построенная же из гладкой мускулатуры стенка кишечника усиливает свои медленные, червеобразные движения, желудок начинает отделять желудочный сок. При распространении возбуждения по нервному волокну каждая его точка, одна за другой, начиная от места, где нанесено раздражение, приходит в состояние возбуждения. Процесс возбуждения одного участка нервного волокна передается соседнему участку. Ответом любого органа на возбуждение иннервирующих, т. е. оканчивающихся в его тканях, нервных волокон может быть или переход от состояния покоя к деятельности, или усиление уже имеющейся деятельности, а в ряде случаев, как это видно на сердце при раздражении блуждающего нерва, и ослабление этой деятельности, т. е. торможение. Общим, однако, всегда является то, что деятельность или состояние любого органа изменяется, если и н-нервирующие его нервные волокна приходят в состояние возбуждения. Нервное возбуждение является, следовательно, процессом, который меняет физиологическое состояние тех образований, в которых заканчиваются возбудившиеся нервные волокна. Такими образованиями являются либо ткани различных органов тела (например мышечная, эпителиальная, соединительная), либо другие нервные клетки. В последнем случае возбуждение, возникшее в одном невроне, вызывает возбуждение или иное изменение в другом невроне. Нервное возбуждение является, как говорил II. П. Павлов, «раздражительным процессом».
Уровни функционирования организма.
-организменный(функции целостного организма(поведение человека))
-системный(функции отдельных систем(пищеварения,выделения))
-органный(ф-ии органов(почки,печень)
-тканевый(ф-ции тканей(соед.мышеч))
-клеточный(лейкоциты,микрофаги...)
-субклеточный(ионные и молекулярные основы физиологических мханизмов
Взаимоотношение функции и структуры.
- Структура - это совокупность устойчивых связей элементов сис-
темы, обеспечивающей ее целостность при разных внешних и внут-
ренних изменениях.
- Функциональные изменения при этом можно рассматривать как
выражение внутренних перестроек структуры живого. Структура и
функция составляют динамическое единство.
Нервно-рефлекторные механизмы регуляции функций.
Структурной основой рефлекса является рефлекторная дуга — последовательно соединенная цепочка нервных клеток, обеспечивающая осуществление реакции, или ответа, на раздражение. Рефлекторная дуга состоит из афферентного, центрального и эфферентного звеньев, связанных между собой синаптическими соединениями (рис. 4.1). Афферентная часть дуги начинается рецепторными образованиями, назначение которых заключается в трансформации энергии внешних раздражений в энергию нервного импульса, поступающего по афферентному звену дуги рефлекса в центральную нервную систему.
В зависимости от сложности структуры рефлекторной дуги различают моно- и полисинаптические рефлексы. В простейшем случае импульсы, поступающие в центральные нервные структуры по афферентным путям, переключаются непосредственно на эфферентную нервную клетку, т. е. в системе рефлекторной дуги имеется одно синаптическое соединение. Такая рефлекторная дуга называется моносинаптической (например, рефлекторная дуга сухожильного рефлекса в ответ на растяжение). Наличие в структуре рефлекторной дуги двух и более синаптических переключений (т. е. три и более нейронов), позволяет характеризовать ее как полисинаптическую.
Объяснение физиологических механизмов обучения, приобретенного навыка, коррекции выполненного ответа на основе принципа рефлекторной реакции неверно, неточно и потребовало существенного пересмотра классической схемы рефлекторной реакции, определяемой простой прямой связью: стимул à нервный центр à реакция.
Представление о рефлекторной реакции как о целесообразном ответе организма диктует необходимость дополнить рефлекторную дугу еще одним звеном — петлей обратной связи, призванной установить связь между реализованным результатом рефлекторной реакции и нервным центром, выдающим исполнительные команды. Обратная связь трансформирует открытую рефлекторную дугу в закрытую. Она может быть реализована разными способами: от исполнительной структуры к нервному центру (промежуточному или эфферентному двигательному нейрону), например, через возвратную аксонную коллатераль пирамидного нейрона коры больших полушарий или двигательной моторной клетки переднего рога спинного мозга. Обратная связь может обеспечиваться и нервными волокнами, поступающими к рецепторным структурам и управляющими чувствительностью рецепторных афферентных структур анализатора. Такая структура рефлекторной дуги превращает ее в самонастраивающийся нервный контур регуляции физиологической функции, совершенствуя рефлекторную реакцию и, в целом, оптимизируя поведение организма.
Сложные формы высшей нервной деятельности представлены психическими реакциями, возникающими на основе интеграции элементарных условных рефлексов и аналитико-синтетических механизмов абстрагирования. Абстрагирование от конкретного содержания безусловных подкрепляющих раздражителей обеспечивает возможность более полного и целостного восприятия окружающего мира, адекватного прогнозирования и программирования поведения. В качестве вызывающих подобные реакции стимулов обычно выступают сложные комплексные раздражители. Часто такие рефлекторные реакции имеют усеченную рефлекторную дугу (отсутствует эфферентное звено рефлекторной дуги). Сложные формы высшей нервной деятельности оказываются связанными с синтетическими процессами, обеспечивающими целостные субъективные образы внешнего мира, целенаправленные программы поведения, различные проявления абстрактной мыслительной деятельности человека (психонервная деятельность, рассудочное мышление, функции второй сигнальной системы).
Виды возбудимых тканей, их функциональные свойства.
_-Нерная
Основными функциональными свойствами нервной ткани являются возбудимость и проводимость. Эти свойства нервной ткани обеспечиваются изменением заряда (разности потенциалов между наружной и внутренней поверхностями) плазматической мембраны нервных клеток. Снижение заряда мембраны ниже уровня заряда невозбужжденной мембраны называется деполяризацией и вызывает возбуждение. После возбуждения следует реполяризация мембраны и возвращение уровня разности потенциалов к исходному уровню. Повышение заряда мембраны выше нормального уровня (уровня заряда невозбужжденной мембраны) называется гиперполяризацией и вызывает торможение. Явления возбуждения и торможения сменяют друг друга.
Нервная ткань обеспечивает генерацию возбуждения в ответ на раздражение. При этом энергия внешнего воздействия любого типа "преобразуется" в энергию электрического возбуждения мембраны нервной клетки. Таким образом, информация стимула переводится на язык, который используется в нервной системе.
Этот процесс осуществляется в чувствительных (рецепторных) структурах. При этом (за исключением фоторецепторных клеток) происходит деполяризация мембраны и возникает градуальный рецепторный потенциал, который затем преобразуется в потенциал действия (нервный импульс), распространяющийся по аксонам нейронов.
Возбуждение передается следующему звену нервной цепи через специальные контакты (синапсы). При этом в следующем звене происходит деполяризация мембраны и возникает ее возбуждение. Однако, нейроны могут вызывать не только возбуждение, но и торможение (гиперполяризацию) следующего звена цепи.
Воспринимаемые нейроном возбуждающие и тормозящие сигналы суммируются на поверхности мембраны нейрона. Если результирующее суммарное возбуждение (деполяризация) плазматической мембраны нейрона достигает определенной пороговой величины, то в области аксонального холмика генерируется потенциал действия (нервный импульс).
_-Мышечная
Основными свойствами мышечной ткани является возбудимость и сократимость. Эти свойства мышечной ткани определяют ее основную функцию - обеспечение двигательных реакций организма. Выделяют три вида мышечной ткани:
Гладкая мышечная ткань состоит из одноядерных клеток - миоцитов веретеновидной формы длиной 20 – 500 мкм. Их цитоплазма в световом микроскопе выглядит однородно, без поперечной исчерченности. Эта ткань обладает особыми свойствами: она медленно сокращается и расслабляется, обладает автоматией, является непроизвольной (т.е. ее деятельность не управляется по воле человека). Входит в состав стенок внутренних органов: кровеносных и лимфатических сосудов, мочевыводящих путей, пищеварительного тракта.
Поперечно-полосатая скелетная мышечная ткань состоит из миоцитов, имеющих большую длину (до нескольких сантиметров) и диаметр около 1 микрона; эти клетки многоядерные, содержат до 100 и более ядер; в световом микроскопе цитоплазма выглядит как чередование темных и светлых полосок. Свойствами этой мышечной ткани является высокая скорость сокращения и расслабления и произвольность (т.е. ее деятельность управляется по воле человека). Эта мышечная ткань входит в состав скелетных мышц, а также в стенку глотки, верхней части пищевода, ею образован язык, глазодвигательные мышцы.
Поперечно-полосатая сердечная мышечная ткань состоит из многоядерных кардиомиоцитов, имеющих поперечную исчерченность цитоплазмы. Кардиомиоциты разветвлены и образуют между собой соединения – вставочные диски, в которых объединяется их цитоплазма. Этот вид мышечной ткани образует миокард сердца. Особым свойством этой ткани является автоматия - способность ритмично сокращаться и расслабляться под действием возбуждения, возникающего в самих клетках. Эта ткань является непроизвольной.
-Железистая
синтез и секреция в-ва
Строение мембраны возбудимой ткани, ее функции.
тонкая липопротеиновая пластинка.Липидов 40% беоков 60%
Изнутри выстлана тонким слоем гиалоплазмы,практически лишённой
органелл.На внешней стороне мембраны !0% углеводов,которые
соеденины либо с белками,либо с липидами.и образуют гипокаликс
Структурная основа мембраны-бимолекулярный слой фосфолипидов.
Молекулы фофолипидов мембраны состоят из 2х частей:одна заряжена и
гидрофобна,другая не заряжена и гидрофильна.
Белки: поверхностные,погружённые,пронизывающие
1.Барьерная функция выражается в том, что мембрана при помощи соответствующих механизмов участвует в создании концентрационных градиентов, препятствуя свободной диффузии. При этом мембрана принимает участие в механизмах электрогенеза. К ним относятся механизмы создания потенциала покоя, генерация потенциала действия, механизмы распространения биоэлектрических импульсов по однородной и неоднородной возбудимым структурам.
2.Регуляторная функция клеточной мембраны заключается в тонкой регуляции внутриклеточного содержимого и внутриклеточных реакций за счет рецепции внеклеточных биологически активных веществ, что приводит к изменению активности ферментных систем мембраны и запуску механизмов вторичных «месенджеров» («посредников»).
3.Преобразование внешних стимулов неэлектрической природы в электрические сигналы (в рецепторах).
4.Высвобождение нейромедиаторов в синаптических окончаниях.
Иошше каналы, их виды, функции, ионные насосы.
Итак, ионный канал - это интегральный белок, образующий в мембране пору для обмена клетки с окружающей средой ионами K+, Na+, H+, Ca2+, Cl-, а также водой, и способный изменять свою проницаемость.
Аквапорины - водные неионные каналы мембраны
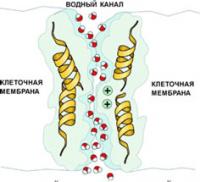
В мембране существуют и неионные каналы. Например,аквапорины — это специальные водные каналы, пропускающие через себя воду. Это тожемембранные каналы, хотя их формально нельзя назвать "ионными каналами".
Пространственная структура канального белка-аквапорина представляет собой цилиндрический канал, по которому движутся молекулы воды. Через него проходит вода и только вода, но не ионы. Аминокислоты в этом белке расположены таким образом, что полярность создаваемого ими электростатического поля переключается в центре канала на обратную. Поэтому молекулы воды, дойдя до середины канала, переворачиваются так, что их дипольные моменты в верхней и нижней части канала оказываются направленными в противоположные стороны. Такое переориентирование предотвращает просачивание через канал заряженных ионов. Аквапорин не пропускает даже ионы гидроксония H3O+ (то есть гидратированные протоны, или ионы водорода), от концентрации которых зависит кислотность среды. При этом клеточный мембранный "водопровод" обладает потрясающей пропускной способностью: он пропускает до миллиарда молекул воды в секунду. Сейчас известно уже около 200 разновидностей белков водных каналов у растений и животных, в том числе 11 - у человека. Благодаря аквапоринам клетки не только регулируют свой объём и внутреннее давление, но и выполняют такие важные функции, как всасывание воды в почках животных и корешках растений.
В настоящее время в молекулярной биологии в основном завершён описательный период в исследовании многообразия катион-транспортирующих ионных каналов в клетках эукариот. Теперь на первый план выходят проблемы познания механизмов регуляции ионных каналов и описание их участия в реакциях живой клетки на различные воздействия и на изменение её микроокружения.
Регулирумый перенос ионов через гидрофильные поры мембраны с помощью управляемых ИК является важнейшим свойством живых клеток, как электровозбудимых, так и невозбудимых.
В связи с этим целесообразно использовать в классификации ионных каналов именно принцип управления их деятельностью. Принцип управления состоянием ионных каналов и был положен в основу предложенной нами (Сазонов В.Ф., 2011.) функциональной классификации ионных каналов.
Строение ИК
ИК состоят из белков сложной структуры (белков-каналоформеров). Схематические изображения ИК приведены ниже, например: натриевый НАХ-рецепторный ионный канал.
На рисунке справа показан натриевый канал: вид сверху, с наружной стороны мембраны (Источник: Horn R. (2011).Peering into the spark of life. Nature 475, 305–306).
Белки ИК имеют определённую конформацию, образующую трансмембранную пору, и "вшиты" в липидный слой мембраны. Канальный белковый комплекс может состоять либо из одной белковой молекулы, либо из нескольких белковых субъединиц, одинаковых или разных по строению. Эти субъединицы могут кодироваться разными генами, синтезироваться на рибосомах по-отдельности и затем собираться в виде целостного канала. В другом случае канал может представлять собой единый полипептид, который в виде петель прошивает мембрану несколько раз. На начало XXI века известно более 400 белков-каналоформеров, для биосинтеза которых используется 1-2% генома человека.
Домены - это отдельные компактно оформленные части канального белка или субъединиц. Сегменты - это части белкка-каналоформера, свёрнутые спирально и прошивающие мембрану. Концевые домены белка-каналоформера (N- и С-терминальные домены) могут торчать из мембраны как наружу, так и внутрь клетки.
Практически все ИК имеют в составе своих субъединиц регуляторные домены, способные связываться с различными управляющими веществами (регуляторными молекулами) и за счёт этого менять состояние или свойства канала. В потенциал-активируемых ИК один из трансмембранных сегментов содержит специальный набор аминокислот с положительными зарядами и работает как сенсор электрического потенциала мембраны. При изменении потенциала такой сенсор меняет состояние канала с открытого на закрытое или наоборот. Таким образом, ИК могут управляться определёнными воздействиями извне, это важное их свойство.
ИК в своём составе могут иметь также вспомогательные субъединицы, выполняющие модуляторные, структурные или стабилизирующие функции. Один класс таких субъединиц - внутриклеточные, расположенные полностью в цитоплазме, а второй - мембранные, т.к. они имеют трансмембранные домены, прошивающие мембрану.
По структуре ИК возможно провести их классификацию, о чём будет сказано ниже.
Свойства ИК
Селективность - это избирательная повышенная проницаемость ИК для определённых ионов. Для других ионов проницаемость понижена. Такая избирательность определяетсяселективным фильтром - самым узким местом канальной поры. Фильтр, кроме узких размеров, может иметь также локальный электрический заряд. Например, катион-селективные каналы обычно имеют в области своего селективного фильтра отрицательно заряженные остатки аминокислот в составе белковой молекулы, которые притягивают положительные катионы и отталкивают отрицательные анионы, не пропуская их через пору.
Управляемая проницаемость - это способность ИК открываться или закрываться при определённых управляющих воздействиях на канал. Понятно, что закрытый канал имеет пониженную проницаемость, а открытый - повышенную. По этому свойству ИК можно классифицировать в зависимости от способов их открытия: например, потенциал-активируемые, лиганд-активируемые и т.д.
Инактивация - это способность ИК через некоторое время после своего открытия автоматически понижать свою проницаемость даже в том случае, когда открывший их активирующий фактор продолжает действовать.
Быстрая инактивация - это особый процесс со своим особым механизмом, отличающийся от медленного закрытия канала (медленной инактивации). Закрытие (медленная инактивация) канала происходит за счёт процессов, противоположных процессам, обеспечившим его открытие, т.е. за счёт изменения конформации канального белка. А вот, например, у потенциал-активируемых каналов быстрая инактивация происходит с помощью специальной молекулярной "пробки-затычки", напоминающей пробку на цепочке, которую обычно используют в ваннах. Эта пробка представляет собой аминокислотную (полипептидную) петлю с утолщением на конце в виде трёх аминокислот, которым и затыкается внутреннее устье канала со стороны цитоплазмы. Именно поэтому потенциал-зависимые ИК для натрия, обеспечивающие развитие потенциала действия и движение нервного импульса, могут пропускать в клетку ионы натрия только в течение нескольких миллисекунд, а затем они автоматически закрываются своими молекулярными пробками, несмотря на то, что открывающая их деполяризация продолжает действовать. Другим механизмом инактивации ИК может служить модификация дополнительными субъединицами внутриклеточного устья канала.
Блокировка - это способность ИК под действием веществ-блокаторов фиксировать какое-то одно своё состояние и не реагировать на обычные управляющие воздействия. В таком состоянии канал просто перестаёт давать ответы на управляющие воздействия. Блокировку вызывают вещества-блокаторы, которые могут называться антагонистами, блокаторамиили литиками.
Антагонисты - это вещества, препятствующие активирующему действию других веществ на ИК. Такие вещества способны хорошо связываться с рецепторным участком ИК, но не способны изменить состояние канала, вызвать его ответную реакцию. Получается блокада рецептора и вместе с ним - блокада ИК. Следует помнить, что антагонисты не обязательно вызывают полную блокаду рецептора и его ИК, они могут действовать более слабо и лишь ингибировать (угнетать) работу канала, но не прекращать её полностью
Агонисты-антагонисты - это вещества, которые обладают слабым стимулирующим влиянием на рецептор, но при этом блокируют действие естественных эндогенных управляющих веществ.
Блокаторы - это вещества, препятствующее работе ионного канала, например, взаимодействию медиатора с молекулярным рецептором к нему и, следовательно, нарушающие управление каналом, блокирующие его. Например, действие ацетилхолина блокируют холиноблокаторы; норадреналина с адреналином - адреноблокаторы; гистамина - гистаминоблокаторы и т. д. Многие блокаторы применяются в терапевтических целях как лекарственные препараты.
Литики - это те же блокаторы в отношении ИК, но этот термин более старый. Он используется как синоним для блокатора: холинолитик, адренолитик и т.д. В то же время в фармакологии термин "литик" применяется в более широком смысле - как вещество, препятствующее не только действию лиганда, а вообще препятствующее передаче возбуждения.
Пластичность - это способность ИК изменять свои свойства, свои характеристики. Наиболее распространённый механизм, обеспечивающий пластичность - это фосфорилирование аминокислот канальных белков с внутренней стороны мембраны ферментами-протеинкиназами. К канальным белкам присоединяются фосфорные остатки от АТФ или ГТФ - и канал меняет свои свойства. Накпример, фиксируется в постоянно закрытом состоянии, или, наоборот, в открытом.
Функции ИК
Главная функция ИК - обеспечивать управляемое перемещение ионов через мембрану.
В зависимости от проходящих через них ионов ИК подразделяют на натриевые, калиевые, кальциевые, хлорные, протонные (водородные).
Функции ИК:
1. Регуляция водного обмена клетки: объём и тургор.
2. Регуляция pH: закисление и защелачивание.
3. Регуляция ионного обмена (обмен солей): изменение внутриклеточного ионного состава и концентрации.
4. Создание и изменение мембранных потенциалов: потенциал покоя; в возбудимых клетках - локальные потенциалы, потенциал действия.
5. Проведение возбуждения в возбудимых клетках: обеспечение движения нервных импульсов.
6. Трансдукция в сенсорных рецепторах: преобразование раздражения (стимула) в возбуждение.
7. Управление активностью клетки: за счёт обеспечения потоков вторичного мессенджера - Са2+.
Функциональные состояния ИК
1. Открытое. Канал открыт и через него происходит перемещение ионов.
2. Закрытое. Канал закрыт и ионы не проходят через него.
3. Активированное. Канал может выполнять свои функции, т.е. открываться и закрываться под действием его регуляторов (управляющих веществ или электрических потенциалов).
4. Инактивированное. Канал не может выполнять свои функции, т.е. открываться и закрываться, он "фиксируется" в каком-то одном состоянии.
5. Блокированное. Канал перекрыт, инактивирован веществом-антагонистом (блокатором), занявшем место управляющего вещества.
6. Модулированное (фосфорилированное). Канал изменяет свои обычные свойства под действием фосфорилирования - присоединения к какому-то его участку фосфатного остатка.
Ионные насосы
ИОННЫЕ НАСОСЫ
молекулярные структуры, встроенные в биол. мембраны и осуществляющие перенос ионов в сторону более высокого электрохим. потенциала (активный транспорт); функционируют за счёт энергии гидролиза АТФ или энергии, высвобождающейся в ходе переноса электронов по дыхат. цепи. Активный транспорт ионов лежит в основе биоэнергетики клетки, процессов клеточного возбуждения, всасывания, а также выведения веществ из клетки и организма в целом. Перенос ионов при гидролизе АТФ обеспечивается транспортными ферментами аденозинтрифосфатазами (АТФазами), к к-рым относятся Н+-АТФаза мембран митохондрий, хлоропластов и бактериальных клеток, Са+-АТФаза внутриклеточных мембран мышечных клеток (мембран саркоплазматич. ретикулума) и эритроцитов и Na+/K+-ATOa3a, содержащаяся практически во всех плазматич. мембранах.. На каждую молекулу гидролизованной АТФ эти АТФазы переносят через мембрану соответственно 2Н+, 2Са2+, 2К+ и 3Na+, причём протоны переносятся из митохондрий и хлоропластов в цитоплазму, ионы Са2+ — из цитоплазмы в пузырьки саркоплазматич. ретикулума и внеклеточное пространство, ионы К+ — в клетку, а ионы Na+ из клетки. В результате создаётся неравновесное распределение ионов и генерируется разность электрич. потенциалов на мембране. При этом происходит запасание энергии, к-рая м. б. в принципе использована для синтеза АТФ, а также для генерации потенциалов действия в нервных и мышечных клетках, для сопряжённого с пассивным транспортом Na активного (вторичного) транспорта аминокислот, углеводов и др. Цикл работы трансп. АТФаз включает связывание АТФ и ионов на поверхности фермента, фосфорилирование фермента, перенос ионов через мембрану, отщепление ортофосфата от белка, изменение прочности связи ионов с ферментом, возврат системы в исходное состояние. Процессы перемещения ионов через мембраны связаны с изменением пространств, структуры белковой части трансп. АТФаз, однако детальный механизм процесса пока не установлен. По-видимому, в состав АТФаз входят ферментный центр, ионный канал и какие-то структурные элементы, препятствующие обратной утечке ионов во время работы И. н. В мембранах митохондрий, хлоропластов и клеток бактерий функционирует и др. механизм активного переноса протонов: сопряжение трансмембранного переноса протонов с переносом электронов через оцредел. участки окислитель-но-восстановит. системы — цепи транспорта электрона. Работа таких систем вместе с работой обращенной Н+-АТФазы приводит к окислительному фосфорили-рованию в митохондриях и у бактерий и к фотосинтетич. фосфорилированию в хлоропластах растений и хроматофорах фотосинтезирующих бактерий. (см. ХЕМИОСМОТИЧЕСКАЯ ТЕОРИЯ). Нарушение работы И. н. сопровождается развитием патологич. состояний в организме (так, необратимое повреждение клеток при недостатке кислорода связано с выключением трансп. АТФаз из-за отсутствия АТФ в условиях тканевой гипоксии). Нек-рые лекарств, препараты, напр. сердечные гликозиды, могут регулировать активность И. н.
Мембранный потенциал, механизмы его формирования.