

Голандрический тип наследования. Сцепленный с хромосомой y тип наследования.
Гены, ответственные за развитие патологического признака, локализованы в хромосоме Y.
Примеры признаков: гипертрихоз ушных раковин, избыточный рост волос на средних фалангах пальцев кистей, азооспермия.
Родословная с Y-сцепленным типом наследования избыточного оволосения ушных раковин в четырёх поколениях представлена на рисунке.
Родословная с Y-сцепленным (голандрическим) типом наследования. Кружок— пол женский, квадрат— пол мужской, тёмный кружок и/или квадрат — больной.
37, 38
Хромосомная теория наследственности— теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности, то есть преемственность свойств организмов в ряду поколений определяется преемственностью их хромосом. Хромосомная теория наследственности возникла в начале 20 в. на основе клеточной теории и использовалась для изучения наследственных свойств организмов гибридологического анализа.
Формирование хромосомной теории
В 1902-1903 гг. американский цитолог У. Сеттон и немецкий цитолог и эмбриолог Т. Бовери независимо друг от друга выявили параллелизм в поведении генов и хромосом в ходе формирования гамет и оплодотворения. Эти наблюдения послужили основой для предположения, что гены расположены в хромосомах. Однако экспериментальное доказательство локализации конкретных генов в конкретных хромосомах было получено только в 1910 г. американским генетиком Т. Морганом, который в последующие годы (1911—1926) обосновал хромосомную теорию наследственности. Согласно этой теории, передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Таким образом, именно хромосомы представляют собой материальную основу наследственности.
Формированию хромосомной теории способствовали данные, полученные при изучении генетики пола, когда были установлены различия в наборе хромосом у организмов различных полов.
Генетика пола
Пол, как и любой другой признак организма, наследственно детерминирован. Важнейшая роль в генетической детерминации пола и в поддержании закономерного соотношения полов принадлежит хромосомному аппарату.
У раздельнополых организмов (животных и двудомных растении) соотношение полов обычно составляет 1:1, то есть мужские и женские особи встречаются одинаково часто. Это соотношение совпадает с расщеплением в анализирующем скрещивании, когда одна из скрещиваемых форм является гетерозиготной (Аа), а другая — гомозиготной по рецессивным аллелям (аа). В потомстве в этом случае наблюдается расщепление в отношении 1Аа:1аа. Если пол наследуется по такому же принципу, то вполне логично было бы предположить, что один пол должен быть гомозиготным, а другой — гетерозиготным. Тогда расщепление по полу должно быть в каждом поколении равным 1:1, что и наблюдается в действительности.
При изучении хромосомных наборов самцов и самок ряда животных между ними были обнаружены некоторые различия. Как у мужских, так и у женских особей во всех клетках имеются пары одинаковых (гомологичных) хромосом, но по одной паре хромосом они различаются. Так, у самки дрозофилы имеются две палочковидные хромосомы, а у самца — одна такая же палочковидная, а вторая, парная первой, — изогнутая. Такие хромосомы, по которым самцы и самки отличаются друг от друга, называют половыми хромосомами. Те из них, которые являются парными у одного из полов, называют X-хромосомами (например, у дрозофилы и млекопитающих) или Z-хромосомами (например, у птиц). Непарная половая хромосома, имеющаяся у особей только одного пола, была названа У-хромосомой (у дрозофилы и млекопитающих) или W-хромосомой (у птиц). Хромосомы, в отношении которых между самцами и самками нет различий, называют аутосомами. Следовательно, у дрозофилы особи обоих полов имеют по шесть одинаковых аутосом плюс две половые хромосомы (ХХ у самок и XY у самцов).
Пол, имеющий различные половые хромосомы (X и У), образует гаметы двух типов (половина с X-хромосомой и половина с У-хромосомой), то есть, является гетерогаметным, а пол, содержащий в каждой клетке одинаковые половые хромосомы (X-хромосомы), — гомогаметным.
Открытие половых хромосом и установление их роли в определении пола послужило важным доводом в пользу того, что хромосомы определяют признаки организма.
Определение пола
От чего же зависит рождение мужских и женских особей? Рассмотрим это на примере определения пола у дрозофилы. В ходе гаметогенеза у самок образуется один тип гамет, содержащий гаплоидный набор аутосом и одну X-хромосому. Самцы образуют два типа гамет, половина из которых содержит три аутосомы и одну X-хромосому (ЗА+Х), а половина — три аутосомы и одну У-хромосому (ЗА+У). При оплодотворении яйцеклеток (ЗА+Х) сперматозоидами с X-хромосомами будут формироваться самки (6А+ХХ), а от слияния яйцеклеток со сперматозоидами, несущими У-хромосому, — самцы (6A+XY). Поскольку число мужских гамет с X- и У-хромосомами одинаково, то и количество самцов и самок тоже одинаково. В данном случае пол организма определяется в момент оплодотворения и зависит от хромосомного набора зиготы.
Сходный способ определения пола (XY-тип) присущ всем млекопитающим, в том числе и человеку, клетки которого содержат 44 аутосомы и две X-хромосомы у женщин либо XY-хромосомы у мужчин.
Таким образом, XY-тип определения пола, или тип дрозофилы и человека, — самый распространенный способ определения пола, характерный для большинства позвоночных и некоторых беспозвоночных. Х0-тип встречается у большинства прямокрылых, клопов, жуков, пауков, у которых Y-хромосомы нет вовсе, так что самец имеет генотип Х0, а самка — XX.
У всех птиц, большинства бабочек и некоторых пресмыкающихся самцы являются гомогаметным полом, а самки —- гетерогаметным (типа XY или типа ХО). Половые хромосомы у этих видов обозначают буквами Z и W, чтобы выделить таким образом данный способ определения пола; при этом набор хромосом самцов обозначают символом ZZ, а самки — символом ZW или Z0.
Доказательства того, что половые хромосомы определяют пол организма, были получены при изучении нерасхождения половых хромосом у дрозофилы. Если в одну из гамет попадут обе половые хромосом, а в другую — ни одной, то при слиянии таких гамет с нормальными могут получиться особи с набором половых хромосом ХХХ, ХО, ХХУ и др. Выяснилось, что у дрозофилы особи с набором ХО — самцы, а с набором ХХУ — самки (у человека — наоборот). Особи с набором ХХХ имеют гипертрофированные признаки женского пола (сверхсамки). (Особи со всеми этими хромосомными аберрациями у дрозофилы стерильны). В дальнейшем было доказано, что у дрозофилы пол определяется соотношением (балансом) между числом X-хромосом и числом наборов аутосом.
Наследование признаков, сцепленных с полом
В том случае, когда гены, контролирующие формирование того или иного признака, локализованы в аутосомах, наследование осуществляется независимо от того, кто из родителей (мать или отец) является носителем изучаемого признака. Если же гены находятся в половых хромосомах, характер наследования признаков резко изменяется. Например, у дрозофилы гены, локализованные в X-хромосоме, как правило, не имеют аллелей в У-хромосоме. По этой причине рецессивные гены в X-хромосоме гетерогаметного пола практически всегда проявляются, будучи в единственном числе.
Признаки, гены которых локализованы в половых хромосомах, называются признаками, сцепленными с полом. Явление наследования, сцепленного с полом, было открыто Т. Морганом у дрозофилы.
Х- и У-хромосомы у человека имеют гомологичный (псевдоаутосомный) участок, где локализованы гены, наследование которых не отличается от наследования аутосомных генов.
Помимо гомологичных участков, X- и У-хромосомы имеют негомологичные участки. Негомологичный участок У-хромосомы, кроме генов, определяющих мужской пол, содержит гены перепонок между пальцами ног и волосатых ушей у человека. Патологические признаки, сцепленные с негомологичным участком У-хромосомы, передаются всем сыновьям, поскольку они получают от отца У-хромосому.
Негомологичный участок X-хромосомы содержит в своем составе ряд важных для жизнедеятельности организмов генов. Поскольку у гетерогаметного пола (ХУ) X-хромосома представлена в единственном числе, то признаки, определяемые генами негомологичного участка X-хромосомы, будут проявляться даже в том случае, если они рецессивны. Такое состояние генов называется гемизиготным. Примером такого рода X-сцепленных рецессивных признаков у человека являются гемофилия, мышечная дистрофия Дюшена, атрофия зрительного нерва, дальтонизм (цветовая слепота) и др.
Гемофилия — это наследственная болезнь, при которой кровь теряет способность свертываться. Ранение, даже царапина или ушиб, могут вызвать обильные наружные или внутренние кровотечения, которые нередко заканчиваются смертью. Это заболевание встречается, за редким исключением, только у мужчин. Было установлено, что обе наиболее распространенные формы гемофилии (гемофилия А и гемофилия В) обусловлена рецессивными генами, локализованными в X-хромосоме. Гетерозиготные по данным генам женщины (носительницы) обладают нормальной или несколько пониженной свертываемостью крови.
Фенотипическое проявление гемофилии у девочек будет наблюдаться в том случае, если мать девочки является носительницей гена гемофилии, а отец — гемофиликом. Подобная закономерность наследования характерна и для других рецессивных, сцепленных с полом признаков.
Сцепленное наследование
Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом. Например, у кукурузы до эры молекулярной биологии было изучено более 500 генов, у мухи дрозофилы — более 1 тыс., а у человека — около 2 тыс. генов, тогда как хромосом у них 10, 4 и 23 пары соответственно. То, что число генов у высших организмов составляет несколько тысяч, было ясно уже У. Сэттону в начале XX века. Это дало основание предположить, что в каждой хромосоме локализовано множество генов. Гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются вместе.
Совместное наследование генов Т. Морган предложил назвать сцепленным наследованием. Число групп сцепления соответствует гаплоидному числу хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены. (У особей гетерогаметного пола, например, у самцов млекопитающих, групп сцепления на самом деле на одну больше, так как X- и У-хромосомы содержат разные гены и представляют собой две разные группы сцепления. Таким образом, у женщин 23 группы сцепления, а у мужчин — 24).
Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигетерозиготная особь образует четыре типа гамет (АВ, Ab, аВ и ab) в равных количествах, то при сцепленном наследовании (в отсутствие кроссинговера) такая же дигетерозигота образует только два типа гамет: (АВ и ab) тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме родителя.
Было установлено, однако, что кроме обычных (некроссоверных) гамет возникают и другие (кроссоверные) гаметы с новыми комбинациями генов — Ab и аВ, отличающимися от комбинаций генов в хромосомах родителя. Причиной возникновения таких гамет является обмен участками гомологичных хромосом, или кроссинговер.
Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом. В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, получили название кроссинговерных или рекомбинантных.
Частота (процент) перекреста между двумя генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они расположены. По мере увеличения расстояния между генами все более возрастает вероятность того, что кроссинговер разведет их по двум разным гомологичным хромосомам.
Расстояние между генами характеризует силу их сцепления. Имеются гены с высоким процентом сцепления и такие, где сцепление почти не обнаруживается. Однако при сцепленном наследовании максимальная частота кроссинговера не превышает 50 %. Если же она выше, то наблюдается свободное комбинирование между парами аллелей, не отличимое от независимого наследования.
Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость, которая дает широкие возможности адаптации организма в различных условиях среды. Человек специально проводит гибридизацию с целью получения необходимых вариантов комбинаций для использования в селекционной работе.
Понятие о генетической карте
Т. Морган и его сотрудники К. Бриджес, А. Г. Стертевант и Г. Дж. Меллер экспериментально показали, что знание явлений сцепления и кроссинговера позволяет не только установить группу сцепления генов, но и построить генетические карты хромосом, на которых указаны порядок расположения генов в хромосоме и относительные расстояния между ними.
Генетической картой хромосом называют схему взаимного расположения генов, находящихся в одной группе сцепления. Такие карты составляются для каждой пары гомологичных хромосом.
Возможность подобного картирования основана на постоянстве процента кроссинговера между определенными генами. Генетические карты хромосом составлены для многих видов организмов: насекомых (дрозофила, комар, таракан и др.), грибов (дрожжи, аспергилл), для бактерий и вирусов.
Наличие генетической карты свидетельствует о высокой степени изученности того или иного вида организма и представляет большой научный интерес. Такой организм является прекрасным объектом для проведения дальнейших экспериментальных работ, имеющих не только научное, но и практическое значение. В частности, знание генетических карт позволяет планировать работы по получению организмов с определенными сочетаниями признаков, что теперь широко используется в селекционной практике. Так, создание штаммов микроорганизмов, способных синтезировать необходимые для фармакологии и сельского хозяйства белки, гормоны и другие сложные органические вещества, возможно только на основе методов генной инженерии, которые, в свою очередь, базируются на знании генетических карт соответствующих микроорганизмов.
Генетические карты человека также могут оказаться полезными в здравоохранении и медицине. Знания о локализации гена в определенной хромосоме используются при диагностике ряда тяжелых наследственных заболеваний человека. Уже теперь появилась возможность для генной терапии, то есть для исправления структуры или функции генов.
Сравнение генетических карт разных видов живых организмов способствует также пониманию эволюционного процесса.
Основные положения хромосомной теории наследственности
Анализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности:
Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален.
Аллельные гены занимают одинаковые локусы в гомологичных хромосомах.
Гены расположены в хромосоме в линейной последовательности.
Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцепленно (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола).
Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами).
Каждый биологический вид характеризуется определенным набором хромосом — кариотипом.
39.
Комбинативная изменчивость - это следствие перекреста гомологичных хромосом, их случайного расхождения в мейозе и случайного сочетания гамет при оплодотворении. Комбинативная изменчивость ведет к появлению бесконечно большого разнообразия генотипов и фенотипов. Она служит неиссякаемым источником наследственного разнообразия видов и основой для естественного отбора.
Громадное генотипическое и, следовательно, фенотипическое разнообразие в природных популяциях является тем исходным эволюционным материалом, с которым оперирует естественный отбор.
40. Закономерности наследования внеядерных генов. Цитоплазматическое наследование
Наличие некоторого количества наследственного материала в цитоплазме в виде кольцевых молекул ДНК митохондрий и пластид, а также других внеядерных генетических элементов дает основание специально остановиться на их участии в формировании фенотипа в процессе индивидуального развития. Цитоплазматические гены не подчиняются менделевским закономерностям наследования, которые определяются поведением хромосом при митозе, мейозе и оплодотворении. В связи с тем что организм, образуемый вследствие оплодотворения, получает цитоплазматические структуры главным образом с яйцеклеткой, цитоплазматическое наследование признаков осуществляется по материнской линии. Такой тип наследования был впервые описан в 1908 г. К. Корренсом в отношении признака пестрых листьев у некоторых растений (рис. 6.21).
Как было установлено позднее, развитие этого признака обусловлено мутацией, возникающей в ДНК хлоропластов и нарушающей синтез хлорофилла в них. Размножение в клетках нормальных (зеленых) и мутантных (бесцветных) пластид и последующее случайное распределение их между дочерними клетками приводят к появлению отдельных клеток, совершенно лишенных нормальных пластид. Потомство этих клеток образует обесцвеченные участки на листьях. Фенотип потомства, таким образом, зависит от фенотипа материнского растения. У растения с зелеными листьями потомство абсолютно нормально. У растения с бесцветными листьями потомство имеет такой же фенотип. У материнского растения с пестрыми листьями потомки могут иметь все описанные фенотипы по данному признаку. При этом внешний вид потомства не зависит от признака отцовского растения.
Другим примером цитоплазматического наследования признаков могут служить некоторые патологические состояния, описанные у человека, причиной которых является первичный дефект митохондриальной ДНК (мтДНК) (см. гл. 4.1 и гл. 6.4.1.4).
Наряду с описанными выше типами и вариантами наследования ядерных и цитоплазматических генов в последнее время внимание ученых привлекает нетрадиционное наследование некоторых признаков и патологических состояний у человека (см. гл. 6.4.1.4.).
41.
Модификационная (фенотипическая) изменчивость — изменения в организме, связанные с изменением фенотипа вследствие влияния окружающей среды и носящие, в большинстве случаев, адаптивный характер. Генотип при этом не изменяется. В целом современное понятие «адаптивные модификации» соответствует понятию «определенной изменчивости», которое ввел в науку Чарльз Дарвин.
Условная классификация модификационной изменчивости:
По изменяющимся признакам организма:
морфологические изменения
физиологические и биохимические адаптации — гомеостаз (повышение уровня эритроцитов в горах и т. д.)
По размаху нормы реакции :
узкая (более характерна для качественных признаков)
широкая (более характерна для количественных признаков)
По значению:
модификации (полезные для организма — проявляются как приспособительная реакция на условия окружающей среды)
морфозы (ненаследственные изменения фенотипа под влиянием экстремальных факторов окружающей среды или модификации, возникающие как выражение вновь возникших мутаций, не имеющие приспособительного характера)
фенокопии (различные ненаследственные изменения, копирующие проявление различных мутаций)— разновидность морфозов
По длительности:
есть лишь у особи или группы особей, которые подверглись влиянию окружающей среды (не наследуются)
длительные модификации — сохраняются на два-три поколения
Механизм модификационной изменчивости
Окружающая среда как причина модификаций
Модификационная изменчивость — это результат не изменений генотипа, а его реакции на условия окружающей среды. При модификационной изменчивости наследственный материал не изменяется, — изменяется проявление генов.
Под действием определенных условий окружающей среды на организм изменяется течение ферментативных реакций (активность ферментов) и может происходить синтез специализированных ферментов, некоторые из которых (MAP-киназа и др.) ответственны за регуляцию транскрипции генов, зависящую от изменений окружающей среды. Таким образом, факторы окружающей среды способны регулировать экспрессию генов, то есть интенсивность выработки ими специфических белков, функции которых отвечают специфическим факторам окружающей среды.
Например, за выработку меланина ответственны четыре гена, которые находятся в разных хромосомах. Наибольшее количество доминантных аллелей этих генов — 8 — содержится у людей негроидной расы. При воздействии специфической окружающей среды, например, интенсивного воздействия ультрафиолетовых лучей, происходит разрушение клеток эпидермиса, что приводит к выделению эндотелина-1 и эйкозаноидов. Они вызывают активацию фермента тирозиназы и его биосинтез. Тирозиназа, в свою очередь, катализирует окисление аминокислоты тирозина. Дальнейшее образование меланина проходит без участия ферментов, однако большее количество фермента обуславливает более интенсивную пигментацию.
Норма реакции
Предел проявления модификационной изменчивости организма при неизменном генотипе — норма реакции. Норма реакции обусловлена генотипом и различается у разных особей данного вида. Фактически норма реакции — спектр возможных уровней экспрессии генов, из которого выбирается уровень экспрессии, наиболее подходящий для данных условий окружающей среды. Норма реакции имеет предел для каждого вида — например, усиленное кормление приведет к увеличению массы животного, однако она будет находиться в пределах нормы реакции, характерной для данного вида или породы. Норма реакции генетически детерминирована и наследуется. Для разных изменений есть разные пределы нормы реакции. Например, сильно варьируют величина удоя, продуктивность злаков (количественные изменения), слабо — интенсивность окраски животных и т. д. (качественные изменения).
В соответствии с этим норма реакции может быть широкой (количественные изменения — размеры листьев многих растений, размеры тела многих насекомых в зависимости от условий питания их личинок) и узкой (качественные изменения — окраска у куколок и имаго некоторых бабочек). Тем не менее, для некоторых количественных признаков характерна узкая норма реакции (жирность молока, число пальцев на ногах у морских свинок), а для некоторых качественных признаков — широкая (например, сезонные изменения окраски у многих видов животных северных широт).
Характеристика модификационной изменчивости
-
обратимость — изменения исчезают при смене специфических условий окружающей среды, спровоцировавших их
-
групповой характер
-
изменения в фенотипе не наследуются, наследуется норма реакции генотипа
-
статистическая закономерность вариационных рядов
-
затрагивает фенотип, при этом не затрагивая сам генотип.
42.
Онтогене́з (от греч. οντογένεση: ον — существо и γένεση — происхождение, рождение) — индивидуальное развитие организма от оплодотворения (при половом размножении) или от момента отделения от материнской особи (при бесполом размножении) до смерти.
У многоклеточных животных в составе онтогенеза принято различать фазы эмбрионального (под покровом яйцевых оболочек) и постэмбрионального (за пределами яйца) развития, а у живородящих животных пренатальный (до рождения) и постнатальный (после рождения) онтогенез.
У семенных растений к эмбриональному развитию относят процессы развития зародыша, происходящие в семени.
Термин «онтогенез» впервые был введён Э. Геккелем в 1866 году. В ходе онтогенеза происходит процесс реализации генетической информации, полученной от родителей.
Раздел современной биологии, изучающий онтогенез, называется биологией развития; начальные этапы онтогенеза изучаются также эмбриологией.
Онтогенез животных
Сравнение зародышей позвоночных на разных стадиях эмбрионального развития. Печально знаменитая иллюстрация из работы Эрнста Геккеля, на которой различия между эмбрионами искусственно занижены, ради большего соответствия теории рекапитуляции (повторения филогенеза в онтогенезе). Следует отметить, что фальсификация данной иллюстрации не отменяет того, что зародыши обычно действительно представляются более сходными между собой, чем взрослые организмы, что было отмечено эмбриологами еще до возникновения теории эволюции.
Онтогенез делится на два периода:
эмбриональный — от образования зиготы до рождения или выхода из яйцевых оболочек;
постэмбриональный — от выхода из яйцевых оболочек или рождения до смерти организма.
Эмбриональный период
В эмбриональном периоде выделяют три основных этапа: дробление, гаструляцию и первичный органогенез. Эмбриональный, или зародышевый, период онтогенеза начинается с момента оплодотворения и продолжается до выхода зародыша из яйцевых оболочек. У большинства позвоночных он включает стадии (фазы) дробления, гаструляции, гисто- и органогенеза.
Дробление
Дробление — ряд последовательных митотических делений оплодотворенного или инициированного к развитию яйца. Дробление представляет собой первый период эмбрионального развития, который присутствует в онтогенезе всех многоклеточных животных и приводит к образованию зародыша, называемого бластулой (зародыш однослойный). При этом масса зародыша и его объем не меняются, то есть они остаются такими же, как у зиготы, а яйцо разделяется на все более мелкие клетки — бластомеры. После каждого деления дробления клетки зародыша становятся все более мелкими, то-есть меняются ядерно-плазменные отношения: ядро остается таким же, а объем цитоплазмы уменьшается. Процесс протекает до тех пор, пока эти показатели не достигнут значений, характерных для соматических клеток. Тип дробления зависит от количества желтка и его расположения в яйце. Если желтка мало и он равномерно распределен в цитоплазме (изолецитальные яйца: иглокожие, плоские черви, млекопитающие), то дробление протекает по типу полного равномерного: бластомеры одинаковы по размерам, дробится все яйцо. Если желток распределен неравномерно (телолецитальные яйца: амфибии), то дробление протекает по типу полного неравномерного: бластомеры — разной величины, те, которые содержат желток — крупнее, яйцо дробится целиком. При неполном дроблении желтка в яйцах настолько много, что борозды дробления не могут разделить его целиком. Дробление яйца, у которого дробится только сконцентрированная на анимальном полюсе «шапочка» цитоплазмы, где находится ядро зиготы, называется неполным дискоидальным (телолецитальные яйца: пресмыкающиеся, птицы). При неполном поверхностном дроблении в глубине желтка происходят первые синхронные ядерные деления, не сопровождающиеся образованием межклеточных границ. Ядра, окруженные небольшим количеством цитоплазмы, равномерно распределяются в желтке. Когда их становится достаточно много, они мигрируют в цитоплазму, где затем после образования межклеточных границ возникает бластодерма (центролецитальные яйца: насекомые).
Гаструляция
Гаструляция (впячивание) — гаструла формируется в результате инвагинации клеток. В ходе гаструляции клетки зародыша практически не делятся и не растут. Происходит активное передвижение клеточных масс (морфогенетические движения). В результате гаструляции формируются зародышевые листки (пласты клеток). Гаструляция приводит к образованию зародыша, называемого гаструлой.
Первичный органогенез
Первичный органогенез — процесс образования комплекса осевых органов. В разных группах животных этот процесс характеризуется своими особенностями. Например, у хордовых на этом этапе происходит закладка нервной трубки, хорды и кишечной трубки.
В ходе дальнейшего развития формирование зародыша осуществляется за счет процессов роста, дифференцировки и морфогенеза. Рост обеспечивает накопление клеточной массы зародыша. В ходе процесса дифференцировки возникают различно специализированные клетки, формирующие различные ткани и органы. Процесс морфогенеза обеспечивает приобретение зародышем специфической формы.
Постэмбриональное развитие
Постэмбриональное развитие бывает прямым и непрямым.
Прямое развитие — развитие, при котором появившийся организм идентичен по строению взрослому организму, но имеет меньшие размеры и не обладает половой зрелостью. Дальнейшее развитие связано с увеличением размеров и приобретением половой зрелости. Например: развитие рептилий, птиц, млекопитающих.
Непрямое развитие (личиночное развитие, развитие с метаморфозом) — появившийся организм отличается по строению от взрослого организма, обычно устроен проще, может иметь специфические органы, такой зародыш называется личинкой. Личинка питается, растет и со временем личиночные органы заменяются органами, свойственными взрослому организму (имаго). Например: развитие лягушки, некоторых насекомых, различных червей.
Постэмбриональное развитие сопровождается ростом.
47.
Основы генетики человека
Генетика человека изучает явления наследственности и изменчивости в популяциях людей, особенности наследования нормальных и патологических признаков, зависимость заболевания от генетической предрасположенности и факторов среды.
Задачей медицинской генетики является выявление и профилактика наследственных болезней.
Одним из основоположников медицинской генетики является выдающийся советский невролог С.Н. Давиденков (1880-1961), начинавший свою плодотворную работу в двадцатых годах на Украине. Он впервые применил идеи генетики в клинике, дал анализ ряда наследственных заболеваний, часть из которых была описана им впервые.
Важной заслугой С.Н. Давиденкова является разработка методов медико-генетического консультирования и его первое практическое применение в нашей стране.
Особенности генетики человека
Исследование генетики человека с большими трудностями, причины которых связаны:
•с невозможностью экспериментального скрещивания
•c медленной сменой поколений
•c малым количеством потомков в каждой семье
•c тем, что у человека сложный кариотип, большое число групп сцепления
Однако, несмотря на все эти затруднения, генетика человека успешно развивается. Невозможность экспериментального скрещивания компенсируется тем, что исследователь, наблюдая обширную человеческую популяцию, может брать из тысячи брачных пар те, которые необходимы для генетического анализа. Метод гибридизации соматических клеток позволяет экспериментально изучать локализацию генов в хромосомах, проводить анализ групп сцепления.
При изучении генетики человека используются следующие методы:
•генеалогический
•близнецовый
•популяционно-статистический
•дерматоглифический
•биохимический
•цитогенетический
•гибридизации соматических клеток
•моделирования
Методы изучения наследственности у человека
Генеалогический метод
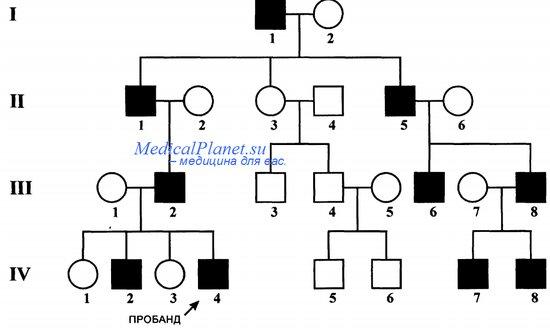
Этот метод основан на прослеживании какого-либо нормального или патологического признака в ряде поколений с указанием родственных связей между членами родословной.
Генеалогический метод является основным связующим звеном между теоретической генетикой человека и применением ее достижений в медицинской практике.
Суть этого метода состоит в том. чтобы выяснить родственные связи и проследить наличие нормального или патологического признака среди близких и дальних родственников в данной семье. Сбор сведений начинается от пробанда. Пробандом называется лицо, родословную которого необходимо составить. Им может быть больной или здоровый человек – носитель какого-либо признака или лицо, обратившееся за советом к врачу-генетику. Братья и сестры пробанда называются сибсами. Обычно родословная составляется по одному или нескольким признакам.
Метод включает два этапа:
•сбор сведений о семье
•генеалогический анализ
Для составления родословной проводят краткие записи о каждом члене родословной с точным указанием его родства по отношению к пробанду. Затем делают графическое изображение родословной. Генеалогический метод тем информативнее, чем больше имеется достоверных сведений о здоровье родственников больного. При собирании генетических сведений и их анализе надо иметь в виду, что признак может быть выражен в разной степени, иногда незначительной – микропризнаки.
После составления родословной начинается второй этап – генеалогический анализ, целью которого является установление генетических закономерностей:
•в начале требуется установить имеет ли признак наследственный характер; если какой-либо признак встречался в родословной несколько раз, то можно думать о его наследственной природе; однако это может быть и не так, например, какие-то внешние факторы или профессиональные вредности могут вызывать сходные заболевания у членов одной семьи
•в случае обнаружения наследственного характера признака необходимо установить тип наследования: доминантный, рецессивный, сцепленный с полом
Основные признаки аутосомно-доминантного наследования:
•проявление признака в равной мере у представителей обоих полов
•наличие больных во всех поколениях (по вертикали) при относительно большом количестве сибсов
•наличие больных и по горизонтали (у сестер и братьев пробанда)
•у гетерозиготного родителя вероятность рождения больного ребенка (если второй родитель здоров) составляет 50%
Следует учесть, что при доминантном типе наследования может быть пропуск в поколениях за счет слабо выраженных, «стертых» форм заболевания (малая эксперссивность мутантного гена) или за счет его низкой пенетрантности (когда у носителя донного гена признак отсутствует).
Основные признаки аутосомно-рецессивного наследования:
•относительно небольшое число больных в родословной
•наличие больных «по горизонтали» (болеют сибсы – родные, двоюродные)
•родители больного ребенка чаще фенотипически здоровы, но являются гетерозиготными носителями рецессивного гена
•вероятность рождения больного ребенка составляет 25%
Рецессивный признак проявляется тогда, когда в генотипе имеются оба рецессивных аллеля.
При проявлении рецессивных заболеваний нередко встречается кровное родство родителей больных. Следует иметь в виду, что наличие отдаленного родства бывает неизвестно членам семьи. Приходиться учитывать косвенные соображения , например, происхождение из одного и того же малонаселенного пункта, или принадлежность к какой-либо изолированной этнической или социальной группе.
Основные признаки наследования, сцепленного с полом:
•заболевания, обусловленные геном, локализованным в Х-хромосоме, могут быть как доминантными, так и рецессивными
•при доминантном Х-сцепленном наследовании заболевание одинаково проявляется как у мужчин, так и у женщин и в дальнейшем может передаваться по потомству (в этом случае женщина может передавать этот ген половине дочерей и половине сыновей)
•при рецессивном наследовании заболеваний, сцепленными с Х-хромосомой, как правило страдают мужчины (гетерозиготная носительница – мать – передает мутантный ген половине сыновей, которые будут больны и половине дочерей, которые оставаясь фенотипически здоровыми, как и мать, тоже являются носительницами и передают рецессивный ген вместе с Х-хромосомой следующему поколению)
Близнецовый метод
Это один из наиболее ранних методов изучения генетики человека, однако он не утратил своего значения и в настоящее время. Близнецовый метод был введен Ф.Гамильтоном, который выделил среди близнецов две группы:
•одняйцевые (монозиготные)
•двуяйцевые (дизиготные)
Монозиготные близнецы при нормальном эмбриональном развитии всегда одного пола. Дизиготные близнецы рождаются чаще (2/3 общего количества двоен), они развиваются из двух одновременно созревших и оплодотворенных яйцеклеток. Такие близнецы могут быть и однополые и разнополые. С генетической точки зрения они сходны как обычные сибсы, но у них большая общность факторов среды во внутриутробном (пренатальном) и частично в постнатальном периодах.
Если изучаемый признак проявляется у обоих близнецов пары, их называют конкордантными. Конкордантность – это процент сходства по изучаемому признаку. Отсутствие признака у одного из близнецов – дискордантность.
Близнецовый метод используется в генетике человека для того, чтобы оценить степень влияния наследственности и среды на развитие какого-либо нормального или патологического признака.
Для оценки роли наследственности в развитии того или иного признака производят расчет по формуле:
Н = (% сходства ОБ - % сходства ДБ) / (100 - % сходства ДБ)
где:
Н- коэффицент наследственности
ОБ – однояйцевые близнецы
ДБ – двуяйцевые близнецы
•При Н = 1 признак полностью определяется наследственным компонентом
•При Н = 0 признак определяется влиянием среды
•При Н = близкий к 0,5 признак определяется примерно одинаковым влиянием наследственности и среды на формирование признака
Метод дерматоглифики
Дерматоглифка – это изучение рельефа кожи на пальцах, ладонях и подошвенных поверхностях стоп, который образован эпидермальными выступами – гребнями, которые образуют сложные узоры.
Ф. Гальтон предложил предложил классификацию этих узоров, позволившую использовать этот метод для идентификации личности в криминалистике.
Разделы дерматоглифики:
•дактилоскопия – изучение узоров на подушечках пальцев
•пальмоскопия – изучение рисунка на ладонях
•плантоскопия – изучение дерматоглифики подошвенной поверхности стопы
Дактилоскопия. Гребни на коже пальцев рук соответствуют сосочкам дермы, поэтому их называют также папиллярными линиями, рельеф этих выступов повторяет пласт эпидермиса. Межсосочковые углубления образуют бороздки. Закладка узоров происходит между 10 и 19 неделями внутриутробного развития; у 20 недельных плодов уже хорошо различимы формы узоров. Формирование папиллярного рельефа зависит от характера ветвления нервных волокон. Полное формирование деталей строения тактильных узоров отмечается к шести месяцам, после чего они остаются неизменными до конца жизни. Дерматоглифические исследования имеют важное значение в определении зиготности близнецов, в диагностике некоторых наследственных заболеваний, в судебной медицине, в криминалистике для идентификации личности.
Пальмоскопия. Ладонный рельеф очень сложный, в нем выделяют ряд полей, подушечек и ладонных линий. У правшей более сложные узоры встречаются на правой руке, у левшей – на левой. Индивидуальные особенности кожных узоров наследственно обусловлены. Это доказано многими генетическими исследованиями, в частности, на монозиготных близнецах.
Обширные исследования по изучению особенностей дерматоглифики проведены у нас в стране Т.Д. Гладковой (1996), а по наследственной обусловленности кожных узоров – И.С. Гусевой (1970, 1980). на основании этих работ был сделан вывод, что количественные показатели рельефа гребневой кожи программируются полигенной системой, включающей небольшое число аддитивно действующих генов. гены гребневой кожи проявляют свой морфогенетический эффект, влияя на степень ветвления нервного волокна, и фенотипически определяют гребневую плотность. На формирование дерматоглифических узоров могут оказывать влияние некоторые повреждающие факторы на ранних стадиях эмбрионального развития.
Биохимические методы
Эти методы используются для диагностики болезней обмена веществ, причиной которых является изменение активности определенных ферментов. С помощью биохимических методов открыто около 500 молекулярных болезней, являющихся следствием проявления мутантных генов. Эти методы отличаются большой трудоемкостью, требуют специального оборудования и поэтому не могут быть широко использованы для массовых популяционных исследований с целью раннего выявления больных с наследственной патологией обмена.
В последние десятилетия в разных странах разрабатываются и применяются для массовых исследований специальные программы:
•первый этап такой программы состоит в том, чтобы среди большого количесества обследуемых выделить предположительно больных, имеющих какое-то наследственное отклоенение от нормы. такая программа называется просеивающей, или скрининг-программой. для этого этапа обычно используется небольшое количество простых, доступных методик (экспересс-методов).
•второй этап проводится с целью уточнения (подтверждение диагноза или отклонение при ложно-положительной реакции на первом этапе). Для этого используются точные хроматографические методы определения ферментов, аминокислот и т.п.
Применяют также микробиологические тесты, они основаны на том, что некоторые штаммы бактерий могут расти только на средах, содержащих определенные аминокислоты, углеводы.
Популяционно-статистический метод
Этот метод позволяет изучить распространение отдельных генов в человеческих популяциях. Обычно производится непосредственное выборочное исследование части популяции либо изучают архивы больниц, родильных домов, а также проводят опрос путем анкетирования. Выбор способа зависит от цели исследования. Последний этап состоит в статистическом анализе. Одним из наиболее простых и универсальных математических методов является метод, предложенный Г.Харди и В. Вайнбергом (в данной статье не рассмотрен). Имеется и ряд других специальных математических методов. В результате становится возможным определить частоту генов в различных группах населения, частоту гетерозиготных носителей ряда наследственных аномалий и болезней.
Изучение распространенности генов на определенных территориях показывает, что в этом отношении их можно разделить на две категории:
•имеющие универсальное распространение (к их числу относится большинство известных генов)
•встречающиеся локально, приемущественно в определенных районах; к их числу относятся, например, ген серповидноклеточной анемии и ген, определяющий врожденный вывих бедра
Популяционно-статистический метод позволяет определить генетическую структуру популяций (соотношение между частотой гомозигот и гетерозигот). Знание генетического состава популяций имеет большое значение для социальной гигиены и профилактической медицины.
Цитогенетический метод
Принципы цитогенетических исследований сформировались в течение 20-30-х годов на классическом объекте генетики – дрозофиле и на некоторых растениях. метод основан на микроскопическом исследовании хромосом.
Для идентификации хромосом применяют количественный морфометрический анализ. С этой целью проводят измерение длины хромосомы в микрометрах (микроскопия хромосом производится в остановленной фазе митоза посредством колхицина и отброшенными посредством гипотонического раствора в результате чего хромосомы лежат свободно), определяют также соотношение длины короткого плеча к длине всей хромосомы (центромерный индекс).
В 1960 году была разработана первая классификация хромосом человека (Денверская). в основу ее были положены особенности величины хромосом и расположение первичной перетяжки. По форме и общим размерам все аутосомы человека подразделяются на 7 групп, обозначаемых латинскими буквами: A, B, C, D, E, F, G. Все хромосомы имеют порядковые номера. Наиболее крупная пара гомологичных хромосом имеет №1, следующая - №2 и т.д. Половые хромосомы - крупная X и мелкаяY – выделяются отдельно. В последнее время разрабатываются автоматические системы для измерения и количественного анализа хромосом. Однако идентификация хромосом только по указанным признакам встречает большие затруднения.
В 1968-1970 гг. были опубликованы работы шведского генетика Касперссона, который применил для изучения хромосом флюоресцентные красители, в частности акрихин-иприт и его производные. Последующее изучение в люминесцентном микроскопе показало, что хромосомы не дают равномерного свечения по длине. В ней выявляется несколько светящихся полос, совпадающих с локализацией структурного гетерохромтина. После удаления их хромосом ДНК они теряют почти полностью способность к флюоресценции.
Если после денатурации ДНК, вызванной нагреванием и некоторыми другими факторами, провести затем ее ренатурацию – восстановление исходной двунитчатой структуры, а затем окрасить хромосомы красителем Гимзы, то в них выявляется четкая дифференцировка на темноокрашенные и светлые полосы – диски. Последовательность расположения этих дисков, их рисунок – строго специфичен для каждой хромосомы. В результате различных вариантов метода удается выявить центромерный и околоцентромерный гетерохроматин (С-диски), диски расположенные по длине хромосом (соответственно Гимзы-диски, G-диски).
Захаровым был разработан перспективный метод изучения хромосом. В основу его положен процесс неодновременной репликации хромосом: одни участки реплицируются раньше, у других этот процесс задерживается и репликация происходит значительно позднее. Неодновременно идет процесс спирализации хромосом, вступающих в митоз. Однако, к тому моменту, когда хромосомы вступают в метафазу, успевет завершиться процесс выравнивания этих различий, и степень конденсации метафазных хромосом становится одинаковой. Было показано, сто можно задерджать этот процесс путем введения 5-бромдезоксиуридина (5-БДУ), который является аналогом тимидина – предшественника ДНК. Если 5-БДУ вводить в конце S-периода. то он включается в синтез ДНК, то есть участки хромосом, где находится это вещество, остаются слабоокрашенными, так как была задержана спирализация. Рано редуплицировавшиеся участки хромосомы, успевшие спирализоваться, интенсивно окрашиваются (Р-диски). Расположение темных и светлых дисков при этом методе противоположно тому, что наблюдается при G-окраске.
Сравнительный анализ различных методов окраски показал, что один и тот же диск может выделяться как светлый неокрашенный или темноокрашенный, но порядок расположения дисков идентичен при и всех методиках. Следовательно, не вызывает сомнения, что их расположение и последовательность имеют закономерный характер. специфичный для каждой хромосомы.
Если нарушения касаются половых хромосом, то методика упрощается. В этом случае проводится не полное кариотипирование, а применяется метод исследования полового хроматина в соматических клетках.
Половой хроматин – это небольшое дисковидное тельце, интенсивно окрашивающееся гематоксилином и другими основными красителями. Они обнаруживаются в интерфазных клеточных ядрах млекопитающих и человека. непосредственно под ядерной мембраной.
Определение полового хроматина нашло применение в судебной медицине, когда требуется по пятнам крови установить половую принадлежность, при анализе. когда надо установить, мужчине или женщине принадлежит найденная часть трупа, даже спустя довольно большой срок после смерти.
При трансплантации тканей тельце полового хроматина может служить своеобразной меткой (если донор и реципиент разных полов). Анализ дает возможность проследить приживление или рассасывание трансплантата.
Методы гибридизации соматических клеток
Соматические клетки содержат весь объем генетической информации. Это дает возможность изучать многие вопросы генетики человека, которые невозможно исследовать на целом организме. Благодаря методам генетики соматических клеток человек как бы стал одним из экспериментальных объектов. Чаще всего используют клетки соединительной ткани (фибробласты) и лимфоциты крови. культивирование клеток вне организма позволяет получить достаточное количество материала для исследования. что не всегда возможно взять у человека без ущерба для здоровья.
Находящиеся в культуре клетки какой-либо ткани можно подвергать изучению различными методами: цитологическим, биохимическим, иммунологическим. такое исследование может быть в ряде случаев более точным, чем на уровне целостного организма, так как метаболические процессы удается выделить из сложной цепи взаимосвязанных реакций. происходящих в организме.
В 1960 г. французский биолог Ж. Барский, выращивая вне организма в культуре ткани клетки двух линий мышей, обнаружил, что некоторые клетки по своим морфологическим и биохимическим признакам были промежуточными между исходными родительскими клетками. Эти клетки оказались гибридными. Такое спонтанное слияние клеток в культуре ткани происходит довольно редко. В дальнейшем оказалось, что частота гибридизации соматических клеток повышается при введении в культуру клеток РНК-содержащего вируса парагриппа Сендай, который как вообще все вирусы, изменяет свойства клеточных мембран и делает возможным слияние клеток. Под влиянием такого вируса в смешанной культуре двух типов клеток образуются клетки, содержащие в общей цитоплазме ядра обеих родительских клеток – гетерокарионы. После митоза и последующего разделения цитоплазмы из двуядерного гетерокариона образуются две одноядерные клетки, каждая из которых представляет собой синкарион – настоящую гибридную клетку, имеющую хромосомы обеих родительских клеток.
В зависимости от целей анализа исследование проводят на гетерокарионах или синкарионах. Синкарионы обычно удается получить при гибридизации в пределах класса. Это истинные гибридные клетки, так как в них произошло объединение двух геномов. Применение метода генетики соматических клеток дает возможность изучать механизмы первичного действия генов и взаимодействия генов.
Метод моделирования
Теоретическую основу биологического моделирования в генетике дает закон гомологических рядов наследственной изменчевости, открытый Н.И. Вавиловым, согласно которому генетически близкие виды и роды характерезуются сходными рядами наследственной изменчивости. Исходя из этого закона, можно предвидеть, что в переделах класса млекопитающих (и даже за его пределами), можно обнаружить многие мутации, вызывающие такие же изменения фенотипических признаков, как и у человека. для моделирования определенных наследственных аномалий человека подбирают и изучают мутантные линии животных, имеющих сходные нарушения.
Были и описаны и изучены многие генные мутации у животных, имеющих сходство с соответствующими наследственными аномалиями человека. Гемофилия А и В встречается у собак, и обусловлена как и у человека, рецессивными генами, локализованными на Х-хромосоме. У хомяков и крыс обнаружены патологические мутации, проявляющиеся как гемофилия, сахарный диабет, ахондроплазия, мышечная дистрофия и некоторые другие. Эпилептоидные припадки встречаются у некоторых кроликов, крыс под влиянием сильного звукового раздражителя.
Мутантные линии животных путем возвратного скрещивания переведены в генетически близкие, в результате получены линии, различающиеся только по аллелям одного локуса. Это дает возможность уточнить механизм развития данной аномалии. Мутантные линии животных не являются точным воспроизведением наследственных болезней человека. Однако, даже частичное моделирование, то есть воспроизведение не всего заболевания в целом, а только патологического процесса или даже его фрагмента, позволяет в ряде случаев обнаружить механизмы первичного отклонения от нормы.