

Механизм действия гормонов через рецепторы, обладающие ферментативной активностью
Гормоны, у которых рецепторы находятся на поверхности клетки, в нее не проникают, а действуют через вторых посредников.
Второй посредник - цГМФ.
Рецептор состоит из двух частей:
наружная - для соединения с гормоном
цитоплазматическая - обладает ферментативной активностью.

Рецептор пронизывает мембрану. Внутренняя часть обладает гуанилатциклазной активностью.
При присоединении гормона меняется конформация ферментативной части, и гуанилатциклаза (ГЦ) активируется. Она превращает сотни ГТФ в цГМФ (усиление сигнала).
В цитоплазме в неактивном виде находится протеинкиназа G, которая в неактивном виде состоит из 2 субъединиц, соединенных вместе в неактивном виде.
Присоединяя к себе 4 молекулы цГМФ, протеинкиназа G активируется.
Она активирует имеющиеся ферменты путем фосфорилирования или дефосфорилирования за счет концевой группы АТФ.
В некоторых клетках находится цитоплазматическая ГЦ, которая имеет в своем составе гем.
Она может активироваться оксидом азота (NO) эндотелия кровеносных сосудов и обладает расслабляющим эффектом.
NO взаимодействует с железом гема. В результате ГЦ активируется и превращает ГТФ в сотни цГМФ.
Вопрос №143.
Ионы кальция как вторые посредники. Каскад ионов Са2+.
Са2+ как второй посредник
Гормоны, у которых рецепторы находятся на поверхности клетки, в нее не проникают, а действуют через вторых посредников.
И спользование
Са2+
как
второго посредника возможно благодаря
тому, что в цитоплазме может повышаться
его концентрация.
спользование
Са2+
как
второго посредника возможно благодаря
тому, что в цитоплазме может повышаться
его концентрация.
В цитоплазме концентрация Са2+ в норме 10-7 моль, а снаружи – 10-5. Под воздействием каких-либо факторов (химических – гормоны, или электрических – потенциал-зависимые Са2+-каналы) в мембранах открываются специальные Са2+-каналы, через которые Са2+ поступает в цитоплазму. Следовательно, его концентрация в цитоплазме повышается.
В цитоплазме Са2+ оказывает несколько эффектов: может связываться со специфическим белком кальмодулином (Ка) (связывающим 4 Са2+), который может менять свою конформацию и…
Активировать Са2+-кальмодулин-зависимую протеинкиназу (Са2+/КаПК).
Са2+/КаПК может активировать ферменты путем фосфорилирования или дефосфорилирования за счет концевой группы АТФ.
Са2+ может связываться с другими цитоплазматическими белками-ферментами (тропонин, пируваткиназа, амилаза).
Небольшое количество Са2+ связывается со специфическими цитоплазматическими белками для депонирования.
Вопрос №144
Инозитол-3-фосфат как вторичный посредник.
Механизм действия через липиды мембран
Гормоны, у которых рецепторы находятся на поверхности клетки, в нее не проникают, а действуют через вторых посредников.
Участвуют два посредника - ИФ3 (инозитолтрифосфат) и ДАГ (дтацилглицерол), которые образуются из фосфолипидов мембран (ФИФ2 - фосфатидилинозитол-4,5-бифосфат).
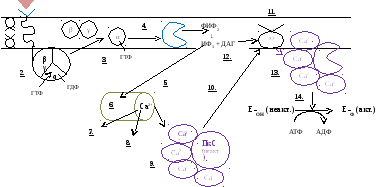
Вторые посредники образуются через 3 встроенных в мембрану белков:
белок-рецептор
G-белок
фосфолипаза С
Гормон соединяется с рецептором менее 1 с и называется первичным вестником сигнала. Образуется гормон-рецепторный комплекс, при этом меняется конформация интегральной части рецептора, и эти изменения передаются на G-белок.
У G-белка отщепляется ГДФ и присоединяется ГТФ. В результате G-белок активируется, сродство α-ГТФ к β и γ ослабевает, она отделяется от них.
α-ГТФ перемещается по мембране (латеральная диффузия), активирует фосфолипазу С, которая расщепляет ФИФ2 на ИФ3 и ДАГ.
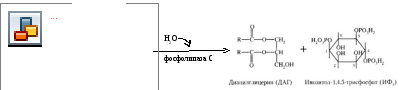
ИФ3 гидрофилен и выходит из мембраны в цитоплазму.
ИФ3 стимулирует выход Са2+ из внутриклеточных депо.
Са2+ может взаимодействовать с кальмодулином (см. кальмодулиновый каскад).
Са2+ может взаимодействовать с цитоплазматическими белками.
Са2+ может активировать протеинкиназу С, которая в неактивном виде находится в цитоплазме.
Протеинкиназа С соединяется с Са2+ и подходит к мембране в место,...
где находится фосфолипид фосфатидилсерин.
Этот фосфолипид узнает второй посредник ДАГ и соединяется с ним.
В этом комплексе протеинкиназа С активируется.
Она активирует ферменты путем фосфорилирования за счет АТФ.