

Обмен нуклеотидов Вопрос №102
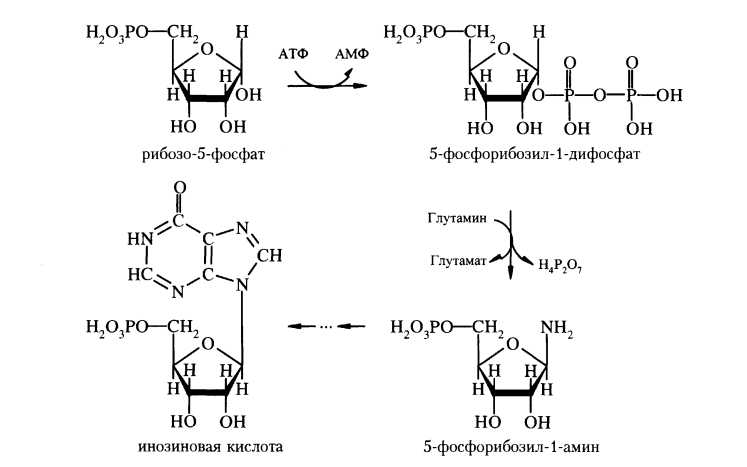
Синтез пуриновых нуклеотидов. Регуляторные реакции.
З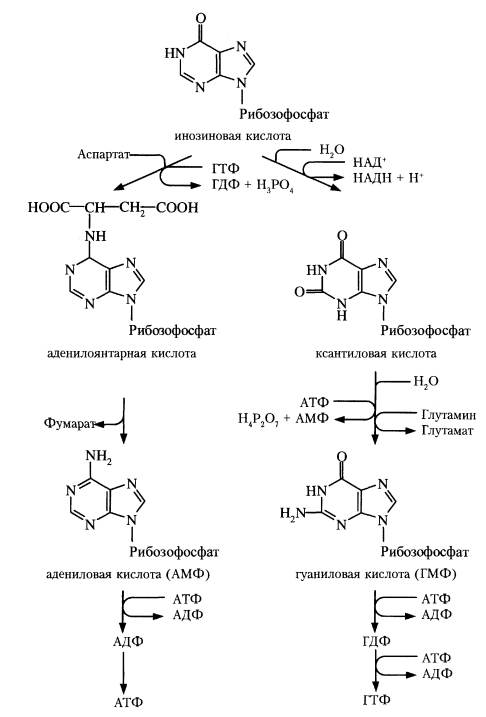
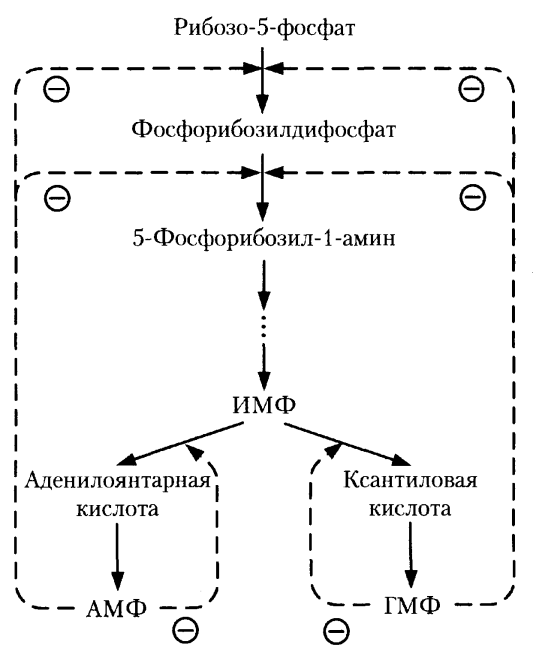 начение
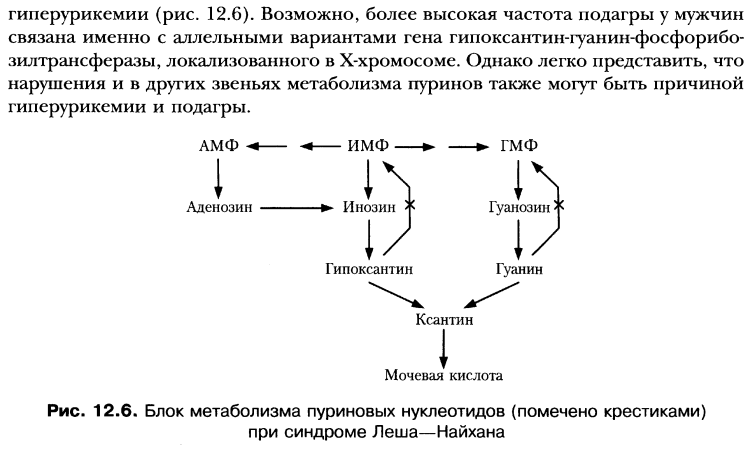
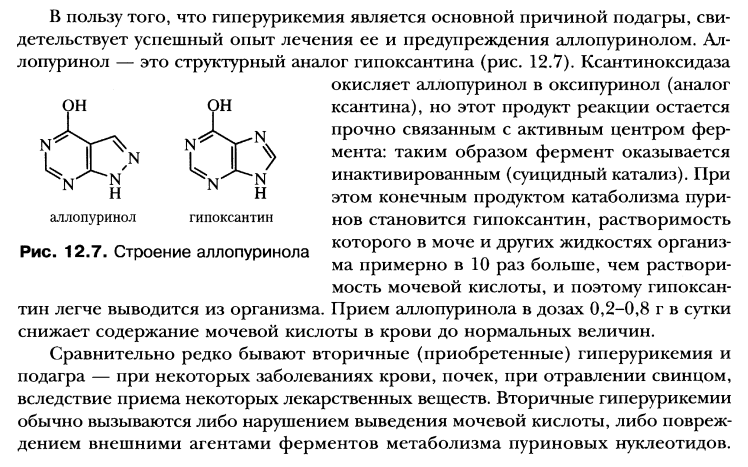
"путей спасения". Синдром Леша-Нихана.
начение
"путей спасения". Синдром Леша-Нихана.
Не синтезируются в лейкоцитах и
эритроцитах.


Вопрос №103
К атаболизм
пуриновых нуклеотидов. Нормальная
концентрация мочевой кислоты в крови.
Подагра: причины возникновения, лечение.
атаболизм
пуриновых нуклеотидов. Нормальная
концентрация мочевой кислоты в крови.
Подагра: причины возникновения, лечение.
Н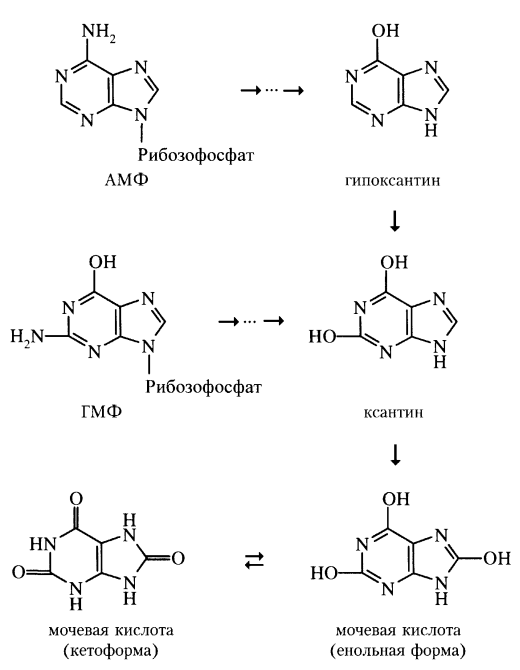 ормальная
концентрация мочевой кислоты: 0,15-0,47
ммоль/л.
ормальная
концентрация мочевой кислоты: 0,15-0,47
ммоль/л.
Причины
подагры
Характер
питания пациента (в меню преобладали
мясные продукты, рыба, алкогольные
напитки, растительные жиры, высококалорийные
блюда).
Провоцирующие
факторы для повторного приступа
(излишние физические нагрузки, факт
переохлаждения или перегревания
суставов)
Наследственность
(больше склонны мужчины).
Ожирение.
Злоупотребление
алкоголем (Алкоголь затрудняет выведение
солей почками. Отсюда появляются
солевые кристаллы, которые и откладываются
на суставах).
Вопрос №104
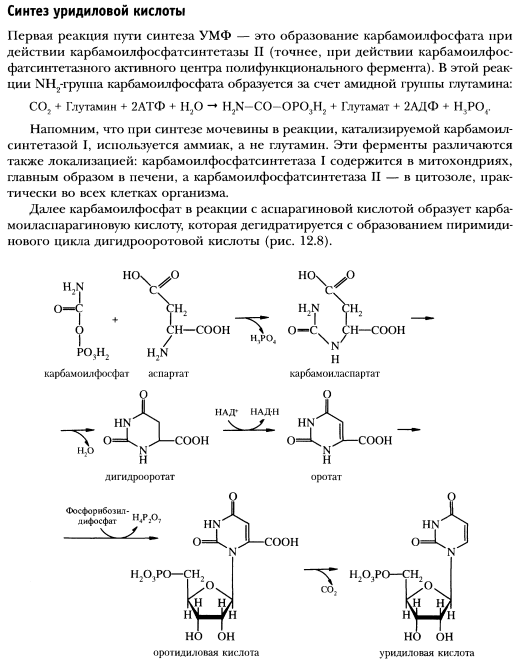
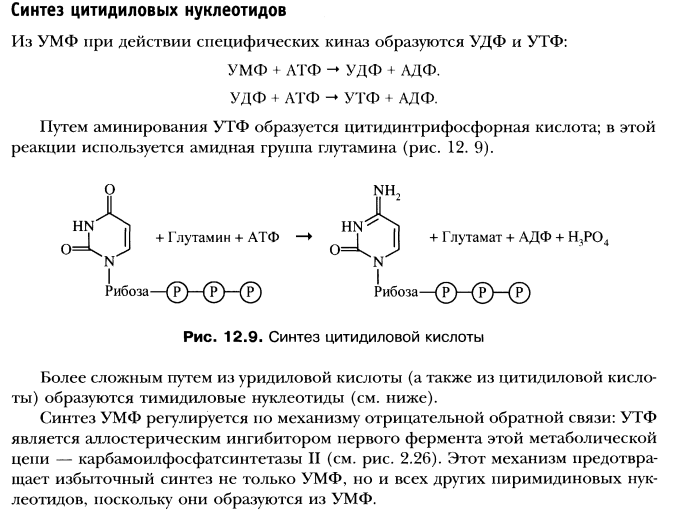
Катаболизм и синтез пиримидиновых нуклеотидов. Регуляция.



Вопрос №105
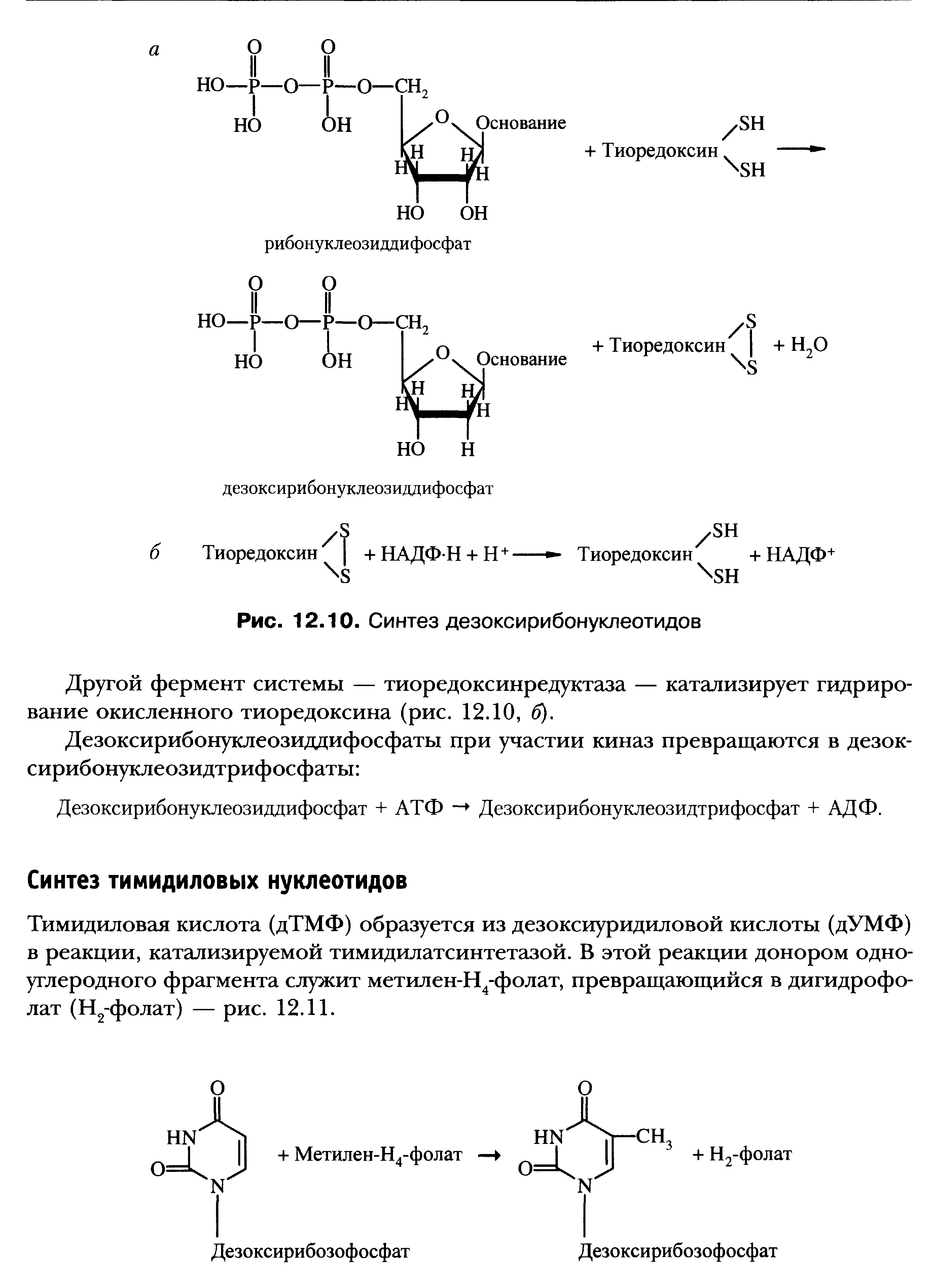
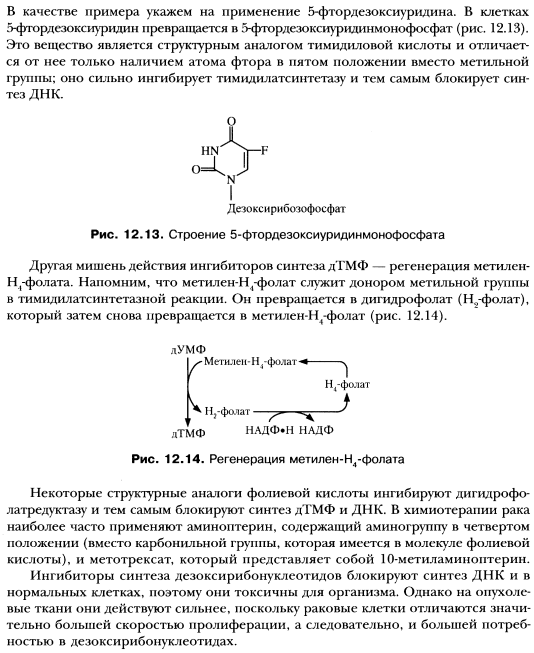
Синтез дезоксирибонуклеотидов с помощью рибонуклеотидредуктазного комплекса. Регуляция. Роль Н4-фолата в образовании дТМФ.
Ингибиторы тимидилсинтетазы и дигидрофолатредуктазы.


Вопрос №106
Изменение гормонального статуса и метаболизма при голодании
1. В постабсорбтивном периоде и голодании уровень глюкозы в плазме крови падает до нижней границы нормы. Отношение инсулин - глюкагон снижается. В этих условиях возникает состояние, для которого характерно преобладание процессов катаболизма жиров, гликогена и белков на фоне общего снижения скорости метаболизма . Под влиянием контринсулярных гормонов в этот период происходит обмен субстратами между печенью, жировой тканью, мышцами и мозгом. Этот обмен служит двум целям:
• поддержанию концентрации глюкозы в крови за счет глюконеогенеза для обеспечения глюкозозависимых тканей (мозга, эритроцитов);
• мобилизации других «топливных» молекул, в первую очередь жиров, для обеспечения энергией всех других тканей.
Проявление этих изменений позволяет условно выделить три фазы голодания . Вследствие переключения метаболизма на режим мобилизации энергоносителей даже после 5-6 недель голодания концентрация глюкозы в крови составляет не менее 65 мг/дл. Основные изменения при голодании происходят в печени, жировой ткани и мышцах.
2. Фазы голодания . Голодание может быть кратковременным - в течение суток (первая фаза), продолжаться в течение недели (вторая фаза) или нескольких недель (третья фаза).
В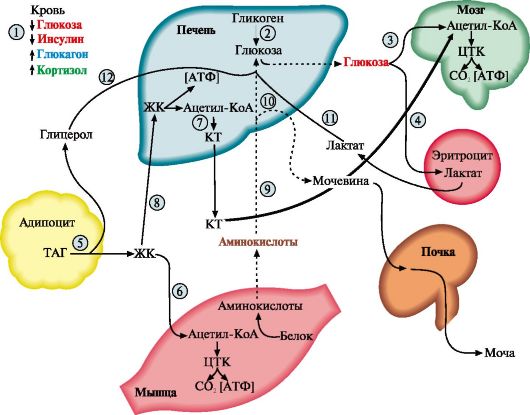 первую фазу концентрация инсулина в
крови снижается примерно в 10-15 раз по
сравнению с периодом пищеварения, а
концентрация глюкагона и кортизола
увеличивается. Запасы гликогена
исчерпываются, нарастает скорость
мобилизации жиров и скорость
глюконеогенеза из аминокислот и
глицерола, концентрация глюкозы в крови
снижается до нижней границы нормы (60
мг/дл).
первую фазу концентрация инсулина в
крови снижается примерно в 10-15 раз по
сравнению с периодом пищеварения, а
концентрация глюкагона и кортизола
увеличивается. Запасы гликогена
исчерпываются, нарастает скорость
мобилизации жиров и скорость
глюконеогенеза из аминокислот и
глицерола, концентрация глюкозы в крови
снижается до нижней границы нормы (60
мг/дл).
1 - снижение инсулин-глюкогонового индекса; 2 - мобилизация гликогена; 3, 4 - транспорт ГЛК в мозг и эритроциты; 5 - мобилизация ТАГ; 6 - транспорт ЖК в мышцы; 7 - синтез кетоновых тел; 8 - транспорт ЖК в печени; 9 - транспорт АК в печень; 10 - глюконеогенез из АК; 11 - транспорт лактата в печень; 12 - транспорт глицерола в печень. Пунктиром обозначены процессы, скорость которых снижается
Во вторую фазу продолжается мобилизация жиров, концентрация жирных кислот в крови повышается, увеличивается скорость образования кетоновых тел в печени и , соответственно, их концентрация в крови; ощущается запах ацетона, который выделяется с выдыхаемым воздухом и потом от голодающего человека. Глюконеогенез продолжается за счет распада тканевых белков.
В третью фазу снижается скорость распада белков и скорость глюконеогенеза из аминокислот. Скорость метаболизма замедляется. Азотистый баланс во все фазы голодания отрицательный. Для мозга важным источником энергии, наряду с глюкозой, становятся кетоновые тела.
3. Изменения метаболизма основных энергоносителей при голодании . Обмен углеводов. Запасы гликогена в организме истощаются в течение 24-часового голодания. Таким образом, за счет мобилизации гликогена обеспечивается только кратковременное голодание. Основной процесс, обеспечивающий ткани глюкозой в период голодания, - глюконеогенез. Глюконеогенез начинает ускоряться через 4-6 часов после последнего приема пищи и становится единственным источником глюкозы в период длительного голодания. Основные субстраты глюконеогенеза - аминокислоты, глицерол и лактат.
4. Обмен жиров и кетоновых тел. Основным источником энергии в первые дни голодания становятся жирные кислоты, которые образуются из ТАГ в жировой ткани. В печени ускоряется синтез кетоновых тел. Синтез кетоновых тел начинается в первые дни голодания. Используются кетоновые тела в основном в мышцах. Энергетические потребности мозга частично обеспечиваются также кетоновыми телами. После 3 недель голодания в мышцах снижается скорость окисления кетоновых тел и мышцы почти исключительно используют жирные кислоты. Концентрация кетоновых тел в крови возрастает. Использование кетоновых тел мозгом продолжается, но становится менее активным из-за снижения скорости глюконеогенеза и снижения концентрации глюкозы.
5. Обмен белков. В течение нескольких первых дней голодания быстро распадаются мышечные белки - основной источник субстратов для глюконеогенеза. После нескольких недель голодания скорость глюконеогенеза из аминокислот снижается в основном вследствие снижения потребления глюкозы и использования кетоновых тел в мозге. Снижение скорости глюконеогенеза из аминокислот необходимо для сбережения белков, так как потеря 1/3 всех белков может привести к смерти. Продолжительность голодания зависит от того, как долго могут синтезироваться и использоваться кетоновые тела. Однако для окисления кетоновых тел необходим оксалоацетат и другие компоненты ЦТК. В норме они образуется из глюкозы и аминокислот, а при голодании только из аминокислот.