

1. Тиролиберин (Піро-Глу-Гіс-Про-nh 2):
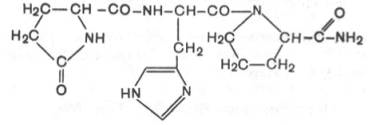
Тиролиберин представлений трипептидом, що складається з піроглутамінової (циклічної) кислоти, гістидину і пролінаміда, з'єднаних пептидними зв'язками. На відміну від класичних пептидів він не містить вільних NH2-і СООН-груп у N-і С-кінцевих амінокислот.
Тиреотропін-рилізинг-гормон(ТРГ) - один з представників класу рилізинг-гормонів гіпоталамуса. Існує також аналогічний гормон епіфіза.
ТРГ викликає посилення секреції передньою долею гіпофіза тиреотропного гормону, а також, в меншій мірі, посилення секреції пролактину.
ТРГ також є нейропептид, які приймає участь в регуляції деяких психічних функцій. Зокрема, встановлено наявність антидепресивної дії екзогенного ТРГ при депресіях, незалежного від збільшення секреції тиреоїдних гормонів, які також володіють деякою антидепресивною активністю.
Супутнє підвищення секреції пролактину під дією ТРГ є однією з причин, який нерідко спостерігається при первинному гіпотиреозі (при якому підвищений рівень ТРГ внаслідок зменшення переважної дії тиреоїдних гормонів на тиреотропну функцію гіпоталамуса) гіперпролактинемії. Іноді гіперпролактинемія при цьому буває настільки значною, що призводить до розвитку гінекомастії, галактореї та імпотенції у чоловіків, галактореї або патологічно рясною і тривалою фізіологічної лактації у жінок, мастопатій, аменореї.
Синтез. ТРГ виробляється в гіпоталамусі у нейросекреторних клітинах. Спочатку він синтезується як 242-кислотний амінопласти, попередник поліпептиду, що містить 6 копій послідовності Glu-His-Pro-Gly, обрамлює двохосновні пептидами, який пізніше розщеплюється, щоб отримати готову молекулу ТРГ. Вона проходить через серединний горбок в попередню гіпофізну залозу через гіпофізальну портальну систему, де він стимулює клітини, звані териотропами, які відповідальні за випуск гормону, що стимулює роботу щитовидної залози. ТРГ також може виявитися не тільки в мозку, а й в інших частинах тіла, включаючи шлунково-кишковий тракт і панкреатичні острівці.
Історія. Послідовність ТРГ було вперше визначено і синтезована Роджером Гуіллеміном і Ендрю В. Шеллі в 1969 році.
Клінічне значення. Використовується у фармакології для того, щоб перевірити відповідь попередньої гіпофізарної залозі. Медичні препарати на основі ТРГ використовуються в діагностиці порушень роботи щитовидної залози і акромегалії.
2. Гонадоліберину є декапептид, що складається з 10 амінокислот в послідовності:
Пиро-Глу–Гис–Трп–Сер–Тир–Гли–Лей–Арг–Про–Гли-NН2
Кінцева С-амінокислота представлена гліцинамідом.
Гонадотропін-рилізинг-гормон (ГнРГ) - один з представників класу рилізинг-гормонів гіпоталамуса. Існує також аналогічний гормон епіфіза.
ГнРГ викликає посилення секреції передньою долею гіпофіза гонадотропних гормонів - лютеїнізуючого гормону і фолікулостимулюючого гормону. При цьому ГнРГ більшою мірою впливає на секрецію лютеїнізуючого, ніж фолікулостимулюючого гормону, за що і називається люліберин або лютрелин.
Гонадотропін-рилізинг-гормон за будовою є поліпептидний гормон.
Секреція ГнРГ відбувається не постійно, а у вигляді коротких піків, наступних один за одним з чітко визначеними часовими інтервалами. При цьому інтервали ці різні в чоловіків і у жінок: у нормі у жінок викиди ГнРГ слідують кожні 15 хв у фолікулярній фазі циклу і кожні 45 хв в лютеїновій фазі і під час вагітності, а у чоловіків - кожні 90 хв.
Введення екзогенного ГнРГ в режимі постійної краплинної інфузії або введення довготривалих синтетичних аналогів ГнРГ викликає короткочасне збільшення секреції гонадотропних гормонів, швидко змінюються глибоким пригніченням і навіть вимиканням гонадотропної функції гіпофіза і функції статевих залоз внаслідок десенсибілізації рецепторів ГнРГ гіпофіза.
У той же час введення екзогенного ГнРГ за допомогою спеціальної помпи, що імітує природний ритм пульсації секреції ГнРГ, забезпечує тривалу та стійку стимуляцію гонадотропної функції гіпофіза, причому правильний режим помпи забезпечує правильне співвідношення ЛГ і ФСГ по фазах циклу у жінок і правильне, характерне для чоловіків, співвідношення ЛГ і ФСГ у чоловіків.
Механізм дії. ГнРГ стимулює передню частку гіпофіза, клітини-гонадотропи, в мембранах яких знаходяться ГнРГ-рецептори, до секреції двох гормонів: фолікулостимулюючий гормон (ФСГ) і лютеїнізуючий гомін (ЛГ). Ці гормони також об'єднують під загальною назвою гонадотропінів. Гонадотропін - це гормон, що стимулює активність гонад, в даному випадку насінників. ФСГ стимулює сперматогенез, внаслідок чого клітини Сертоли сприяють завершенню розвитку з сперматид сперміїв. ЛГ спонукає клітини Лейдіга або інтерстиціальні клітини семенника синтезувати гормон тестостерон. Тестостерон стимулює інтерстиціальні клітини чоловіка, він представляє собою стероїдний гормон, що утворюється з холестеролу. Він викликає ріст і розвиток сперміїв з клітин зачаткового епітелію, також разом з ФСГ він надає стимулюючу дію на клітини Сертоли. Механізм, що діє за принципом негативного зворотнього зв'язку, підвищує рівень тестостерону, і це веде до зменшення швидкості секреції ГнРГ гіпоталамусом, що, в свою чергу, веде до зниження рівнів ЛГ і ФСГ. Тестостерон також впливає на передню долю гіпофіза, знижуючи секрецію ЛГ, але це дія виражена слабше. Клітини Сертоли виробляють глікопротеїновий гормон - інгібін. Він регулює утворення сперміїв за принципом негативного зворотного зв'язку. Якщо сперматогенез відбувається занадто швидко, то починає синтезуватися інгібін, який, впливаючи на передню частку гіпофіза, знижує секрецію ФСГ. Інгібін впливає на гіпоталамус, зменшуючи секрецію ГнРГ. Якщо сперматогенез йде повільно, то ФСГ стимулює сперматогенез. ФСГ і ЛГ викликають у клітинах, які вони стимулюють, виділення цАМФ в цитоплазму, а потім у ядро, де стимулюється синтез ферментів.
3. Соматостатин є циклічним тетрадекапептідом (складається з 14 амінокислотних залишків):

Відрізняється цей гормон від двох попередніх, крім циклічної структури, тим, що не містить на N-кінці піроглутамінової кислоти: дисульфідний зв'язок утворюється між двома залишками цистеїну в 3-му і 14-му положеннях. Слід зазначити, що синтетичний лінійний аналог соматостатину також наділений аналогічною біологічною активністю, що свідчить про неістотність дисульфідного містка природного гормону. Крім гіпоталамуса, соматостатин продукується нейронами центральної і периферичної нервових систем, а також синтезується в S-клітинах панкреатичних острівців (острівців Лангерганса) в підшлунковій залозі і клітинах кишечника. Він надає широкий спектр біологічної дії, зокрема, показано інгібуючу дію на синтез гормону росту в аденогипофізе, а також пряму гальмівну дію його на біосинтез інсуліну і глюкагону в β-і α-клітинах острівців Лангерганса.
Соматостатин - гормон дельта-клітин острівців Лангерганса підшлункової залози, а також один з гормонів гіпоталамуса. За хімічною будовою є пептидним гормоном.
Структура. Соматостатин існує у двох біологічно активних формах, що походять від одного попередника і різняться довжиною N-кінця: SST-14 (14 амінокислот: AGCKNFFWKTFTSC, між залишками цистеїну дисульфідний зв'язок) і SST-28 (28 амінокислот). Крім соматостатину, близькоспоріднені пептиди кортистатин (PCKNFFWKTFSSCK), триттіі (13 N-кінцевих амінокислот SST-28) і уротензин II (ETPDCFWKYCV) також беруть участь в регуляції соматостатинергічної системи.
Функція. Соматостатин пригнічує секрецію гіпоталамусом соматотропін-рилізинг-гормону і секрецію передньою долею гіпофіза соматотропного гормону та тиреотропного гормону.
Крім того, він пригнічує також секрецію різних гормонально активних пептидів і серотоніну, що продукуються в шлунку, кишечнику, печінки і підшлункової залози. Зокрема, він знижує секрецію інсуліну, глюкагону, гастрину, холецистокинина, вазоактивного інтестинального пептиду.
Соматостатинові рецептори поділяються на дві групи - SRIF1 (sst2, sst3, sst5) і SRIF2 (sst1, sst4) - на підставі зв'язування з класичними октапептидними і гексапептидними аналогами соматостатину (октреотид, лантреотід, сеглітід, вапреотід): з ними легко зв'язуються SRIF1-, але не SRIF2-рецептори.
Терапевтичний потенціал. Найбільш поширений фармакологічний вплив на соматостатинергічну систему у зв'язку з інгібуванням викиду гормону росту, що робить дану систему досить перспективною при лікуванні пухлинних захворювань і акромегалії. Речовини, що підсилюють викид соматостатину, можуть проявляти властивості когнітивних стимуляторів.
Комерційні препарати. Найбільш поширений і відомий октреотид (і його генерики) - синтетичне похідне гормону соматостатину, що виявляє подібні з ним фармакологічні ефекти і значно більшою тривалістю дії. Пригнічує патологічно підвищену секрецію гормону росту, пептидних гормонів і серотоніну, що продуктуються в гастроентеропанкреатичнйі ендокринній системі. Введення октреотиду не супроводжується феноменом гіперсекреції гормонів за механізмом «негативного зворотного зв'язку».
У РФ соматостатин проводиться у формі розчину для ін'єкцій 0,01% і 0,005%, а так само у формі ліофілізату для приготування суспензії для внутрішньом'язового введення пролонгованої дії (октреотид-депо).
Крім перерахованих гіпоталамічних гормонів, інтенсивно вивчалася хімічна природа іншого гормону - кортиколиберина. Активні препарати були виділені як з тканини гіпоталамуса, так і з задньої долі гіпофіза, існує думка, що остання може служити депо гормону вазопресину для і окситоцину. Місцем синтезу гіпоталамічних гормонів, наймовірніше, є нервові закінчення - синаптосами гіпоталамуса, оскільки саме там відзначена найбільша концентрація гормонів і біогенних амінів. Останні розглядаються поряд з гормонами периферичних залоз внутрішньої секреції, що діють за принципом зворотного зв'язку, в якості основних регуляторів секреції та синтезу гормонів гіпоталамуса. Механізм біосинтезу тиролиберина, що здійснюється, швидше за все, включає участь SH-яка містить синтетази або комплексу ферментів, які каталізують циклізації глутамінової кислоти в піроглутамінову. Існування такого механізму біосинтезу за участю відповідних синтетаз допускається також відносно гонадолиберину і соматолиберину.
У гіпоталамусі людини і ряду тварин відкритий специфічний фермент піроглутамілпептидаза, яка каталізує відщеплення від тиролиберина або гонадолиберину молекули піроглутамінової кислоти.
Гіпоталамічні гормони безпосередньо впливають на секрецію (точніше, звільнення) «готових» гормонів і біосинтез цих гормонів заново. Доведено, що цАМФ бере участь у передачі гормонального сигналу. Показано існування в плазматичних мембранах клітин гіпофіза специфічних аденогіпофізних рецепторів, з якими зв'язуються гормони гіпоталамуса, після чого через систему аденилатциклази і мембранних комплексів Са2 +-АТФ і Mg2 +-АТФ звільняються іони Са2 + і цАМФ, останній діє як на звільнення, так і на синтез відповідного гормону гіпофіза шляхом активації протеїнкінази.
Для з'ясування механізму дії рилізинг-факторів, включаючи їх взаємодію з відповідними рецепторами, велику роль зіграли структурні аналоги тиролиберина і гонадолиберину. Деякі з цих аналогів володіють навіть більш високою гормональною активністю і пролонговану дію, ніж природні гормони гіпоталамуса.