

-
Їх утворення.
Майже всі біологічно активні пептиди, у тому числі пептидні гормони, синтезуються в організмі у вигляді білкових попередників, з яких вони утворюються в результаті розпаду пептидних зв’язків під дією ферментів.
Пептиди виконують важливу функцію в організмі. Гіпоталамічний нейрон - гормони регулюють діяльність гіпофіза, що контролює функції всіх інших залоз внутрішньої секреції. Пептиди, що володіють морфино подібною дією (ендогенні опіати) впливають на механізми сприйняття болючих відчуттів. Вазопресин, окситоцин, кортикотропін впливають на кровоносні судини, скорочення м’язів матки, а також на поведінку, пам’яті, мотивацію й навчання.
Основна функція біологічно активних пептидів полягає в тому, що вони переносять інформацію про те, як правильно повинна працювати клітина. З розвитком нанотехнологій стало можливо використовувати зверх малі пептиди для того, щоб контролювати й відновлювати роботу клітин. Сьогодні пептиди найчастіше застосовують у геронтології й косметології. Але вже не за горами час, коли будуть розроблені лікарські препарати на основі пептидів для лікування самих різних захворювань.
Розділ 2. Біологічно активні пептиди.
У цьому розділі не вистачає посилань на джерела інформації. Інформація повинна бути перевірена, інакше вона може бути поставлена під сумнів і вилучена. Пептиди, володіючи високою фізіологічною активністю, регулюють різні біологічні процеси. За своїми біорегуляторними діями пептиди прийнято ділити на кілька груп:
-
Сполуки, що володіють гормональною активністю (глюкагон, вазопрессин, кальцитонін і ін.) і гормони Гіпоталамуса;
Глюкагон.
Глюкагон
– це поліпептид, який складається із
29 амінокислотних залишків. Глюкагон
вперше був виявлений в комерційних
препаратах інсуліну ще в 1923р., Проте
тільки в 1953р., угорський
біохімік Штрауб отримав цей гормон в
гомогенному стані. Синтезується
з білка-попередника
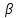 - клітинах підшлункової залози:
препроглюкагон – проглюкагон –
глюкагон.У глюкагону, на відміну від
інсуліну, відсутні пролін, ізолейцин і
цистин, зате є залишки триптофану і
метіоніну, яких немає в інсуліні.
- клітинах підшлункової залози:
препроглюкагон – проглюкагон –
глюкагон.У глюкагону, на відміну від
інсуліну, відсутні пролін, ізолейцин і
цистин, зате є залишки триптофану і
метіоніну, яких немає в інсуліні.
H-His-Ser-Gln-Gly-Thr-Phe-Thr-Ser-Asp-Tyr-Ser-Lys-Tyr-Leu-Asp
|
HO-Thr-Asn-Met-Leu-Try-Gln-Val-Phe-Asp-Gln-Ala-Arg-Arg-Ser
Глюкагон
Первинна структура глюкагону людини і тварин виявилася ідентичною.За біологічних дій глюкагон відноситься до гіперглікемічних факторів, причому цей ефект обумовлений головним чином розпадом глікогену. Органами - мішенями для глюкагону є печінка, міокард, жирова тканина, але не скелетні м'язи. Біосинтез і секреція глюкагону контролюються головним чином концентрацією глюкози за принципом зворотного зв'язку; аналогічною властивістю володіють амінокислоти і вільні жирні кислоти. На секрецію глюкагону впливають також інсулін і недавно виділений пептид, названий вазоактивним пептидом, що володіє широким спектром біологічної дії. У механізмі дії глюкагону первинним є зв'язування зі специфічними рецепторами мембрани клітин; утворився глюкагонрецепторний комплекс активує аденілатциклазу і відповідно утворення цАМФ.
Фосфорилювання регуляторних ферментів під дією протеїнкіназ стимулює одні метаболічні процеси і гальмує інші.
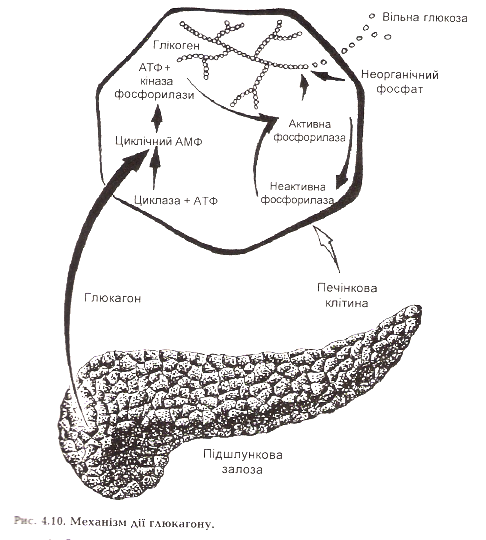
Ефекти глюкагону:
-
Стимулює розщеплення глікогену печінки до вільної глюкози (активація фосфорилази).
-
Пригнічує гліколіз внаслідок гальмування активності фосфофруктокінази, піруваткінази, піруватдегідрогенази.
-
Стимулює розщеплення білків, особливо у мязах, що забезпечує постачання амінокислот для глюконеогенезу.
-
Стимулює глюконеогенез у печінці, що забезпечується надходженням субстратів – амінокислот, гліцерину і активацією ключових ферментів процесу – піруваткарбоксилази, фруктозо-1,6-дифосфатази.
-
Стимулює розщеплення жирів у жировій тканині (активація гормоночутливої ліпази), підвищення рівня жирних кислот у крові і утилізації їх у тканинах.
-
Стимулює утворення кетонових тіл у печінці.
-
Гальмує синтез білків, жирів, фосфоліпідів, холестерину.
-
Збільшує клубочкову фільтрацію.
Останній, будучи універсальним ефекторів внутрішньоклітинних ферментів, активує протеїн, яка в свою чергу фосфорилірує кіназу фосфорілази і глікогенсінтази. Фосфорилювання першого ферменту сприяє формуванню активної глікогенфосфорилазу і відповідно розпаду глікогену до стадії глюкозо - 1 - фосфат, в той час як фосфорилювання глікогенсинтази супроводжується переведенням її в неактивну форму і відповідно до блокування синтезу глікогену. Загальним підсумком дії глюкагону є прискорення розпаду глікогену і гальмування його синтезу, результатом якого є глікогеноліз - розпад глікогену до глюкози в печінці.
Гіперглікемічний ефект глюкагону обумовлений, однак, не тільки глікогенолізу. Є безперечні докази існування глюконеогенетичного механізму гіперглікемії, викликаної глюкагоном. Виявляється, що глюкагон сприяє утворенню глюкози з проміжних продуктів обміну білків і жирів. Зокрема, джерелами глюконеогеназа є амінокислоти, піровиноградна та молочна кислоти та ін. Глюкагон стимулює утворення глюкози з амінокислот шляхом індукції синтезу ферментів глюконеогенезу за участю цАМФ. Слід зазначити, що, крім панкреатичного глюкагону, останнім часом отримані докази існування кишкового глюкагону, який синтезує в кров. Первинна структура кишкового глюкагону поки точно не розшифрована, оскільки він ще не отриманий в чистому вигляді, а проте в кишковому глюкагону відкрита ідентична N-кінцева та середня ділянки панкреатичного глюкагону послідовність амінокислот і різна С-кінцева послідовність амінокислот.
Таким чином, острівці Лангерганса, які синтезують дві протилежні дії гормону - інсулін і глюкагон, виконують ключову роль в регуляції обміну речовин на молекулярному рівні.
Вазопресин. Вазопресин синтезується у тілах нейронів гіпоталамуса, по аксонах переміщається до задньої частини гіпофіза і через нервові закінчення виділяється у кров. За хімічною природою утворюються із більших попередників. Пропресофізин дає вазопресин і білок нейрофізин. Біологічна роль нейрофізинів полягає в не ковалентному зв’язуванні вазопресину й окситоцину та транспорті їх із гіпоталамуса. У нейрогіпофізі комплекси розпадаються і вільні гормони декретуються у кров.
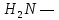 цис—тир—фен—глн—асн—цис—про—арг—глі—C—
цис—тир—фен—глн—асн—цис—про—арг—глі—C—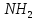
| | ǁ
S————————————S O вазопресин
Відрізняються вони тільки двома амінокислотними залишками (в 3 і 8 положеннях).
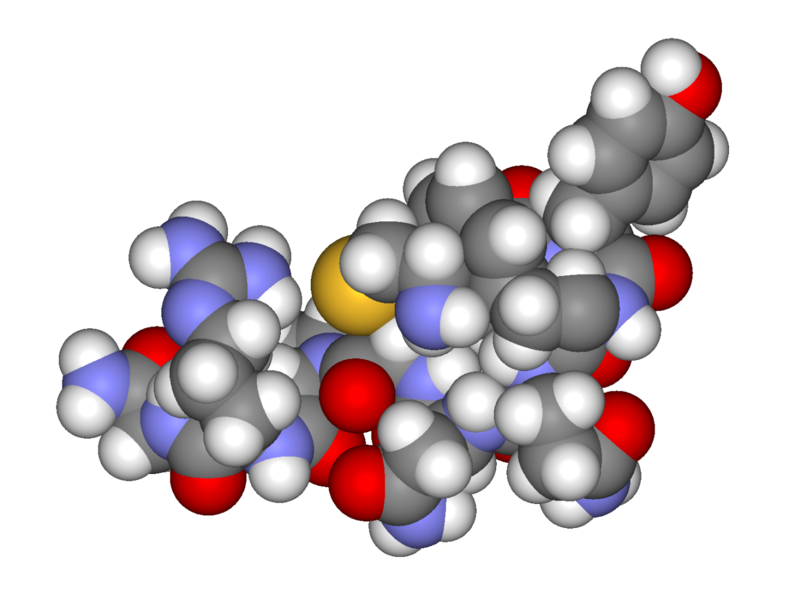
Дія вазопресину характеризуються такими ефектами:
-
Антидіуретична дія. У клітинах ниркових канальців взаємодія АДГ зV2-рецепторами викликає підвищення рівня цАМФ, фосфорилювання поки що невідомих білків, що зумовлює збільшення проникності мембрани для води, і реабсорбцію води, вільної від іонів, за градієнтом концентрації із гіпотонічної первинної сечі через клітини в позаклітинну рідину. В результаті осмотичний тиск плазми крові і тканинної рідини зменшується і секреція гормону припиняється.
-
Підтримка артеріального тиску. Взаємодія АДГ з V1-рецепторами гладком'язових клітин в судинах викликає збільшення концентрації іонів кальцію в клітинах і скорочення мязів, звуження судин, підвищення кровяного тиску. Пресорний ефект вазопресину спостерігається при дії значної кількості гормону.
-
Участь у механізмах памяті. АДГ позитивно діє на закріплення памяті й мобілізацію інформації, що зберігається.
Секреція АДГ регується змінами осмотичного тиску і об’єму циркулюючої крові, а також різними нейрогенними стимулами. Специфічні осморецептори мозку реагують на підвищення осмотичного тиску плазми крові і тканинної рідини сигналами про виділення вазопресину в кров і навпаки. При крововтраті, зниженні об’єму крові барорецептори клітин кровоносних судин передають сигналами в ЦНС і стимулюють секрецію АДГ, а також альдостерону. Вивільнення АДГ гальмується адреналіном.
При недостатності АДГ виникає нецукровий діабет, при якому за добу із організму виводиться 10-20л дуже гіпотонічної сечі. Лікується природним гормоном чи синтетичними аналогами. Відомі препарати з чистою антидіуретичною дією без пресорної активності. Нейрогенний нецукровий діабет зумовлюється втратою здатності рецепторів клітин дистальних відділів нефрону реагувати на АДГ.
Кальцитонін.
Кальцитонін
– пептид із 32 амінокислотних залишків,
синтезується в пара фолікулярних
клітинах щитоподібної залози, які
відкриті також у тканині пар щитоподібних
залоз і тимуса людини. Утворення і
секреція кальцитоніну в кров регулюється
концентрацією іонного кальцію в плазмі.
При нормальній концентрації
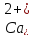 декретуються
невеликі кількості обох гормонів. При
зниженні концентрації
декретуються
невеликі кількості обох гормонів. При
зниженні концентрації
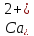 пара
щитоподібних залоз збільшують секрецію
паратгормону, а секреція кальцитоніну
гальмується. При підвищенні концентрації
пара
щитоподібних залоз збільшують секрецію
паратгормону, а секреція кальцитоніну
гальмується. При підвищенні концентрації
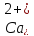 виділяється
кальцитонін, а активність пар щитоподібних
залоз знижується.
виділяється
кальцитонін, а активність пар щитоподібних
залоз знижується.
Кальцитонін
гальмує резорбцію кісткової тканини
остеобластями і остеоцитами, що
супроводжується зниженням вмісту в
плазмі крові кальцію і фосфатів. Дія
кальцитоніну реалізується через
аденілатциклазну систему. При тривалій
дії кальцитоніну зменшується утворення
остеобластів із клітин - попередників,
що вторинно викликає зменшення числа
остеобластів. У нирках збільшується
екскреція фосфатів, напевно, внаслідок
зміни концентрації
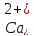 в
плазмі. Таким чином, на виведення фосфатів
кальцитонін діє як синергіст, а на
концентрацію кальцію в крові і на кістки
– як антагоніст.
в
плазмі. Таким чином, на виведення фосфатів
кальцитонін діє як синергіст, а на
концентрацію кальцію в крові і на кістки
– як антагоніст.
Гіпоталамус служить місцем безпосередньої взаємодії вищих відділів центрально нервової системи (ЦНС) та ендокринної системи. Природа зв'язків, що існують між ЦНС і ендокринною системою, стала прояснюватися в останні десятиліття, коли з гіпоталамуса були виділені перші гуморальні фактори, що опинилися гормональними речовинами з надзвичайно високою біологічною активністю. Знадобилося чимало праці та експериментальної майстерності, щоб довести, що ці речовини утворюються в нервових клітинах гіпоталамуса, звідки за системою портальних капілярів досягають гіпофіза і регулюють секрецію гіпофізарних гормонів, точніше їх звільнення (можливо, і біосинтез). Ці речовини одержали спочатку найменування нейрогормонів, а потім рилізинг-факторів (від англ реліз -. Звільняти), або ліберинів. Речовини з протилежною дією, тобто пригнічують звільнення (і, можливо, біосинтез) гіпофізарних гормонів, які стали називати інгібуючими факторами, або статинами. Таким чином, до гормонів гіпоталамуса належить ключова роль у фізіологічній системі гормональної регуляції багатосторонніх біологічних функцій окремих органів, тканин і цілісного організму.
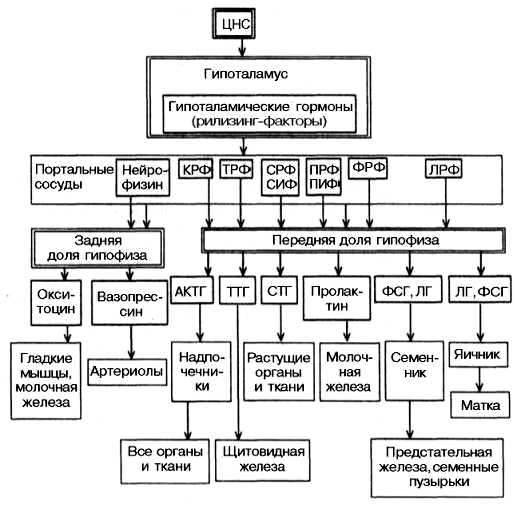
До теперішнього часу в гіпоталамусі відкрито 7 стимуляторів (ліберин) і 3 інгібітора (статини) секреції гормонів гіпофіза, а саме: кортиколіберин, тиролиберин, люлиберин, фоллілиберин, соматолиберин, пролактолиберин, меланолиберин, соматостатин, пролактостатин і меланостатин. У чистому вигляді виділено 5 гормонів, для яких встановлена первинна структура, підтверджена хімічним синтезом.
Великі труднощі при отриманні гормонів гіпоталамуса в чистому вигляді пояснюються надзвичайно низьким вмістом їх у вихідної тканини. Так, для виділення всього 1 мг тиролиберина знадобилося переробити 7т гіпоталамуса, отриманих від 5 млн овець.
Слід зазначити, що не всі гормони гіпоталамуса, мабуть, строго специфічні щодо одного будь-якого гіпофізарного гормону. Зокрема, для тиролиберина показана здатність звільняти, крім тиротропіна, також пролактин, а для люліберина, крім лютенізіруючого гормону, - також фолікулостимулюючий гормон.
Гіпоталамічні гормони не мають твердо встановлених найменувань. Рекомендується в першій частині назви гормону гіпофіза додавати закінчення «ліберинів»; наприклад, «тиролиберин» означає гормон гіпоталамуса, що стимулює звільнення (і, можливо, синтез) тиротропіна - відповідного гормону гіпофіза. Аналогічним чином утворюють назви факторів гіпоталамуса, що інгібують звільнення (і, можливо, синтез) стежинок гормонів гіпофіза, - додають закінчення «статин». Наприклад, «соматостатин» означає гіпоталамічний пептид, що інгібує звільнення (або синтез) гормону росту гіпофізу - соматотропіну.
Встановлено, що за хімічною будовою всі гормони гіпоталамуса є низькомолекулярними пептидами, так званими олігопептидами незвичайної будови, хоча точний амінокислотний склад і первинна структура з'ясовані не для всіх.