

18:1;9 Олеиновая
18:2;9,12 Линоленовая
20:4;5,8,11,14 Арахидоновая
Жирные кислоты, содержащие двойные связи, называют ненасыщенными. Ненасыщенные жирные кислоты представлены моноеновыми (с одной двойной связью) и полиеновыми (с двумя и большим числом двойных связей). Если в составе жирной кислоты содержатся две и более двойных связей, то они располагаются через СН2-группу.
Структурная формула моноеновой (пример,олеиновая)
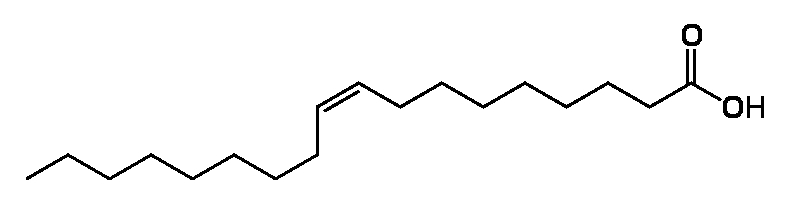
Стр.ф-ла палеиновой (линолевая)
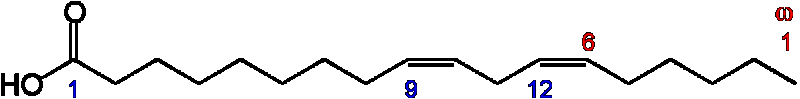
Св-ва,распространение.
НЖК с одной или несколькими двойными связями являются структурными элементами фосфолипидов мембран и имеются в организме человека в значительных количествах (незаменимые жирные кислоты — линолевая, линоленовая, арахидоновая — поступают в организм с пищей). Самой распространенной из ненасыщенных жирных кислот является олеиновая. В фосфолипидах животных тканей очень мало содержится линолевой кислоты (0,05-0,4%), так как она превращается в линоленовую и арахидоновую. Линоленовая НЖК содержится в значительных количествах — 4-24%, содержание арахидоновой кислоты в фосфолипидах тканей составляет 0,2-22% (табл. 1). Биологическое значение ненасыщенных жирных кислот в метаболизме окончательно не выяснено, механизмы катаболизма НЖК в клетках животных также досконально не изучены. В молекулах НЖК две двойные связи, расположенные следующим образом: -СН=СН-СН=СН–, называются сопряженными (конъюгированными) (Л. Уайт и соавт., 1981).
Двойные связи определяют существование двух разных жирных кислот с 18-20 углеродными атомами, имеющими различное положение в пространстве: трансизомер имеет прямую форму, а углеродная цепь цисизомера всегда изогнута в месте двойной связи. Ненасыщенные жирные кислоты являются только цисизомерами, т.е. они все изогнуты. Жирные кислоты в свободном состоянии редко встречаются в составе мембран. Они являются важным фактором регулирования проницаемости мембран (влияют на поверхностные свойства фосфолипидов, белок-липидные и липид-липидные взаимодействия), функционирования мембранно-связанных ферментов (П. Г. Богач и соавт., 1981). В мембранах располагаются ферменты, активность которых зависит от липидного окружения. В этом окружении ферменты имеют определенную конформацию. Изменение липидного окружения (делипидирование, использование липолитических ферментов, липидообменивающих белков) ведет к изменению конформации белков (ферментов), изменению их каталитической активности (Н. Е. Кучеренко, А. Н. Васильев, 1985). Активность ферментов в мембранах связана с вязкостью липидной фазы мембран, составом липидов (П. Г. Богач и соавт., 1981; Н. Е. Кучеренко, А. Н. Васильев, 1985). Метаболическая активность липидзависимых ферментов определяется изменениями в липидном микроокружении и в первую очередь это касается фосфолипидов: от их состава и метаболизма зависят ферментативные процессы. Это подтверждено для микросомальной монооксигеназной системы. Липидные молекулы являются матриксом, оптимальным для функционирования мембранно-связанных ферментов. НЖК в мембранах придают им такое качество, как жидкостность (текучесть). Увеличение в мембранах содержания холестерина, насыщенность жирнокислотных радикалов в фосфолипидах снижают жидкостность мембран. Подвижность липидов изменяет конформацию полярных головок. Регулирующее влияние на мембранно-связанные ферменты оказывают гликофосфолипиды (стабилизируя мембраны). При модификации липидного состава теряется чувствительность к гормонам, фосфолипиды влияют на функционирование рецепторов, могут регулировать их число (Н. Е. Кучеренко, А. В. Васильев, 1985), взаимодействовать с токсинами. Интенсивность обновления фосфолипидов зависит от скорости синтеза ДНК в клетке. Имеется связь синтеза ДНК с составом липидов, перераспределением фракций фосфолипидов, степенью ненасыщенности жирнокислотных радикалов (насыщенные жирные кислоты тормозят синтез ДНК). Такие фракции фосфолипидов, как фосфатидилэтаноламины, кардиолипины дестабилизируют молекулы ДНК путем усиления активности ДНК-полимеразы. Фосфолипиды влияют на прочность ДНК (стабилизацию структуры). Все эти данные (Н. Е. Кучеренко, А. Н. Васильев, 1985) свидетельствуют о важной регуляторной роли фосфолипидов мембран, составной частью которых являются ненасыщенные жирные кислоты.
Арахидоновая кислота и ее производные, их роль в обмене веществ.
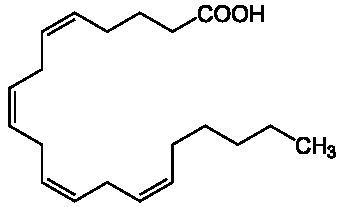
рахидоновая кислота - ( эйкозатетраеновая кислота ;5,8,11,14-эйкозатетpаеновая) - полиненасыщенная жирная кислота, входит в состав мембранных фосфолипидов тромбоцитов и эндотелиальных клеток. Высвобождается из фосфолипидов под действием фосфолипазы А2 и фосфолипазы С ( рис. 2.3 allerg), активируемых коллагеном, тромбином, АДФ и другими биологически активными веществами. Арахидоновая кислота является предшественником в синтезе оксилипинов см. рис. 1-1 и рис. 1-2. Свободная арахидоновая кислота быстро метаболизируется, превращаясь в высокоактивные соединения - простагландины и тромбоксаны.
В составе липидов арахидоновая кислота присутствует в мозге, печени и молочном жире млекопитающих. В фосфолипидах надпочечников арахидоновая кислота составляет около 20 % от суммы жирных кислот. При гидрировании арахидоновая кислота образует арахиновую кислоту, встречающуюся в маслах бобовых растений, в частности — арахиса.[1] Метаболиты АК являются эндогенными лигандами каннабиноидных рецепторов (1992 - 95 гг.). Наиболее важными среди них являются продукт неокислительного метаболизма арахидоновой кислоты - арахидонилэтаноламид (анандамид) и 2- арахидонилглицерин, 2-АГ Оба соединения выполняют функции нейромодулятора и нейромедиатора и служат "эндогенными каннабиоидами".
Охарактеризуйте простые липиды: ацилглицерины, воски. Напишите структурную формулу триацилглицерина, дайте его полное название (с указанием жирнокислотных остатков).
Глицериды (ацилглицерины, или ацилглицеролы *) представляют собой сложные эфиры трехатомного спирта глицерина и высших жирных кислот. Если жирными кислотами этерифицированы все три гидроксильные группы глицерина (ацильные радикалы R1, R2 и R3 могут быть одинаковы или различны), то такое соединение называют триглицеридом (триацилглицерол), если две – диглицеридом (диацилглицерол) и, наконец, если этерифицирована одна группа – моноглицеридом (моноацилглицерол).
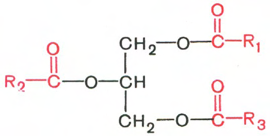
Триацилглицерол
Наиболее распространенными являются триглицериды, часто называемые нейтральными жирами или просто жирами. Нейтральные жиры находятся в организме либо в форме протоплазматического жира, являющегося структурным компонентом клеток, либо в форме запасного, резервного, жира. Роль этих двух форм жира в организме неодинакова. Протоплазматический жир имеет постоянный химический состав и содержится в тканях в определенном количестве, не изменяющемся даже при патологическом ожирении, в то время как количество резервного жира подвергается большим колебаниям. Как отмечалось, основную массу природных нейтральных жиров составляют триглицериды. Жирные кислоты в триглицеридах могут быть насыщенными и ненасыщенными. Из жирных кислот чаще встречаются пальмитиновая, стеариновая и олеиновые кислоты. Если все три кислотных радикала принадлежат одной и той же жирной кислоте, то такие триглицериды называют простыми (например, трипальмитин, тристеарин, триолеин и т.д.), если разным жирным кислотам, то смешанными. Названия смешанных триглицеридов образуются в зависимости от входящих в их состав жирных кислот, при этом цифры 1, 2 и 3 указывают на связь остатка жирной кислоты с соответствующей спиртовой группой в молекуле глицерина (например, 1-олео-2-пальмитостеарин). Необходимо отметить, что положение крайних атомов в молекуле глицерина на первый взгляд равнозначно, тем не менее их обозначают сверху вниз – 1 и 3. Это объясняется прежде всего тем, что в структуре триглицерида при пространственном ее рассмотрении крайние ≪глицериновые≫ атомы углерода становятся уже не равнозначными, если гидроксилы 1 и 3 ацилированы разными жирными кислотами. При необходимости применяют также систему стереохимической нумерации (обозначают sn –stereochemical numbering): например, 1,2-дистеарил-3-пальмитил-sn-глицерол.
По этой системе, если в проекции Фишера гидроксильная группа при 2-м углеродном атоме глицерина располагается слева, атому углерода, находящемуся над ним, присваивается номер 1, а расположенному под ним – номер 3. Действительно, углероды 1 и 3 глицерола, учитывая их пространственное расположение, неидентичны. Особенно четко это видно на примере молекулы триглицерида. Ферменты это различают и всегда специфичны только к одному из трех углеродов глицерина. Так, глицеролкиназа фосфорилирует глицерин в положении sn-3, в результате чего образуется глицерол-3-фосфат, но не глицерол-1-фосфат. Жирные кислоты, входящие в состав триглицеридов, практически определяют их физико-химические свойства. Так, температура плавления триглицеридов повышается с увеличением числа и длины остатков насыщенных жирных кислот. Напротив, чем выше содержание ненасыщенных жирных кислот, или кислот с короткой цепью, тем ниже точка плавления. Животные жиры (сало) обычно содержат значительное количество насыщенных жирных кислот (пальмитиновой, стеариновой и др.). благодаря чему при комнатной температуре они твердые. Жиры, в состав которых входит много ненасыщенных кислот, при обычной температуре жидкие и называются маслами. Так, в конопляном масле 95% всех жирных кислот приходится на долю олеиновой, линолевой и линоленовой кислот и только 5% – на долю стеариновой и пальмитиновой кислот. В жире человека, плавящемся при температуре 15°С (при температуре тела он жидкий), содержится 70% олеиновой кислоты. Глицериды способны вступать во все химические реакции, свойственные сложным эфирам. Наибольшее значение имеет реакция омыления, в результате которой из триглицеридов образуются глицерол и жирные кислоты. Омыление жира * может происходить как при ферментативном гидролизе, так и при действии кислот или щелочей.
Воска – сложные эфиры высших жирных кислот и высших одноатомных или двухатомных спиртов с числом углеродных атомов от 16 до 22. Общие их формулы можно представить так:
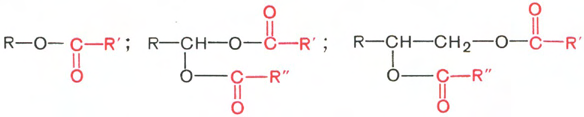
В этих формулах R, R' и R" – возможные радикалы.
Воска могут входить в состав жира, покрывающего кожу, шерсть, перья. У растений 80% от всех липидов, образующих пленку на поверхности листьев и плодов, составляют воска. Известно также, что воска являются нормальными метаболитами некоторых микроорганизмов. Природные воска (например, пчелиный воск, спермацет, ланолин) обычно содержат, кроме указанных сложных эфиров, некоторое количество свободных жирных кислот, спиртов и углеводородов с числом углеродных атомов 21–35.
Глицерофосфолипиды, их строение, разнообразие и свойства. Приведите примеры и напишите структурные формулы.
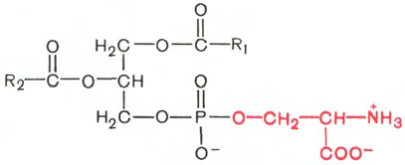
Глицерофосфолипиды являются производными фосфатидной кислоты. В их состав входят глицерин, жирные кислоты, фосфорная кислота и обычно азотсодержащие соединения. Общая формула глицерофосфолипидов выглядит так:

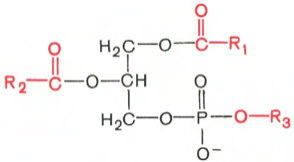
Фосфатидная кислота Глицеролфосфолипид
В этих формулах R1 и R2 – радикалы высших жирных кислот, a R3 – чаще радикал азотистого соединения. Для всех глицерофосфолипидов характерно, что одна часть их молекул (радикалы R1 и R2 ) обнаруживает резко выраженную гидрофобность, тогда как другая часть гидрофильна благодаря отрицательному заряду фосфорной кислоты и положительному заряду радикала R3.
Из всех липидов глицерофосфолипиды обладают наиболее выраженными полярными свойствами. При помещении глицерофосфолипидов в воду в истинный раствор переходит лишь небольшая их часть, основная же масса липидов находится в виде мицелл. Существует несколько групп (подклассов) глицерофосфолипидов. В зависимости от характера азотистого основания, присоединенного к фосфорной кислоте, Глицерофосфолипиды подразделяют на фосфатидилхолины (лецитины), фосфатидилэтаноламины (кефалины) и фосфатидилсерины. В состав некоторых глицерофосфолипидов вместо азотсодержащих соединений входит не содержащий азота шестиуглеродный циклический спирт инозит, называемый также инозитолом. Эти липиды называются фосфатидилинозитолами.
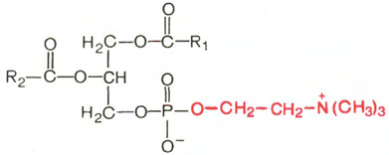
Фосфатидилхолины (лецитины). В отличие от триглицеридов в молекуле фосфатидилхолина одна из трех гидроксильных групп глицерина связана не с жирной, а с фосфорной кислотой. Кроме того, фосфорная кислота в свою очередь соединена эфирной связью с азотистым основанием – холином [НО—СН2—СН2—N+(CH3)3]. Таким образом, в молекуле фосфатидил-холина соединены глицерин, высшие жирные кислоты, фосфорная кислота и холин:
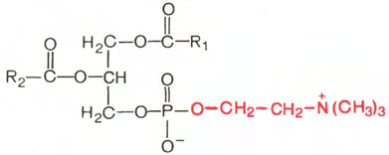
Фосфатидилхолин
Фосфатидилэтаноламины. Основное различие между фосфатидилхолинами и фосфатидилэтаноламинами – наличие в составе последних азотистого основания этаноламина (HO—CH2—CH2—N+H3):
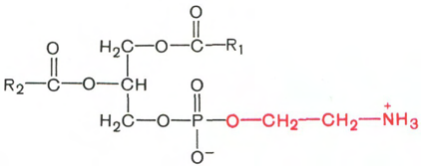
Фосфатидилэтаноламин
Из глицерофосфолипидов в организме животных и высших растений в наибольшем количестве встречаются фосфатидилхолины и фосфатидилэтаноламины. Эти 2 группы глицерофосфолипидов метаболически связаны друг с другом и являются главными липидными компонентами мембран клеток.
Фосфатидилсерины.
В
молекуле фосфатидилсерина азотистым
соединением служит остаток аминокислоты
серина
![]()
Фосфатидилсерин
Фосфатидилсерины распространены гораздо менее широко, чем фосфатидилхолины и фосфоэтаноламины, и их значение определяется в основном тем, что они участвуют в синтезе фосфатидилэтаноламинов.
Глицерофосфолипиды являются производными фосфатидной кислоты. Напишите формулу фосфатидной кислоты. Какими кислотами этерифицированы спиртовые группы большинства природных глицерофосфолипидов?

Фосфатидная кислота
Какими кислотами???
Напишите формулы: а- и b-фосфатидилхолинов, кардиолипина. Охарактеризуйте эти соединения.
Фосфатидилхолин
В отличие от триглицеридов в молекуле фосфатидилхолина одна из трех гидроксильных групп глицерина связана не с жирной, а с фосфорной кислотой. Кроме того, фосфорная кислота в свою очередь соединена эфирной связью с азотистым основанием – холином [НО—СН2—СН2—N+(CH3)3]. Таким образом, в молекуле фосфатидил-холина соединены глицерин, высшие жирные кислоты, фосфорная кислота и холин
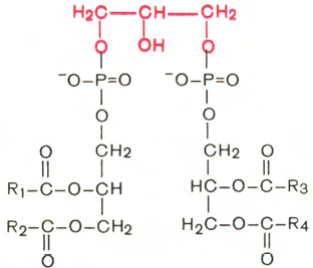
Кардиолипин
Своеобразным представителем глицерофосфолипидов является кардиолипин, впервые выделенный из сердечной мышцы. По своей химической структуре кардиолипин можно рассматривать как соединение, в котором 2 молекулы фосфатидной кислоты связаны с помощью одной молекулы глицерина. В отличие от остальных глицерофосфолипидов кардиолипин является как бы ≪двойным≫ глицерофосфолипидом. Кардиолипин локализован во внутренней мембране митохондрий. Функция его пока неясна, хотя известно, что в отличие от других фосфолипидов кардиолипин обладает иммунными свойствами.
93. Церамиды и сфингомиелины
Церамидами называют ?-ацильные производные сфингозинов, в которых аминогруппа основания ацилирована жирной кислотой. Эти соединения широко распространены в тканях растений и животных, но количества их незначительны. Церамиды различаются остатками жирных кислот, входящими в их состав. Жирные кислоты, присоединенные амидными связями к аминогруппе основания в молекуле церамида, содержат различное число атомов углерода (Ci6, Ci8, С22 или С24). Сфингомиелины могут рассматриваться как фосфохолиновые производные церамидов и являются одной из наиболее важных групп фосфолипидов.
О ? II
ОН N—CR
СН3(СН2)12—СН=СН—СН—СН—СН2ОН цераммЭ
н ?
ОН N—CR О CH3(CH2)12-CH=CH-CH-CH-CH2-0-P-OCH2CH2N=(CH3)3
о-сфмнгомиелмн
Хотя сфингомиелины по химическому строению и отличаются от фосфатидилхолина, эти соединения проявляют определенное сходство, обусловленное типичной для амфифилов структурой, а именно наличием ионной фосфорилхолиновой группировки и двух длинных углеводородных целей.
Впервые сфингомиелины были обнаружены в нервной ткани, однако они входят в состав липидов крови, а также встречаются во многих других тканях. Кроме обычных сфингозинов, рассмотренных выше, сфингомиелин из мозга содержит полиненасыщенные сфингозины, называемые дегидросфингозинами.
95. ЦЕРЕБРОЗИДЫ, сфинголипиды общей ф-лы RCH(OH)CH(NHCOR')CH2OX, где R и R' - алкил, гидроксиалкил, алкенил; X - остаток глюкозы (глюкоцереброзиды), галактозы (галактоцереброзиды) или фукозы (фукоцереброзиды). В глюко- и галакто-цереброзидах присутствует-гликозидная связь, в фуко-цереброзидах-гликозидная связь.
Цереброзиды- твердые в-ва, не раств. в воде, хорошо раств. в орг. р-рителях. При кислотном гидролизе образуют жирные к-ты, сфингозины (сфингозиновые основания) RCH(OH)CH(NH2)CH2OH и углеводы.
Цереброзиды содержатся в тканях человека, животных и высших растений. С их функционированием связана деятельность организма в целом и на клеточном уровне (изолирующие свойства миелиновой оболочки нервных аксонов, межклеточное узнавание, иммунологич. р-ции). Биосинтез цереброзидов протекает по двум путям: гликозилирование церамидов с помощью уридиндифосфатсахаров или гликозилирование сфингозинов с послед. N-ацилированием. Нарушения метаболизма цереброзидов, обусловленные снижением активности или отсутствием ферментов их гидролиза, имеют наследственный характер (т. наз. болезнь Краббе); при этом в органах и тканях, особенно в мозге, накапливаются галактоцереброзиды. Повышение содержания глюкоцереброзидов в мозге, печени свидетельствует о нарушениях обмена ганглиозидов (болезнь Гоше).
Осуществлен полный хим. синтез и полусинтез цереброзидов, их аналогов и меченых производных, используемых в качестве диагностич. и исследовательских биопрепаратов. Осн. путь синтеза - гликозилирование бензоилцерамидов RCH(OCOC,H5)CH(NHCOR')CH2OH по Кёнигса-Кнорра реакции. Наиб. стереоселективно с образованием-аномера гликозилирование происходит в присут. цианида ртути. Другой подход - ацилирование психозина (сфингозил--галактозида), полученного при избират. гидролизе прир. цереброзидов.
Типичными представителями цереброзидов головного мозга являются галактоцереброзиды на основе 4-сфингенина CH3(CH2)12CH = CHCH(OH)CH(NH2)CH2OH: френозин [в общей ф-ле R' = C22H25CH(OH)], т. пл. 195 °С; церазин (R' = С23H47), т. пл. 182 °С; нервон [R' = СН3(СН2)6СН = = СН(СН2)13].
96. При гидролизе распадаются на сфингозин, галактозу или глюкозу, а также остаток жирной кислоты. Цереброзиды содержатся в больших количествах в составе мозга, особенно в белом веществе мозга и мякотных нервных волокнах.
Избыточное накопление цереброзидов в организме и появление их в больших количествах в печени, селезенке, костном мозге и лимфатических узлах связано с некоторыми заболеваниями нервной системы — так называемой нейролипоидозами. Среди этих жирных кислот чаще всего встречается лигноцериновая, нервоновая и цереброновая кислоты, т.е. жирные кислоты, имеющие 24 углеродных атома. Структура цереброзидов может быть представлена следующей схемой:
Сфингозин - СН2-О— гексоза (чаще галактоза)
0 = С — остаток жирной кислоты
97) ГАНГЛИОЗИДЫ (сиалогликосфинголипиды), гликосфин-голипиды общей ф-лы , где Х - углеводная цепь, содержащая остатки сиаловых к-т, связанных с остатком галактозы или гексозамина (см., напр., ф-лу I в ст. Гликосфинголипиды; Y - остаток N-ацетил- или N-гликолилнейраминовой к-ты, к-рые наз. сиаловыми к-тами - см. ф-лу).
В названиях ганглиозидов используют сочетание из двух букв - G (указывает принадлежность соед. к ганглиозидам) и М, D, Т или Q (показывают число сиаловых к-т в молекуле - соотв. 1, 2, 3 или 4) и цифры (иногда с буквами) в ниж. индексе, обозначающие конкретное в-во. Соед. I (ф-ла в ст. Гликосфинголипиды, Y-остаток N-ацетилнейраминовой к-ты) по этой номенклатуре обозначается GM1. В соответствии с правилами ИЮПАК это соед. обозначается II3AcNeuGgOse4Cer, где римская цифра показывает положение остатка моносахарида (считая от церамида; в общей ф-ле Х-Н), с к-рым связан остаток сиаловой к-ты, цифра в верх. индексе-номер углеродного атома в цикле, с к-рым связана эта к-та, AcNeuGgOse4-oлигocaxapиднaя цепь (AcNeu-остаток N-ацетилнейраминовой к-ты, GgOse4- остаток ганглиотет-раозы), Сеr-остаток церамида.
Ганглиозиды широко распространены в тканях животных, особенно в мозге. В растениях и микроорганизмах не встречаются. Локализованы в осн. на пов-сти плазматич. мембраны клетки. Как и др. Гликосфинголипиды участвуют в процессах регуляции роста и адгезии клеток, межклеточных взаимодействиях, иммунология, процессах. Могут входить в состав рецепторов токсинов (напр., GMj входит в состав рецептора токсина холеры), пептидных гормонов, нек-рых вирусов и интерферона.
В клетке ганглиозиды синтезируются в аппарате Гольджи путем переноса остатков моносахаридов и сиаловых к-т от уридиндифосфатсахаров и цитидинмонофосфатсиаловых к-т в присут. специфич. гликозилтрансфераз и сиалилтрансфераз к церамиду. При нарушении метаболизма ганглиозидов возникают болезни (ганглиозидозы). Изменение биосинтеза ганглиозидов происходит и при злокачественной трансформации клеток. В препаративных кол-вах ганглиозиды получают из прир. источников, в осн. из мозга.
98. Стерины
Стеролы, циклические спирты, относящиеся к классу стероидов (См. Стероиды), широко распространены в живой природе. Большинство С. — оптически-активные кристаллические вещества, растворимые в органических растворителях и нерастворимые в воде. Общим предшественником С. у микроорганизмов, растений и животных служит углеводород Сквален, превращающийся в циклический ланостерин (С30Н50О) или в его изомер циклоартенол, из которых затем образуются различные С., содержащие 27—29 атомов углерода. Важнейший С. животных — Холестерин, самый распространённый С. грибов, в том числе дрожжей,— Эргостерин, а из обширной группы С. растений (фитостеринов (См. Фитостерины)) наиболее распространены β-ситостерин и стигмастерин.
У высших животных С. содержатся в нервной ткани, печени, клетках спермы и т.д.: образуя с высшими жирными кислотами сложные эфиры, они служат их переносчиками в организме. В растениях С. находятся в свободном состоянии или в соединении с жирными кислотами (т. н. стериды), углеводами (т. н. фитостеролины) или с теми и другими вместе. С., поступающие с пищей, используются для синтеза гормонов насекомыми. которые не обладают ферментами, необходимыми для первых стадий биосинтеза С. К С. относят гормоны линьки насекомых (см. Экдизон (См. Экдизоны)) и антеридиол, индуцирующий половое размножение некоторых низших грибов. Близки к С. витанолиды (ненасыщенные Лактоны) и кукурбитацины (ненасыщенные горькие вещества из тыквенных растений), а также сапогенины морских беспозвоночных и входящие в состав Ланолина и восков ланостерин и др. тритерпены со стерановым углеродным скелетом (см. Стероиды). Наиболее изученные биохимич. функции С. — превращение холестерина в стероидные гормоны и эргостерина в витамины группы D под действием ультрафиолетового света. С. входят в состав биологических мембран (См. Биологические мембраны) как их структурные компоненты. Доступные С. (холестерин, эргостерин, (β-ситостерин) используются как сырьё для промышленного получения стероидных гормонов и витаминов группы D.
99.Ж ЕЛЧНЫЕ КИСЛОТЫ, монокарбоновые гидроксикислоты, относящиеся к классу стероидов. Почти все желчные кислоты - производные прир. холановой к-ты (ф-ла Iа).
Наиб. распространены ее моно-, ди- и тригидроксизамещенные, содержащие 24 атома С; известны также ди-, три- и тетрагидроксизамещенные желчные кислоты, содержащие 27 (Iб) и 28 (Iв) атомов С. Положение заместителей, расположенных под или над плоскостью молекулы, обозначают соотв. буквами a и b. Желчные кислоты - кристаллы; плохо раств. в воде (см. табл.). Образование желчных кислот происходит гл. обр. в печени нек-рых птиц, млекопитающих и человека из холестерина. Биосинтез желчных кислот из холестерина включает след. р-ции: гидроксилирование по атомам углерода колец В и С, инверсию 3b- в 3a-гидроксигруппу, восстановление двойной связи при атоме С-5 с получением цис-сочленения колец А и В, окислит. отщепление изопропильной группы в боковой цепи с образованием карбоксильной при атоме С-24.
Для млекопитающих характерно наличие в молекуле желчных кислот 24, для земноводных - 27 атомов С. В желчи человека содержатся хенодезоксихолевая, холевая и дезоксихолевая к-ты (соотв. 45, 31 и 24% от общего кол-ва желчных кислот); в желчи быка - гл. обр. холевая и дезоксихолевая к-ты (30 и 10% соотв.). Желчные кислоты содержатся в желчи в виде щелочных солей т. наз. конъюгатов - соединений желчных кислот с таурином или глицином. Так, для холевой к-ты конъюгаты - гликохолевая к-та (в ф-ле I R = CONHCH2COOH) и таурохолевая к-та (R = = CONHCH2CH2SO3H). Натриевые соли желчных кислот - хорошие эмульгаторы; эмульгируя жиры, они способствуют их всасыванию и перевариванию. Важную роль при этом играют, в частности, р-римые комплексы дезоксихолевой к-ты с жирами и жирными к-тами (холеиновые к-ты). Желчные кислоты получают щелочным гидролизом твердого в-ва желчи: нагревают желчь с 5-10%-ным р-ром КОН или NaOH в автоклаве при 120-160°С в течение 8-16 ч, затем подкисляют разб. НСl и экстрагируют эфиром или этилацетатом. На основе прир. желчных кислот осуществлен синтез разл. гидрокси- и оксопроизводных холановой к-ты. Так, дегидратация холевой к-ты при нагр. в вакууме или при действии ZnCl2 в ацетоне приводит к апохолевой к-те [3,12-дигидрокси-5b-хола-8(14)-ен-24-овая к-та], из к-рой м. б. получены холеиновые к-ты. Желчные кислоты применяют для синтеза кортикостероидов и в медицине в качестве лек. препаратов, растворяющих и предотвращающих образование желчных камней (напр., хенодезоксихолевая к-та и ее Mg-соль, урсодезоксихолевая к-та, этиловый эфир 3,7,12-тригидроксихолановой к-ты).
100. Продукция гормонов аденогипофиза зависит от либеринов и статинов - гормонов гипоталамуса , поступающих в воротную систему гипофиза ( рис. 9.1 ).Рилизинг-факторы, или либерины, стимулирующт синтез и выделение гормонов клетками передней доли гипофиза .
Секреция либеринов и статинов контролируется адренергическими , холинергическими и дофаминергическими нейронами высших нервных центров . Кроме того, секреция некоторых гормонов аденогипофиза и либеринов тормозится гормонами периферических эндокринных желез по принципу отрицательной обратной связи. Таким образом, в регуляции роста участвуют гормоны гипоталамуса , аденогипофиза и периферических эндокринных желез - мишеней аденогипофизарных гормонов .
Семейство рилизинг-факторов (гипофизарных либеринов и статинов ) включает небольшие пептиды, образующиеся в нейронах мелкоклеточных ядер медиобазального и отчасти заднего гипоталамуса и хранящиеся в срединном возвышении нейрогипофиза . Рилизинг-факторы, по-видимому, образуются и накапливаются в различных отделах мозга .
К настоящему времени известны 10 рилизинг-факторов, участвующих в регуляции секреции гормонов аденогипофиза .
В 1970-1972 гг группами Гиллемина и Шэлли впервые была расшифрована первичная структура и впервые осуществлен полный химический синтез этих факторов. В настоящее время осуществлен молекулярно-биологический синтез мРНК соматостатина и его гена. Синтезированный ген удалось ввести в бактериальную плазмиду и с ее помощью создать штамм соматостатинпродуцирующих бактерий. Названные рилизинг-факторы состоят из небольшого количества аминокислотных остатков. Так, Тиролиберин является трипептидом, ЛГ/ФСГ-РФ - декапептид, соматостатин - тетрапептид. Их структуры, по-видимому, не обладают видовой специфичностью, в то время как для факторов, контролирующих секрецию МСГ , видовые особенности структуры исключить нельзя.
Предполагают, что у млекопитающих МРФ и МИФ могут быть производными окситоцина: МИФ - боковой цепью, МРФ - лишенными кольцевой структуры и одного остатка цистеина его N-кольцевого фрагмента.
Рилизинг-факторы не всегда обладают строго специфической направленностью действия. Так, ТРФ - стимулятор секреции не только тиреотропного гормона ( ТТГ ), но и пролактина . ЛГ-РФ стимулирует продукцию лютеинизирующего гормона ( ЛГ ) и фолликулостимулирующего гормона ( ФСГ ).
Соматостатин обладает довольно широким спектром ингибирующих эффектов на эндокринные функции: он тормозит секрецию соматотропина ( СТГ ), индуцированную секрецию ТТГ и пролактина передней долей гипофиза , секрецию инсулина и особенно глюкагона поджелудочной железой , а также гормонов желудочно-кишечного тракта . Интересно, что соматостатин образуется не только в гипоталамусе , но и в островковом аппарате поджелудочной железы . МИФ не только тормозит секрецию МСГ средней доли гипофиза , но и оказывает влияние на корковые и подкорковые структуры головного мозга . ТРФ - антидепрессант .
101. Питуитарная железа, называемая также гипофизом, — это мелкая железа около 1 см в диаметре и массой от 0,5 до 1 г, которая лежит в турецком седле (костном образовании основания черепа) и связана с гипоталамусом посредством питуитарного, или гипофизарного, стебля. Физиологически питуитарная железа подразделяется на две независимые части: переднюю долю гипофиза, или аденогипофиз, и заднюю долю гипофиза, или нейрогипофиз. Между ними есть промежуточная доля, относительно плохо васкуляризированная, практически не выраженная у человека и более заметная как структурно, так и функционально у низкоорганизованных организмов.
В эмбриогенезе эти две доли гипофиза развиваются из разных источников: передняя доля — из кармана Ратке, который эмбриологически является инвагинацией фарингеального эпителия, а задняя доля — из нервной ткани, являющейся выростом гипоталамуса. Происхождением передней доли гипофиза из фарингеального эпителия объясняется эпителиоидная структура ее клеток, а наличие глиального типа клеток в задней доле гипофиза — его нейрогенной природой. Передним гипофизом продуцируются 6 важных гормонов-пептидов и несколько менее существенных, в заднем гипофизе содержатся 2 гормона-пептида. Гормоны аденогипофиза играют важную роль в контроле метаболических функций организма:
• гормон роста обеспечивает рост организма, увеличение количества клеток и их дифференцировку, стимулируя образование белков;
• адренокортикотропный гормон (кортикотропин) управляет секрецией некоторых гормонов коры надпочечников, которые, в свою очередь, влияют на метаболизм глюкозы, белков и жиров;
• тиреотропный гормон (тиреотропин, ТТГ) контролирует скорость секреции тироксина и трийодтиронина щитовидной железой. В свою очередь, они регулируют в организме скорость основных химических реакций;
• пролактин обеспечивает развитие молочных желез и продукцию молока;
• два самостоятельных гонадотропных гормона — фолликулостимулирующий и лютеинизирующий — контролируют рост яичников и семенников, их гормональную и репродуктивную активность. Роль гормонов заднего гипофиза иная:
• антидиуретический гормон (также называемый вазопрессином) контролирует скорость экскреции воды с мочой, помогая регулировать количество воды в организме;
• окситоцин стимулирует роды и способствует молокоотделению из молочных желез во время кормления ребенка.
Передняя доля гипофиза содержит много различных типов клеток, синтезирующих и высвобождающих гормоны. Для синтеза определенного типа гормонов в аденогипофизе существует определенный тип клеток. С помощью специальных красителей, реагирующих с комплексом антиген-антитело, где антигеном является определенный гормон, смогли выделить пять разных типов клеток.
1. Соматотропы — гормон роста человека.
2. Кортикотропы — адренокортикотропин.
3. Тиреотропы — тиреотропный гормон.
4. Гонадотропы — гонадотропные гормоны, включая лютеинизирующий гормон, фолликулостимулирующий гормон.
5. Лактотропы — пролактин.
Около 30-40% клеток переднего гипофиза представлены соматотропами, вырабатывающими гормон роста, 20% — кортикотропами, секретирующими АКТГ. Каждый из остальных типов клеток составляет не более 3-5% общего количества, но они секретируют чрезвычайно мощные гормоны, контролирующие функции щитовидной железы, половые функции и секрецию молока.
Соматотропы, избирательно окрашиваемые только кислыми красителями, называют ацидофильными, поэтому опухоли гипофиза, секретирующие большое количество гормона роста, называют ацидофильными опухолями.
Гормоны задней доли гипофиза синтезируются клетками гипоталамуса. Клетки, секретирующие гормоны нейрогипофиза, представляют собой большие нейроны, названные крупноклеточными, и располагаются в супраоптическом и паравентрикулярном ядрах гипоталамуса. Из передних отделов гипоталамуса гормоны транспортируются аксоплазмой нервных волокон в гипофиз.
102.Тиреотропные гормоны
Физиологическими свойствами и производимые в щитовидной железе.
Синтез тиреоидных гормонов в присутствии тиреоидной пероксидазы.[1]
Щитовидная железа производит два тиреоидных гормона, отличающихся лишь наличием или отсутствием одного дополнительного атома йода в молекуле — тироксин (T4) и трийодтиронин (T3). При этом тироксин является относительно малоактивным тиреоидным гормоном, фактически — прогормоном, и слабо связывается непосредственно с рецепторами тиреоидных гормонов в тканях.
Перед тем, как оказать действие на клетки органов-мишеней, большая часть тироксина непосредственно в клетках конвертируется в биологически активную форму — трийодтиронин. Этот процесс происходит при участии металлофермента — селен-зависимой монодейодиназы. При дефиците микроэлемента селена в организме или при генетическом дефекте монодейодиназы, предопределяющем её пониженную активность в тканях, развивается состояние недостаточности гормона щитовидной железы, несмотря на кажущийся нормальным уровень T4 в плазме крови — так называемый англ. euthyroid sick syndrome. Аналогичное состояние может наблюдаться при приёме некоторых лекарственных веществ, угнетающих активность тканевой монодейодиназы.
От 2/3 до 4/5 общего количества тиреоидных гормонов, производимых щитовидной железой, поступает в кровь в форме «прогормона» тироксина, и лишь 1/3-1/5 — в форме биологически активного трийодтиронина.
103.Гормоны поджелудочной железы
Поджелудочная железа является органом смешанной секреции. С одной стороны, она вырабатывает пищеварительные ферменты, которые по специальным каналам поступают в двенадцатиперстную кишку, при этом они находятся в неактивном состоянии. С другой стороны, ее клетки синтезируют рад гормонов, предназначенных для регуляции работы внутренних органов. Ответственны за синтез этих соединений особые клеточные скопления, называемые островками Лангерганса по имени ученого, который их открыл. Они рассредоточены по всему телу железы и не имеют специальных выводных путей. Их секрет поступает непосредственно в кровь и доставляется к органам-мишеням.
Основными гормонами поджелудочной железы являются следующие соединения:
Инсулин
Глюкагон
С-пептид
За синтез инсулина отвечают бета-клетки островков Лангерганса, его основная функция в организме состоит в понижении уровня сахара в крови. Это достигается с помощью одновременного действия по трем направлениям. Инсулин приостанавливает образование глюкозы в печени и повышает количество сахара, который усваивается тканями организма за счет увеличения проницаемости клеточных мембран. В то же время он тормозит распад глюкагона, ведь тот является полимерной цепочкой, состоящей из молекул глюкозы, и может быть использован для увеличения ее концентрации в крови. Под действием ряда факторов в организме возникает недостаток инсулина, ведущий к развитию сахарного диабета.
Выработкой глюкагона занимаются альфа-клетки островков Лангерганса, он отвечает за увеличение концентрации глюкозы в кровяном русле. Это достигается путем стимулирования ее образования в печени. Кроме того, он способствует расщепление липидов в жировой ткани. Таким образом, два описанных выше гормона поджелудочной железы выполняют противоположные функции. Однако в поддержании нормального уровня сахара в крови участвуют и другие биологически активные соединения, вырабатываемые эндокринной системой - соматотропин (гормон роста), кортизол, адреналин.
В диагностике таких заболеваний, как сахарный диабет, ожирение, акромегалия и различные патологии печени, используют анализ крови на инсулин. С-пептид, строго говоря, не является гормоном поджелудочной железы. Он представляет собой часть молекулы проинсулина, которая отделяется от нее в процессе синтеза и оказывается в кровяном русле. Поэтому количество С-пептида в сыворотке крови эквивалентно количеству инсулина, которое секретируется поджелудочной железой. Этот показатель более точен, ведь белковый фрагмент не обладает биологической активностью и не вступает в химические реакции.
В настоящее время его используют в диагностике сахарного диабета I и II типа, опухоли поджелудочной железы и заболеваний печени. Повышенное содержание С-пептида свидетельствует о наличии в организме инсулиномы. Если оно сохраняется после прохождения курса лечения, то велика вероятность развития метастазов или рецидивов заболевания. С-пептид позволяет определять уровень инсулина у больных сахарным диабетом, получающих лечение, и соответственно корректировать характер терапевтических мероприятий. Кроме того, он помогает отслеживать состояние плода у женщин, страдающих диабетом беременных.
104. Гормоны гипоталамуса — важнейшие регуляторные гормоны, производимые гипоталамусом. Все гормоны гипоталамуса имеют пептидное строение и делятся на 3 подкласса: рилизинг-гормоны стимулируют секрецию гормонов передней доли гипофиза, статины тормозят секрецию гормонов передней доли гипофиза, и гормоны задней доли гипофиза традиционно называются гормонами задней доли гипофиза по месту их хранения и высвобождения, хотя на самом деле производятся гипоталамусом.
Гормоны гипоталамуса играют одну из ведущих ролей в деятельности всего человеческого организма. Вырабатываются эти гормоны в отделе головного мозга под названием гипоталамус. Без исключения все эти вещества являются пептидами. При этом все эти гормоны различают трех видов: рилизинг-гормоны, статины и гормоны задней доли гипофиза. В подкласс рилизинг-гормонов гипоталамуса входят следующие гормоны:
кортикотропин-рилизинг-гормон
соматотропин-рилизинг-гормон
тиреотропин-рилизинг-гормон
гонадотропин-рилизинг-гормон.
В подкласс гормонов задней доли гипофиза входят:
антидиуретический гормон, или вазопрессин
окситоцин
Вазопрессин и окситоцин синтезируются в гипоталамусе, а затем поступают в гипофиз. Функция регуляции секреции.
105) КАТЕХОЛАМИНЫ, группа биогенных аминов производных пирокатехина (катехола), осуществляющая регуляцию ф-ций эндокринных желез (надпочечники, щитовидная железа и др.) и передачу нервных импульсов. В первом случае катехоламины рассматривают как гормоны, во втором - как нейромедиаторы. Иногда к катехоламинам относят их синтетич. производные (см. Адреномиметические средства). К биогенным катехоламинам относятся дофамин (ф-ла I; R = R' =Н), L-норадреналин (I; R=ОН, R'=Н) и L-адреналин (I; R=ОН, R'=СН3). Осн. ф-ция дофамина-стимуляция секреции соматотропина (гормона роста) и подавление секреции пролактина (гормона, стимулирующего рост молочных желез, лактацию и др.). Известно также стимулирующее влияние дофамина на гликогенолиз, уровень глюкозы в крови, диурез, кровоток в почках. Нарушение синтеза дофамина в мозгу - причина возникновения болезни Паркинсона. Дофамин выделяется из пресинаптич. окончаний и действует на постсинаптич. рецепторы, к-рые либо активируют фермент адепилатцтлазу (рецепторы типа Д-1), либо оказывают на аденилатцик-лазу ингибирующее влияние и повышают уровень внутриклеточного Са2+ в результате активации фосфоинозитидного обмена (гидролиза полифосфоинозитидов до диацил-глицерина и инозитолфосфатов) или открывания рецепторрегулируемых Са-каналов (рецепторы Д-2). Все биол. эффекты дофамина связаны с его участием в регуляции фос-форилирования внутриклеточных белков. Биосинтез дофамина осуществляется из фенилаланина через тирозин (II) по схеме:
Лимитирующая стадия - синтез дигидроксифенилаланина (ДОФА), к-рая катализируется ферментом тирозингидроксилазой. Последняя активируется Са2+, поэтому факторы, повышающие концентрацию Са2+ в аксоплазме (часть цитоплазмы нейрона, входящая в состав аксона), ускоряют синтез дофамина. Стадия превращения ДОФА в дофамин катализируется декарбоксилазой ароматич. L-аминокислот. Дофамин накапливается гл. обр. в разл. отделах центр, нервной системы. В аксонах симпатич. нервных волокон и в мозговом слое надпочечников дофамин - промежут. метаболит. В этих тканях он при участии b-гидроксилазы гидроксилируется до L-норадреналина. Последний в мозговом слое надпочечников при участии фениламиноэтанол-N-метилтрансферазы и источника метильных групп S-аденозилметионина превращ. в L-адреналин. Нервные импульсы, идущие по симпатич. волокнам, приводят к выбросу L-норадреналина из пресинаптич. мембраны этих волокон и рецепции нейромедиатора на постсинаптич. мембране. В мозговом слое надпочечников секреция этих катехоламинов вызывается путем возбуждения, приходящего по чревному нерву. При этом в кровь поступают преим. L-адреналин и малые кол-ва L-норадреналина. Рецепторы этих катехоламинов есть практически во всех тканях человека и животных. L-Норадреналин взаимод. преим. с a-, a L-адреналин - с a- и b-адренергич. рецепторами. Через a-рецепторы (активируют фосфоинозитидный обмен) осуществляются вазоконстрикторные эффекты катехоламинов (приводят к сужению кровеносных сосудов), стимулируется гликогенолиз, секреция адренокортикотропина, инсулина, ренина и др. физиологически активных в-в. Через b-рецепторы (активируют аденилатциклазу) катехоламины уменьшают периферич. сопротивление сосудов, стимулируют гликогенолиз, липолиз и белковый обмен, повышают частоту и силу сердечных сокращений. Активация синтеза катехоламинов-типичная р-ция на стресс (напр., у животных концентрация адреналина в крови под влиянием страха может возрастать в 100 раз за неск. с); их роль заключается в мобилизации организма на осуществление активной мозговой и мышечной деятельности. Важнейшую роль катехоламины играют также в поддержании гомеостаза (постоянства состава внутр. среды и устойчивости осн. физиол. ф-ций) и адаптации организма. Удаление катехоламинов из синаптич. щели происходит путем обратного захвата в пресинаптич. окончания. Этот активный транспорт происходит с использованием энергии градиента Na+ на мембране. Часть катехоламинов диффундирует из синапса в межклеточное пространство и затем поступает в кровь. Инактивируются катехоламины путем окислит, дезаминирования, катализируемого катехол-О-метилтрансферазой. Метаболиты катехоламинов удаляются из организма почками, поэтому их содержание в моче может указывать на активность симпатич. нервной системы и мозгового слоя надпочечников. Нарушения в метаболизме, секреции и обратном
захвате катехоламинов-причина мн. патологич. процессов. Препараты, воздействующие на обмен катехоламины и их рецепцию (напр., пропранолол, ф-ла III), широко используют для лечения ряда заболеваний человека.
106. Производные аминокислот:
-производные тирозина: тироксин, трийодтиронин, дофамин, адреналин, норадреналин;
-производные триптофана: мелатонин, серотонин; -производные гистидина: гистамин.
107.Пептидные гормоны - наиболее многочисленный и разнообразный по составу и вариабельный в сравнительно-биологическом плане класс гормональных соединений. По особенностям химической структуры, свойств и физиологическим функциям входящих в него гормонов этот класс можно разделить на семейства:
1) нейрогипофизарных пептидов ;
2) гипоталамических пептидных рилизинг-факторов ;
3) ангиотензинов ;
4) олигопептидных гипофизарных гормонов ряда АКТГ ;
5) олигопептидных гормонов типа глюкагона и гормонов желудочно- кишечного тракта ; 6) инсулина ;
7) полипептидных гормонов, регулирующих обмен кальция , и полипептидных гормонов, регулирующих обмен фосфора ;
8) одноцепочечных (мономерных) белково-пептидных гормонов ряда СТГ ;
9) димерных гликопротеидных гормонов .
Предполагается, что представители каждого из большинства перечисленных семейств возникли на самых ранних стадиях эволюции позвоночных из общего гормонального предшественника путем серий последовательных мутаций и дупликаций кодирующего гена, а также ассоциаций модифицированных генов в более крупные.
Это предположение не относится к семейству паратгормона и кальцитонина . В основе типологии гормонов в указанном случае лежит не эволюционно-структурный принцип, а направленность их физиологических эффектов.
К пептидным гормонам относятся также эритропоэтин , гормоны тимуса , соматомедины , некоторые нейросекреторные гормоны насекомых и т.д.
Анализ функциональных свойств различных участков пептидной цепи нейрогипофизарных гормонов показал, что за связывание их с рецепторами соответствующих органов-мишеней ответственна кольцевая часть молекулы гормона и прежде всего аминокислота, стоящая в 3-м положении.
Очевидно, наличие в 3-м положении Фен обеспечивает наилучшее связывание пептидов преимущественно вазопрессиновыми рецепторами клеток экскреторных органов и артериол. Наличие в том же положении изолейцина обусловливает наибольшее сродство гормона к окситоциновым рецепторам клеток миометрия (гладкомышечного слоя матки) и миоэпите- лиальных образований молочных желез. Однако оба типа кольцевой части все же могут связываться, хотя и с разной степенью интенсивности, с обоими типами рецепторов и конкурировать друг с другом за связывание. По- видимому, структура всей 1-6-петли нейрогипофизарных пептидов ответственна за принципиальную возможность гормон-рецепторного взаимодействия, а остатки в 3-й позиции петли определяют силу данного взаимодействия с тем или иным типом рецепторов и специфику эффекта. Роль актона , по существующим представлениям, выполняют боковая цепь и остаток тирозина во 2-м положении.
В настоящее время основными направлениями развития исследований белково - пептидных гормонов являются:
1) изучение тонкой структурно - функциональной организации генов и мРНК, кодирующих белково - пептидные гормоны млекопитающих, выявление основных регуляторных элементов этих генов, анализ их структуры и механизмов тканеспецифичной мультигормональной (мультифакторной) регуляции.
2) изучение генов и мРНК, кодирующих факторы белковой природы, которые регулируют экспрессию данных белково - пептидных гормонов млекопитающих, анализа их структуры и механизмов взаимодействия с регуляторными участками промоторных областей генов белково - пептидных гормонов;
3) исследование структурно - функциональной организации самих белково - пептидных гормонов, выявление функциональной значимости отдельных аминокислотных доменов, выяснение закономерных взаимосвязей между аминокислотной последовательностью и функциональной активностью; 4) выяснение молекулярных механизмов действия белково -пептидных гормонов в клетке - мишени, расшифровка цепи молекулярных сигналов, реализующих воздействие пептидного гормонва с поверхности мембранного рецептора клетки на ген, локализованный в хромосоме.
ЧАСТЬ В
1. Витамин А (ретинол)
Правильное химическое название витамина А — ретинол. Он обнаружен в продуктах животного происхождения. Пигмент каротин (оранжевого цвета), присутствующий в моркови, и схожие пигменты, называемые каротинами, часто встречающиеся в растениях, могут преобразовываться в витамин А в процессе пищеварения. Структура каротинов и витамина А особенно хорошо адаптирована для поглощения света, как в растениях в форме каротинов, так и у животных, у которых витамин А превращается в светопоглощающую молекулу ретиналь. Три группы животных, у которых имеются глаза (моллюски, членистоногие и позвоночные), используют ретиналь в качестве светопоглощающей части фоторецепторных молекул. Свет вызывает довольно большие изменения в структуре ретиналя, достаточные для генерации нервного импульса.
Помимо этого витамин А играет важную роль в сохранении нормального состояния кожи и других эпителиальных (поверхностных) тканей, а маленьким детям он требуется для нормального роста.
Гиповитаминоз А.
Дефицит витамина А приводит к нарушению темновой адаптации (иными словами, нарушает работу палочек, реагирующих на интенсивность света). На первых порах возникает так называемая «куриная слепота», когда человек не видит в сумерках. Это состояние развивается в условиях недостатка ретиналя в палочках. В конечном счете палочки разрушаются. Одновременно возникает сухость конъюнктивы и роговицы (ксерофтальмия; xeros — сухой, ophthalmos — глаз), и нарушаются их функции. На роговице появляются язвы (кератомаляция) и как следствие этого — слепота.
При дефиците витамина А у детей замедляется их рост. Продолжительный дефицит этого витамина может привести к смерти. Подобная картина все еще характерна для некоторых развивающихся стран, где из-за недостатка витамина А у детей нередко наступает слепота. Около 3 млн. детей в возрасте до 10 лет в этих странах слепы именно по этой причине. У человека витамин А хранится в печени, в которой при условии полноценной диеты существует примерно двухлетний его запас. По рекомендации британских диетологов среднесуточное потребление этого витамина должно в два раза превышать потребности организма в нем.
Гипервитаминоз А
Известны редкие случаи отравления, явившиеся результатом приема избыточного количества витамина А. Обычно это бывает при длительном употреблении витаминных препаратов. Проявляется гипервитаминоз А в ломкости костей, выпадении волос, двоении в глазах, рвоте и других осложнениях. Прием большого количества (более 3300 мкг в день) витамина А во время беременности может привести к возникновению врожденных пороков развития у детей. В Соединенном Королевстве беременным женщинам не рекомендуют принимать витамины, содержащие витамин А, без консультации с врачом.
Суточное потребление при регулярном приеме не должно превышать 6000 мкг для подростков, 7500 мкг — для взрослых женщин и 9000 мкг для взрослых мужчин.
2. Витамин Д
Витамины группы D образуются под действием ультрафиолета в тканях животных и растений из стеринов.
витамин D2 - эргокальциферол; выделен из дрожжей, его провитамином является эргостерин;
витамин D3 - холекальциферол; выделен из тканей животных, его провитамин - 7-дегидрохолестерин;
витамин D4 - 22, 23-дигидро-эргокальциферол;
витамин D5 - 24-этилхолекальциферол (ситокальциферол); выделен из масел пшеницы;
витамин D6 - 22-дигидроэтилкальциферол (стигма-кальциферол). Действие
Основная функция витамина D - обеспечение нормального роста и развития костей, предупреждение рахита и остеопороза. Он регулирует минеральный обмен и способствует отложению кальция в костной ткани и дентине, таким образом, препятствуя остеомаляции (размягчению) костей.
Поступая в организм, витамин D всасывается в проксимальном отделе тонкого кишечника, причем обязательно в присутствии желчи. Часть его абсорбируется в средних отделах тонкой кишки, незначительная часть - в подвздошной. После всасывания кальциферол обнаруживается в составе хиломикронов в свободном виде и лишь частично в форме эфира. Биодоступность составляет 60-90%.
Витамин D влияет на общий обмен веществ при метаболизме Ca2+ и фосфата (НРО2-4). Прежде всего, он стимулирует всасывание из кишечника кальция, фосфатов и магния. Важным эффектом витамина при этом процессе является повышение проницаемости эпителия кишечника для Ca2+ и Р.
Витамин D является уникальным - это единственный витамин, действующий и как витамин, и как гормон. Как витамин он поддерживает уровень неорганического Р и Са в плазме крови выше порогового значения и повышает всасывание Са в тонкой кишке.
В качестве гормона действует активный метаболит витамина D - 1,25-диоксихолекациферол, образующийся в почках. Он оказывает влияние на клетки кишечника, почек и мышц: в кишечнике стимулирует выработку белка-носителя, необходимого для транспорта кальция, а в почках и мышцах усиливает реабсорбцию Ca++.
Витамин D3 влияет на ядра клеток-мишеней и стимулирует транскрипцию ДНК и РНК, что сопровождается усилением синтеза специфических протеидов.
Однако роль витамина D не ограничивается защитой костей, от него зависит восприимчивость организма к кожным заболеваниям, болезням сердца и раку. В географических областях, где пища бедна витамином D, повышена заболеваемость атеросклерозом, артритами, диабетом, особенно юношеским.
Он предупреждает слабость мускулов, повышает иммунитет (уровень витамина D в крови служит одним из критериев оценки ожидаемой продолжительной жизни больных СПИДом), необходим для функционирования щитовидной железы и нормальной свертываемости крови.
Так, при наружном применении витамина D3 уменьшается характерная для псориаза чешуйчатость кожи.
Есть данные, что, улучшая усвоение кальция и магния, витамин D помогает организму восстанавливать защитные оболочки, окружающие нервы, поэтому он включается в комплексную терапию рассеянного склероза.
Витамин D3 участвует в регуляции артериального давления (в частности, при гипертонии у беременных) и сердцебиения.
Витамин D препятствует росту раковых и клеток, что делает его эффективным в профилактике и лечении рака груди, яичников, предстательной железы, головного мозга, а также лейкимии.
3. Витамин E жирорастворимый, накапливается в печени, жировых тканях, в сердце, мышцах, яичках, матке, крови, надпочечниках и гипофизе. Токоферол объединяет ряд ненасыщенных спиртов-токоферолов, из которых наиболее активным является альфа-токоферол.
Действие: витамин Е улучшает циркуляцию крови, необходим для регенерации тканей, полезен при предменструальном синдроме и лечении фиброзных заболевания груди. Он обеспечивает нормальную свертываемость крови и заживление, снижает возможность образования шрамов от некоторых ран, снижает кровяное давление, способствует предупреждению катаракт, улучшает атлетические достижения, снимает судороги ног, поддерживает здоровье нервов и мускулов, укрепляя стенки капилляров, предотвращает анемию. В качестве антиоксиданта витамин Е защищает клетки от повреждения, замедляя окисление липидов (жиров) и формирование свободных радикалов. Он защищает другие растворимые жирами витамины от разрушения кислородом, способствует усвоению витамина А и защищает его от кислорода. Витамин Е замедляет старение, может предотвращать появление старческой пигментации. Витамин Е участвует в формировании коллагеновых и эластичных волокон межклеточного вещества. Токоферол предотвращает повышенную свертываемость крови, благоприятно влияет на периферическое кровообращение, участвует в биосинтезе гема и белков, пролиферации клеток, образовании гонадотропинов, развитии плаценты. Витамина Е способен облегчать болезнь Альцгеймера и диабет, а также улучшать иммунную функцию организма. Витамин E играет профилактическую роль - он не может восстановить уже имеющееся повреждение. Эффективность витамина Е повышается в присутствии других питательных веществ-антиоксидантов. Его противораковое защитное действие особенно заметно повышает витамин С.
Взаимодействие: если вы на диете, с высоким содержанием полиненасыщенных масел, вам может понадобиться витамин Е. Неограниченное железо разрушает витамин Е, так что их не следует принимать вместе. Глюконат, пептонат, цитрат или фумерат железа не разрушают витамин Е.
Симптомы дефицита витамина E: первым и наиболее ранним признаком, проявляющимся довольно быстро при недостаточном поступлении с пищей витамина Е и избыточном поступлении ненасыщенных жирных кислот, является мышечная дистрофия. Дистрофия скелетных мышц считается наиболее универсальным проявлением авитаминоза Е. Наиболее тяжелые поражения отмечаются в диафрагме. Мышечные волокна подвергаются распаду, а в некротизированных волокнах откладываются соли кальция. В печени при авитаминозе Е описаны некрозы, жировая дистрофия, расширение синусоидов, уменьшение содержания гликогена. Недостаточность также может провоцировать сокращение длительности жизни красных кровяных клеток (эритроцитов). Исследования на животных доказывают, что при дефиците витамина Е могут также страдать сердечная мышца и репродуктивные функции организма.
Польза: помогает выглядеть моложе, замедляя старение клеток; защищает легкие от загрязненного воздуха; защищает клеточные структуры от разрушения свободными радикалами (действует как антиоксидант); предупреждает появление и растворяет кровяные тромбы; ускоряет заживление ожогов; действуя как мочегонное средство, может понижать кровяное давление; предохраняет от выкидышей; поддерживает иммунитет; обеспечивает нормальное функционирование мускулатуры.
Где содержится: растительные масла: из пшеничных зародышей, оливковое, подсолнечное, хлопковое, кукурузное, соевое; семечки яблок, миндаль, арахис, зеленые листовые овощи, злаковые, бобовые, яичный желток, печень, молоко, овсянка, соя, пшеница и ее проростки, одуванчик, люцерна, льняное семя, крапива, овес, лист малины, плоды шиповника.
Аптека корпорации «Тяньши»: Витамин Е содержится в препарате Вэйкан, который чрезвычайно богат витаминами и минералами. Вэйкан производства «Тяньши» применяют для лечения сердечно-сосудистых заболеваний, таких как инфаркт, инсульт, стенокардия, гипертоническая болезнь, варикозное расширение вен, атеросклероз.
Наилучший вариант для сохранения красоты — употреблять добавки целебных трав китайской медицины, содержащие витамины Е и С. Одной из таких добавок является Лянь Янь, которая содержит оба этих витамина в необходимом количестве. Принимая витамин С 3 раза в день по 400мг и витамин Е 3 раза в день по 200мг, 71,4% пациентов избавились от коричневых пятен за 3 месяца!
Также уникальным источником Витамина Е является - Омега-3 корпорации «Тяньши». Основные компоненты препарата: полиен, облепиховое масло, витамин Е — 1 капсула содержит 0,9г Омега-3 полиненасыщенных жирных кислот (50% суточной дозы) и 1,5мг витамина Е (10% суточной дозы).
4. Витамин РР (ниацин, никотиновая кислота)
Ниацин входит в состав ферментов, участвующих в клеточном дыхании и обмене белков, регулирующих высшую нервную деятельность и функции органов пищеварения. Используется для профилактики и лечения пеллагры, заболеваний желудочно-кишечного тракта, вяло заживающих ран и язв, атеросклероза.
При передозировке или при повышенной чувствительности могут возникать покраснение лица и верхней половины туловища, головокружение, чувство прилива к голове, крапивница. При быстром внутривенном введении возможно сильное понижение артериального давления.
Основными источниками витамина РР служат мясо, печень, почки, яйца, молоко. Содержится витамин PP также в хлебных изделиях из муки грубого помола, в крупах (особенно гречневой), бобовых, присутствует в грибах.
Суточная потребность в витамине РР взрослого человека составляет 14-18 мг; беременных и кормящих - 19-21 мг; детей первого года жизни - 5-7 мг. Витамин РР может синтезироваться в организме человека из незаменимой аминокислоты триптофана, входящей в состав белков.
Витамин РР относительно устойчив к тепловой обработке. Необходимо учитывать, что в зерновых продуктах, особенно в кукурузе, большая часть ниацина находится в связанной форме (ниацитин), эта часть витамина становится доступной только после интенсивной тепловой обработки. В бобовых и продуктах животного происхождения связанная форма отсутствует.
5. КОФЕРМЕНТЫ
Коферменты (синоним коэнзимы) — органические соединения небелковой природы, необходимые для осуществления каталитического действия многих ферментов. Соединяясь с белковой частью молекулы фермента — апоферментом, К. образуют каталитически активный комплекс — холофермент. Прочно связанные с белками К. называются простетическими группами. Многие К. легко отделяются от ферментного белка и служат переносчиками электронов, отдельных атомов или групп атомов субстрата, превращение которого катализирует данный фермент, т.е. функционируют в качестве промежуточных акцепторов. К. могут участвовать в активировании молекул субстратов, образуя с ними реакционно-способные соединения, которые затем подвергаются ферментативному превращению. Некоторые метаболиты, выступающие в ферментативных реакциях как обычные субстраты, в определенных условиях могут выполнять роль К. Многие К. являются производными витаминов, поэтому нарушение обмена веществ при витаминной недостаточности опосредовано через понижение активности определенных ферментов.
Коферменты, как правило, термостабильны, разнообразны по химическому строению и механизму действия. Наиболее распространенную группу составляют соединения нуклеотидной природы, а также К., содержащие остатки фосфорной кислоты. Адениловые нуклеотиды наряду с их ключевой ролью в обмене энергии (см. Макроэргические соединения, Обмен веществ и энергии) в качестве К. участвуют в реакциях переноса и активации орто- и пирофосфатных остатков, аминоацильных групп, остатков неорганических кислот. В группу адениловых нуклеотидов входят аденозинфосфорные кислоты — нуклеотиды, содержащие аденин, рибозу и остатки фосфорной кислоты (АДФ и АМФ). В подобных реакциях могут участвовать в качестве К. также производные инозин-5'-фосфорной и гуанозин-5'-фосфорной кислот. Гуаниловые рибонуклеотиды (гуанозин-5'-моно-, ди- и трифосфорные кислоты) выполняют роль К. в реакциях переноса сукцинильной группы, при биосинтезе рибонуклеопротеинов в микросомах, биосинтезе адениловой кислоты из инозиновой кислоты и др. Цитидиловые рибонуклеотиды (цитидил-5'-моно-, ди- и трифосфорные кислоты) играют роль К. при биосинтезе фосфолипидов, участвуя в переносе остатков, образующих полярные «головки» молекул фосфолипидов (0-фосфоэтанолхолина, 0-фосфоэтаноламина и др.). Уридиловые рибонуклеотиды (уридин-5'-моно, ди- и трифосфорные кислоты) участвуют в качестве К. в процессах трансгликозилирования (переноса остатков простых сахаров и их производных) при биосинтезе ди- и полисахаридов, гликозаминогликанов и реакциях взаимопревращения сахаров.
К важнейшим К. нуклеотидной природы относятся никотинамидные К. никотинамидадениндинуклеотид (НАД, NAD+; синоним: дифосфопиридиннуклеотид, ДПН; устаревшее кофермент I, Kol, козимаза, кодегидр (оген)аза I] и его фосфорилированное производное никотинамидадениндинуклеотидфосфат [НАДФ, NADP+; синоним: трифосфопиридиннуклеотид, ТПН; устаревшее кофермент II, KoII, фосфокозимаза, фосфокодегидр(оген)аза]. Никотинамидные К. входят в состав ряда дегидрогеназ — катализаторов ключевых окислительно-восстановительных реакций энергетического и пластического обмена. Молекула НАД представляет собой динуклеотид, построенный из аденинрибонуклеотида и никотинамидрибонуклеотида (последний отвечает за проявление каталитической активности НАД), связанных фосфоангидридным мостиком, а НАДФ имеет третий остаток фосфорной кислоты в положении 2' рибозы аденилового нуклеотида. Способность НАД и НАДФ переносить электроны и протоны от окисляемого субстрата к другому акцептору обеспечивает выполнение этими К. важной биологической функции в процессе клеточного дыхания (см. Дыхание тканевое). Окислительно-восстановительные реакции, протекающие с участием никотинамидных К., могут быть изображены в виде общего уравнения; АН2 + НАД(Ф)+ = А + НАД(Ф)Н + Н+, где АН2 восстановленная форма субстрата. А — окисленная форме субстрата. Эти реакции состоят в обратимом переносе двух восстановительных эквивалента от субстрата к окисленному никотинамидному К. Один восстановительный эквивалент присутствует в восстановленном К. в виде атома водорода, другой — в виде электроне, при этом катион второго атома водорода переходит в среду а виде свободного Н+. Обнаружено около 350 НАД(Ф)-зависимых дегидрогеназ, как правило, специфичных в отношении либо НАД, либо НАДФ. Обычно связь никотинамидных и других нуклеотидных К. с белками легко диссоциирует. НАД-зависимые дегидрогеназы участвуют преимущественно в процессах катаболизма (например, в клеточном дыхании), в НАДФ-зависимые — главным образом в анаболических процессах (восстановительных биосинтетических реакциях). Содержание никотинамидных К., соотношение между их окисленными и восстановленными формами (НАДН и НАДФН), к также величию отношения НАД/НАДФ являются показателями активности метаболических процессов в ткани, характеризуют ее функциональное состояние. В организме НАД и НАДФ синтезируются из никотиновой кислоты (ниацина, или витамина РР) или никотинамида, поэтому недостаточность ниацина ведет к нарушению биосинтеза никотинамидных коферментов.
5. Флавиновые коферменты ФМН (FMN) и ФАД (FAD) (2, см. с. 86) найдены в дегидрогеназах, оксидазах и монооксигеназах. Обычно оба соединения ковалентно связаны с ферментами. Активной группой обоих коферментов является флавин (изоаллоксазин), имеющий сопряженную систему из трех колец, которая может при восстановлении принимать два электрона и два протона. В ФМН к флавину присоединен фосфорилированный полиол рибит. ФАД состоит из ФМН, связанного с АМФ. Оба соединения являются функционально близкими коферментами.
8.КоА Ацетил коэнзим А или ацетил-КоА – важное соединение в обмене веществ, используемое во многих биохимических реакциях. Его главная функция – доставлять атомы углерода с ацетил-группой в цикл трикарбоновых кислот, чтобы те были окислены с выделением энергии. По своей химической структуре ацетил-КоА – тиоэфир между коферментом А (тиолом) и уксусной кислотой (носителем ацильной группы). Ацетил-КоА образуется во время второго шага кислородного клеточного дыхания, декарбоксилирования пирувата, который происходит в матриксе митохондрии. Ацетил-КоА затем поступает в цикл трикарбоновых кислот.
Ацетил-КоА – важный компонент биологического синтеза нейротрансмиттера ацетилхолина. Холин, в соединении с ацетил-КоА, катализируется ферментом холинацетилтрансферазой, чтобы образовать ацетилхолин и коэнзим А.
9. Пантотеновая кислота, пантотенат (лекарственная форма — пантотенат кальция, также витамин B3, B5. Пантотеновая кислота по химической природе является дипептидом и состоит из остатков аминокислоты β-аланина и пантоевой кислоты.
Пантотеновая кислота получила свое название от греческого «пантотен», что означает «всюду», из-за чрезвычайно широкого её распространения. Пантотеновая кислота, попадая в организм, превращается в пантетин, который входит в состав кофермента А, который играет важную роль в процессах окисления и ацетилирования. Кофермент А — одно из немногих веществ в организме, участвующее в метаболизме и белков, и жиров, и углеводов.
Пантонетовая кислота требуется для обмена жиров, углеводов, аминокислот, синтеза жизненно важных жирных кислот, холестерина, гистамина, ацетилхолина, гемоглобина. Пантотеновая кислота чувствительна к нагреванию, при термической обработке теряется почти 50 % витамина. В клетках животных и растений пантотеновая кислота входит в состав кофермента A (KoA), принимающего участие в важнейших реакциях обмена веществ.
Важнейшим свойством витамин пантотеновой кислоты является ее способность стимулировать производство гормонов надпочечников — глюкокортикоидов, что делает его мощным средством для лечения таких заболеваний как артрит, колит, аллергия и болезни сердца. Витамин играет важную роль в формировании антител, способствует усвоению других витаминов, а также принимает участие в синтезе нейротрансмиттеров.
Пантотеновая кислота участвует в метаболизме жирных кислот. Она нормализует липидный обмен и активирует окислительно-восстановительные процессы в организме.
Пантотеновая кислота оказывает значительное гиполипидемическое действие, обусловленное, по-видимому, ингибированием биосинтеза основных классов липидов, формирующих в печени липопротеины низкой и очень низкой плотности.
10. Тиами́н (витамин B1; старое название — аневрин) — водорастворимый витамин, соединение, отвечающее формуле C12H17N4OS. Бесцветное кристаллическое вещество, хорошо растворимое в воде, нерастворимое в спирте. Разрушается при нагревании.
Известный как витамин B1 тиамин играет важную роль в процессах метаболизма углеводов и жиров. Вещество необходимо для нормального протекания процессов роста и развития и помогает поддерживать надлежащую работу сердца, нервной и пищеварительной систем. Тиамин, являясь водорастворимым соединением, не запасается в организме и не обладает токсическими свойствами. В природе тиамин синтезируется растениями и многими микроорганизмами. Животные и человек не могут синтезировать тиамин и получают его вместе с пищей. В тиамине нуждаются все животные за исключением жвачных, так как бактерии в их кишечнике синтезируют достаточное количество витамина.
Всасываясь из кишечника, тиамин фосфорилируется и превращается в тиаминпирофосфат.
Тиаминпирофосфат (ТПФ) — активная форма тиамина — является коферментом пируватдекарбоксилазного и α-кетоглутаратдекарбоксилазного комплексов, а также транскетолазы. Первые два фермента участвуют в метаболизме углеводов, транскетолаза функционирует в пентозофосфатном пути, участвуя в переносе гликоальдегидного радикала между кето- и альдосахарами. ТПФ синтезируется ферментом тиаминпирофосфокиназой, главным образом в печени и в ткани мозга. Реакция требует присутствия свободного тиамина, ионов Mg2+ и АТФ. Также ТПФ выступает коферментом дегидрогеназы γ-оксиглутаровой кислоты и пируватдекарбоксилазы клеток дрожжей.
Другими производными тиамина являются:
Тиаминтрифосфат, обнаружен у бактерий, грибов, растений и животных[1], у E. coli играет роль сигнальной молекулы при ответе на аминокислотное голодание[2].
Аденозинтиаминдифосфат — накапливается у E. coli в результате углеродного голодания[3].
Аденозинтиаминтрифосфат — присутствует в небольших количествах в печени позвоночных, функция его неизвестна[
11. Витамин B6 — собирательное название производных 3-гидрокси-2-метилпиридинов, обладающих биологической активностью пиридоксина[1] — собственно пиридоксин, пиридоксаль, пиридоксамин, а также их фосфаты, среди которых наиболее важен пиридоксальфосфат.
Различные формы в организме человека превращаются в пиридоксальфосфат — кофактор ферментов, которые катализируют декарбоксилирование и трансаминирование аминокислот.
Витамин B6 (пиридоксин, пиридоксаль, пиридоксамин) содержится во многих продуктах. Особенно много его содержится в зерновых ростках, в грецких орехах и фундуке, в шпинате, картофеле и батате, моркови, цветной и белокочанной капусте, помидорах, клубнике, черешне, апельсинах и лимонах, авокадо. Также он содержится в мясных и молочных продуктах, рыбе, яйцах, крупах и бобовых. Витамин B6 (пиридоксин, пиридоксаль, пиридоксамин) синтезируется в организме кишечной микрофлорой. Пиридоксин
Основная статья: Пиридоксин
Пиридоксин
Пиридоксин представляет собой бесцветные кристаллы, растворимые в воде. В большинстве съедобных растений пиридоксина нет, или он встречается в незначительных количествах. Но существуют растения, например, питайя, исключительно богатые пиридоксином. Пиридоксин синтезируется некоторыми бактериями. Также он содержится в мясных и молочных продуктах, но он менее устойчив к высоким температурам, чем другие формы витамина B, поэтому в варёных и жареных мясных продуктах его мало. Вегетарианцы могут получить пиридоксин из кожицы некоторых овощей, в которых имеются остатки почвы, например, картофеля, моркови. Регистрационный номер CAS 65-23-6, для соли (гидрохлорида) 58-56-0. Имеются данные, что избыточные дозы пиридоксина могут привести к токсическому эффекту.
Пиридоксаль
Брутто-формула пиридоксаля C8H9NO3. Пиридоксаль отличается от пиридоксина тем, что вместо одной из трёх гидроксогрупп к пиридиновому кольцу присоединена карбонильная группа, так что пиридоксаль является одновременно и альдегидом. Молярная масса 0.16716 кг/моль. Представляет собой кристаллический порошок, плавящийся при 165 °C. Пиридоксаль содержится в зелёных частях некоторых растений, в цветной и белокочанной капусте, в мясе. Регистрационный номер CAS 66-72-8, для соли (гидрохлорида) 65-22-5.
Пиридоксамин
Пиридоксамин отличается от пиридоксина тем, что вместо одной из трёх гидроксогрупп к пиридиновому кольцу присоединена аминогруппа. Пиридоксамин содержится в мясе животных. Также пиридоксамин сдержится в некоторых пищевых добавках. Также в некоторых добавках содержится его соль — дигидрохлорид пиридоксамина. Однако в США в 2009 году FDA постановила, что отныне пиридоксамин считается лекарством, и его нельзя вносить в пищевые добавки. Регистрационный номер CAS 85-87-0 Пиридоксальфосфат
Пиридоксальфосфат образуется в организме человека из любого из трёх вышеуказанных веществ. Пиридоксальфосфат может быть получен через пищу в готовом виде из мяса животных. Он также может быть синтезирован искусственно химическим путём. Регистрационный номер CAS 54-47-7.
Пиридоксальфосфат является коферментом большого числа ферментов азотистого обмена (трансаминаз, декарбоксилаз аминокислот) и других ферментов.
Пиридоксальфосфат:
принимает участие в образовании эритроцитов;
участвует в процессах усвоения нервными клетками глюкозы;
необходим для белкового обмена и трансаминирования аминокислот;
принимает участие в обмене жиров;
оказывает гипохолестеринемический эффект;
оказывает липотропный эффект, достаточное количество пиридоксина необходимо для нормального функционирования печени.
14. Вещества, которые оказывают влияние на активность ферментов, называют эффекторами. Это могут быть ингибиторы – соединения, тормозящие каталитический процесс, или активаторы – вещества, которые этот процесс ускоряют. Учение об ингибиторах ферментов имеет большое теоретическое и практическое значение для фармакологии и токсикологии. Многие лекарственные препараты являются ингибиторами ферментов. Например, ингибиторы амилаз успешно применяются для лечения заболеваний, связанных с повышенной активностью этих ферментов – диабета, ожирения, кариеса. Используемые в военном деле нервно-паралитические газы представляют собой специфические ингибиторы ферментов. В научных исследованиях специфические ингибиторы используются для изучения механизма действия ферментов, строения их активного центра. Например, многие из промежуточных продуктов гликолиза и дрожжевого брожения были открыты благодаря использованию ингибиторов, блокирующих последовательные стадии процесса. В результате такого блокирования соответствующие промежуточные продукты накапливались в количествах, достаточных для их выделения и идентификации.
По типу действия ингибиторы можно разделить на обратимые и необратимые. Удаление обратимых ингибиторов из системы (диализом, гельфильтрацией и др.) восстанавливает каталитическую активность фермента.
Обратимо действуют эффекторы:
1. Близкие аналоги субстрата, которые связываются активным центром фермента, но
не подвергаются превращению. Занимая активный центр, они препятствуют связыванию истинного субстрата, конкурируя с ним, и поэтому называются конкурентными ингибиторами.
2. Кофакторы ферментов, без которых апофермент вообще не обладает активностью.
Постепенное добавление их приводит к появлению активности, которая затем повышается до определенного предела, соответствующего полному насыщению.
3. Вещества, которые взаимодействуют с дополнительными, регуляторными центрами, несовпадающими с активным центром. Тем не менее, это взаимодействие изменяет конформацию в районе активного центра и влияет на кинетику ферментативного процесса. Такие соединения называются аллостерическими эффекторами. Они имеют важное биологическое значение, так как с их помощью осуществляется один из механизмов регуляции каталитической активности.
Необратимую инактивацию вызывают соединения (найденные в живой природе или полученные путем синтеза), которые вступают в химическую реакцию с участком фермента, важным для проявления каталитической активности. Такие соединения, специфически реагирующие с определенными группами в молекулах ферментов (групп-специфические реагенты), используют для идентификации функциональных групп активного центра (метод химической модификации).
С этой целью широко используются соединения, блокирующие SH-группы (иодацетамид, n-хлормеркурибензоат и др.), окисляющие остатки триптофана в кислой среде (N-бромсукцинимид), ацетилирующие остатки тирозина (N-ацетилимидазол), связывающие металлы (азид натрия) и т.д.
15. ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ
ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ (протеазы), ферменты класса гидролаз, катализирующие гидролиз (протео-лиз) пептидных связей. Место расщепления пептидной связи в полипептидной цепи определяется позиционной и субстратной специфичностью фермента и пространств. структурой гидролизуемого субстрата (белка или пептида).
Различают экзопептидазы, расщепляющие связи вблизи С- или N-конца цепи (соотв. карбоксипептидазы и аминопеп-тидазы)и эндопептидазы (протеиназы), гидролизующие связи, удаленные от концевых остатков (напр., трипсин). Лишь ограниченное число протеолитических ферментов обладает строгой субстратной специфичностью. К ним относят, напр., ренин, гидроли-зующий связь между остатками лейцина в положениях 10 и 11 в ангиотензиногене (предшественник ангиотензина пептида, участвующего в регуляции кровяного давления), или энтеропептидазу отщепляющую N-концевой гексапептид в трипсиногене (предшественник трипсина). Специфичность большинства протеолитических ферментов определяется в осн. структурой аминокислотного остатка, расположенного рядом с расщепляемой связью. Ферменты трипсинового типа катализируют гидролиз связей, образованных карбоксильной группой основных аминокислот (остатками лизина и аргинина). Для мн. ферментов (химотрипсин, пепсин, субтилизины и др.) важно наличие вблизи расщепляемой связи объемистых гидрофобных остатков (фенилаланина, тирозина, триптофана и лейцина). Протеолитические ферменты типа эластазы (фермент поджелудочной железы) гидролизуют связи, образованные аминокислотными остатками с небольшой боковой группой (напр., остатками аланина и серина). Место расщепления зависит от расположения пептидной связи в пространств. структуре субстрата-легче всего гидролизуются связи на р-изгибах цепи, к-рые расположены на пов-сти молекулы. Углеводные цепи в гликопротеинах могут препятствовать доступу фермента к данной связи.
Многие протеолитические ферменты прочно ассоциированы с клеточными мембранами и поэтому действуют только на определенные белки (т. наз. компартментализация). К ним относят, напр., сигнальные протеазы, участвующие в транспорте белков во внеклеточное пространство. В зависимости от локализации фермента протеолиз происходит при разл. рН. Так, протеолитические ферменты желудка (напр., пепсин, гастриксин) функционируют при рН 1,5-2, лизосомные ферменты-при рН 4-5, а протеолитические ферменты сыворотки крови, тонкого кишечника и др.-при нейтральных или слабощелочных значениях рН. Нек-рые протеолитические ферменты используют в качестве кофактора ионы металлов-Са2+, Mg2+ и др.
Дефектные и чужеродные белки деградируют в клетке при участии АТФ-зависимой системы протеолиза. У эукариот (все организмы, кроме бактерий и синезеленых водорослей) эта система включает низкомол. белок убикитин, образующий с белками-субстратами конъюгат, и протеазы, расщепляющие этот конъюгат.
Протеолитические ферменты играют важную роль во мн. процессах, происходящих в организме, напр. при оплодотворении, биосинтезе белка, свертывании крови и фибринолизе, иммунном ответе (активации системы комплемента), гормональной регуляции. Во мн. этих случаях фермент расщепляет в субстрате лишь одну или неск. связей (ограниченный протеолиз). Активность протеолитических ферментов регулируется на посттрансляц. стадии путем активации их неактивных предшественников (зи-могенов), а также действием прир. ингибиторов ферментов (a2-макроглобулина, a1антитрипсина, секреторного панк-реатич. ингибитора и др.). Нарушения механизмов регуляции активности протеолитических ферментов-причина мн. тяжелых заболеваний (мышечной дистрофии, аутоиммунных заболеваний, эмфиземы легких, панкреатитов и др.).
Протеолитические ферменты применяют в медицине, напр. для коррекции нарушений пищеварения, заживления ран и ожогов и др. Их также используют для получения смесей аминокислот, применяемых для парэнтерального питания, в произ-ве гормональных препаратов и нек-рых антибиотиков, в пищ. и кожевенной пром-сти, произ-ве моющих ср-в.
Регуляция активности ферментов путем ковалентной модификации. Приведите примеры.
Регуляция каталитической активности ферментов путём фосфорилирования/дефосфорилирования
В биологических системах часто встречается механизм регуляции активности ферментов с помощью ковалентной модификации аминокислотных остатков. Быстрый и широко распространённый способ химической модификации ферментов - фосфорилирование/дефосфорилирование. Модификации подвергаются ОН-группы фермента. Фос-форилирование осуществляется ферментами протеинкиназами, а дефосфорилирование
- фосфопротеинфосфатазами. Присоединение остатка фосфорной кислоты приводит к изменению конформации активного центра и его каталитической активности. При этом результат может быть двояким: одни ферменты при фосфорилировании активируются, другие, напротив, становятся менее активными (рис. 2-33).
Изменение активности фермента, вызванное фосфорилированием, обратимо. Отщепление остатка фосфорной кислоты осуществляется ферментами фосфопротеинфосфатазами. Активность протеинкиназ и фосфопротеинфосфатаз регулируется гормонами, что позволяет быстро изменять активность ключевых ферментов метаболических путей в зависимости от условий внешней среды. Антагонистичные по функции гормоны противоположным образом влияют на фосфо-рилирование/дефосфорилирование ферментов, вызывая противоположные эффекты изменения метаболизма клетки.
Например, под действием глюкагона (в период между приёмами пищи) в клетках происходит уменьшение синтеза энергетического материала - жира, гликогена и усиление его распада (мобилизация), вызванного фосфо-рилированием ключевых ферментов этих процессов. А под действием инсулина (во время пищеварения), наоборот, активируется синтез гликогена и ингибируется его распад, так как взаимодействие инсулина с рецептором активирует сигнальный путь, приводящий к дефосфорилированию тех же ключевых ферментов.
Гормональная регуляция активности ферментов с участием вторичных посредников. (??)
Аденилатциклазная система. Аденилатциклаза и протеинкиназа катализируют взаимосвязанные реакции, которые составляют единую регуляторную систему.
С помощью этой системы в клетку передаются сигналы из внеклеточной среды, и в нужном направлении изменяется метаболизм клетки. Внеклеточным вестником сигнала могут быть разные молекулы, в том числе и гормоны. Эти молекулы не проникают внутрь клетки, но «узнаются» мембранными рецепторами. При активации аденилатциклазы происходят следующие этапы:
-изменение конформации рецептора после присоединения к нему сигнальной молекулы и увеличение его сродства к регуляторному G-белку. В результате образуется комплекс рецептора и протомеров G-белка;
-образование этого комплекса приводит к изменению конформации a -протомера G-белка, который теряет сродство к GDP и происходит замена GDP на GTP. В результате комплекс протомеров G-белка распадается; a -протомер взаимодействует с аденилатциклазой, что ведет к изменению ее конформации и как следствие этого - активации;
-после этого аденилатциклаза катализирует синтез cAMP, который в свою очередь активирует cAMP-зависимую протеинкиназу. Активация последней связана с диссоциацией комплекса входящих в нее протомеров после присоединения cAMP. Протеинкиназа фосфорилирует соответствующие ферменты, изменяет их активность и, следовательно, скорость метаболизма в клетке.
Роль внутриклеточных посредников в проведении и усилении гормонального сигнала. (???)
Биосинтез РНК. Этапы транскрипции. Процессинг мРНК.
Транскрипция - первая стадия реализации генетической информации в клетке. В ходе процесса образуются молекулы мРНК, служащие матрицей для синтеза белков, а также транспортные, рибосомальные и другие виды молекул РНК, выполняющие структурные, адапторные и каталитические функции.
Транскрипция у эукариотов происходит в ядре. В основе механизма транскрипции лежит тот же структурный .принцип комплементарного спаривания оснований в молекуле РНК (G ≡ C, A=U и Т=А). ДНК служит только матрицей и в ходе транскрипции не изменяется. Рибонукле-озидтрифосфаты (ЦТФ, ГТФ, АТФ, УТФ) -субстраты и источники энергии, необходимые для протекания полимеразной реакции, образования 3',5'-фосфодиэфирной связи между рибонуклеозидмонофосфатами.
Синтез молекул РНК начинается в определённых последовательностях (сайтах) ДНК, которые называют промоторы, и завершается в терминирующих участках (сайты терминации). Участок ДНК, ограниченный промотором и сайтом терминации, представляет собой единицу транскрипции - транскриптон. У эукариотов в состав транскриптона, как правило, входит один ген, у прокариотов несколько. В каждом транскриптоне присутствует неинформативная зона; она содержит специфические последовательности нуклеотидов, с которыми взаимодействуют регуляторные транскрипционные факторы.
Транскрипционые факторы - белки, взаимодействующие с определёнными регуляторными сайтами и ускоряющие или замедляющие процесс транскрипции. Соотношение информативной и неинформативной частей в транскриптонах эукариотов составляет в среднем 1:9 (у прокариотов 9:1).
Соседние транскриптоны могут быть отделены друг от друга нетранскрибируемыми участками ДНК. Разделение ДНК на множество транскриптонов позволяет осуществлять с разной активностью индивидуальное считывание (транскрипцию) разных генов.
В каждом транскриптоне транскрибируется только одна из двух цепей ДНК, которая называется матричной, вторая, комплементарная ей цепь, называется кодирующей. Синтез цепи РНК идёт от 5'- к З'-концу, при этом матричная цепь ДНК всегда антипараллельна синтезируемой нуклеиновой кислоте.
Транскрипция не связана с фазами клеточного цикла; она может ускоряться и замедляться в зависимости от потребности клетки или организма в определённом белке.
РНК-полимеразы
Биосинтез РНК осуществляется ДНК-зависимыми РНК-полимеразами. В ядрах эукариотов обнаружены 3 специализированные РНК-полимеразы: РНК-полимераза I, синтезирующая пре-рРНК; РНК-полимераза II, ответственная за синтез пре-мРНК; РНК-полимераза III, синтезирующая пре-тРНК. РНК-полимеразы - олигомерные ферменты, состоящие из нескольких субъединиц - 2α, β, β', σ. Субъединица о (сигма) выполняет регуляторную функцию, это один из факторов инициации транскрипции, РНК-полимеразы I, II, III, узнающие разные промоторы, содержат разные по строению субъединицы σ.
Стадии транскрипции
В процессе транскрипции различают 3 стадии: инициацию, элонгацию и терминацию.
Инициация
Активация промотора происходит с помощью большого белка - ТАТА-фактора, называемого так потому, что он взаимодействует со специфической последовательностью нуклеотидов промотора - ТАТААА- (ТАТА-бокс) (рис. 4-29).
Присоединение ТАТА-фактора облегчает взаимодействие промотора с РНК-полимеразой. Факторы инициации вызывают изменение кон-формации РНК-полимеразы и обеспечивают раскручивание примерно одного витка спирали ДНК, т.е. образуется транскрипционная вилка,
Промоторные элементы - специфические последовательности нуклеотидов, характерные для любого промотора, связывающего РНК-полимеразу. Первый промоторный элемент - последовательность АТАТАА- (ТАТА-бокс) отделён от сайта начала транскрипции приблизительно на 25 пар нуклеотидов (п.н.). На расстоянии примерно 40 (иногда до 120) п.н. от него располагается последовательность GGCCAATC- (СААТ-бокс).
После того как синтезирован олигонуклеотид из 8-10 нуклеотидных остатков, σ-субъединица отделяется от РНК-полимеразы, а вместо неё к молекуле фермента присоединяются несколько факторов элонгации.
Элонгация
Факторы элонгации повышают активность РНК-полимеразы и облегчают расхождение цепей ДНК. Синтез молекулы РНК идёт от 5'- к З'-концу комплементарно матричной цепи ДНК. На стадии элонгации, в области транскрипционной вилки, одновременно разделены примерно 18 нуклеотидных пар ДНК. Растущий конец цепи РНК образует временную гибридную спираль, около 12 пар нуклеотидных остатков, с матричной цепью ДНК. По мере продвижения РНК-полимеразы по матрице в направлении от 3'- к 5'-концу впереди неё происходит расхождение, а позади - восстановление двойной спирали ДНК.
Терминация
Раскручивание двойной спирали ДНК в области сайта терминации делает его доступным для фактора терминации. Завершается синтез РНК в стадии транскрипции:1 - присоединение ТАТА-фактора к промотору. Чтобы промотор был узнан РНК-полимеразой, необходимо образование транскрипционного комплекса ТАТА-фактор/ТАТА-бокс (промотор). ТАТА-фактор остаётся связанным с ТАТА-боксом во время транскрипции, это облегчает использование промотора многими молекулами РНК-полимеразы; 2 - образование транскрипционной вилки; 3 - элонгация; 4.- терминация.
строго определенных участках матрицы - терминаторах (сайты терминации). Фактор терминации облегчает отделение первичного транскрипта (пре-мРНК), комплементарного матрице, и РНК-полимеразы от матрицы. РНК-полимераза может вступить в следующий цикл транскрипции после присоединения субъединицы σ.
Ковалентная модификация (процессинг) матричной РНК
Первичные транскрипты мРНК, прежде чем будут использованы в ходе синтеза белка, подвергаются ряду ковалентных модификаций. Эти модификации необходимы для функционирования мРНК в качестве матрицы.
Модификация 5'-конца
Модификации пре-мРНК начинаются на стадии элонгации. Когда длина первичного транскрипта достигает примерно 30 нуклеотидных остатков, происходит кэпирование его 5'-конца. Осуществляет кэпирование гуанилилтрансфераза. Фермент гидролизует макроэргическую связь в молекуле ГТФ и присоединяет нуклеотиддифосфатный остаток 5'-фосфатной группой к 5'-концу синтезированного фрагмента РНК с образованием 5', 5'-фосфодиэфирной связи. Последующее метилирование остатка гуанина в составе ГТФ с образованием N7-метилгуанозина завершает формирование кэпа (рис. 4-31).
Модифицированный 5'-конец обеспечивает инициацию трансляции, удлиняет время жизни мРНК, защищая её от действия 5'-экзонуклеаз в цитоплазме. Кэпирование необходимо для инициации синтеза белка, так как инициирующие триплеты AUG, GUG распознаются рибосомой только если присутствует кэп. Наличие кэпа также необходимо для работы сложной ферментной системы, обеспечивающей удаление нитронов.
Модификация 3'-конца
3'-Конец большинства транскриптов, синтезированных РНК-полимеразой II, также подвергается модификации, при которой специальным ферментом полиА-полимеразой формируется полиА-последовательность (полиА-"хвост"), состоящая из 100-200 остатков аде-ниловой кислоты.
Сигналом к началу полиаденилирования является последовательность -AAUAAA- на растущей цепи РНК. Фермент полиА-полимераза, проявляя экзонуклеазную активность, разрывает 3'-фосфоэфирную связь после появления в цепи РНК специфической последовательности -AAUAAA-. К 3'-концу в точке разрыва полиА-полимераза наращивает по-лиА-"хвост", Наличие полиА-последовательности на 3'-конце облегчает выход мРНК из ядра и замедляет её гидролиз в цитоплазме.
Ферменты, осуществляющие кэширование и полиаденилирование, избирательно связываются с РНК-полимеразой II, и в отсутствие полимеразы неактивны.
Транскрипция: биохимические механизмы и биологическая роль транскрипции.
Транскри́пция (от лат. transcriptio — переписывание) — процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех живых клетках. Другими словами, это перенос генетической информации с ДНК на РНК.
Транскрипция катализируется ферментом ДНК-зависимой РНК-полимеразой. Процесс синтеза РНК протекает в направлении от 5'- к 3'- концу, то есть по матричной цепи ДНК РНК-полимераза движется в направлении 3'->5'[1]
Транскрипция состоит из стадий инициации, элонгации и терминации.
Инициация
Активация промотора происходит с помощью большого белка - ТАТА-фактора, называемого так потому, что он взаимодействует со специфической последовательностью нуклеотидов промотора - ТАТААА- (ТАТА-бокс) (рис. 4-29).
Присоединение ТАТА-фактора облегчает взаимодействие промотора с РНК-полимеразой. Факторы инициации вызывают изменение кон-формации РНК-полимеразы и обеспечивают раскручивание примерно одного витка спирали ДНК, т.е. образуется транскрипционная вилка,
Промоторные элементы - специфические последовательности нуклеотидов, характерные для любого промотора, связывающего РНК-полимеразу. Первый промоторный элемент - последовательность АТАТАА- (ТАТА-бокс) отделён от сайта начала транскрипции приблизительно на 25 пар нуклеотидов (п.н.). На расстоянии примерно 40 (иногда до 120) п.н. от него располагается последовательность GGCCAATC- (СААТ-бокс).
После того как синтезирован олигонуклеотид из 8-10 нуклеотидных остатков, σ-субъединица отделяется от РНК-полимеразы, а вместо неё к молекуле фермента присоединяются несколько факторов элонгации.
Элонгация
Факторы элонгации повышают активность РНК-полимеразы и облегчают расхождение цепей ДНК. Синтез молекулы РНК идёт от 5'- к З'-концу комплементарно матричной цепи ДНК. На стадии элонгации, в области транскрипционной вилки, одновременно разделены примерно 18 нуклеотидных пар ДНК. Растущий конец цепи РНК образует временную гибридную спираль, около 12 пар нуклеотидных остатков, с матричной цепью ДНК. По мере продвижения РНК-полимеразы по матрице в направлении от 3'- к 5'-концу впереди неё происходит расхождение, а позади - восстановление двойной спирали ДНК.
Терминация
Раскручивание двойной спирали ДНК в области сайта терминации делает его доступным для фактора терминации. Завершается синтез РНК в стадии транскрипции:1 - присоединение ТАТА-фактора к промотору. Чтобы промотор был узнан РНК-полимеразой, необходимо образование транскрипционного комплекса ТАТА-фактор/ТАТА-бокс (промотор). ТАТА-фактор остаётся связанным с ТАТА-боксом во время транскрипции, это облегчает использование промотора многими молекулами РНК-полимеразы; 2 - образование транскрипционной вилки; 3 - элонгация; 4.- терминация.
строго определенных участках матрицы - терминаторах (сайты терминации). Фактор терминации облегчает отделение первичного транскрипта (пре-мРНК), комплементарного матрице, и РНК-полимеразы от матрицы. РНК-полимераза может вступить в следующий цикл транскрипции после присоединения субъединицы σ.
Транскрипция в биологии, осуществляющийся в живых клетках биосинтез рибонуклеиновой кислоты (РНК) на матрице — дезоксирибонуклеиновой кислоте (ДНК). Транскрипция (в биологии) — один из фундаментальных биологических процессов, первый этап реализации генетической информации, записанной в ДНК в виде линейной последовательности 4 типов мономерных звеньев — нуклеотидов.
Репликация ДНК; молекулярные механизмы и биологическая роль.
Реплика́ция ДНК — процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты на матрице родительской молекулы ДНК. В ходе последующего деления материнской клетки каждая дочерняя клетка получает по одной копии молекулы ДНК, которая является идентичной ДНК исходной материнской клетки. Этот процесс обеспечивает точную передачу генетической информации из поколения в поколение. Репликацию ДНК осуществляет сложный ферментный комплекс, состоящий из 15—20 различных белков, называемый реплисомой (англ. replisome).
Репликация ДНК — ключевое событие в ходе деления клетки. Принципиально, чтобы к моменту деления ДНК была реплицирована полностью и при этом только один раз. Это обеспечивается определёнными механизмами регуляции репликации ДНК. Репликация проходит в три этапа:
инициация репликации
элонгация
терминация репликации.
Регуляция репликации осуществляется в основном на этапе инициации. Это достаточно легко осуществимо, потому что репликация может начинаться не с любого участка ДНК, а со строго определённого, называемого сайтом инициации репликации. В геноме таких сайтов может быть как всего один, так и много. С понятием сайта инициации репликации тесно связано понятие репликон. Репликон — это участок ДНК, который содержит сайт инициации репликации и реплицируется после начала синтеза ДНК с этого сайта. Геномы бактерий, как правило, представляют собой один репликон, это значит, что репликация всего генома является следствием всего одного акта инициации репликации. Геномы эукариот (а также их отдельные хромосомы) состоят из большого числа самостоятельных репликонов, это значительно сокращает суммарное время репликации отдельной хромосомы. Молекулярные механизмы, которые контролируют количество актов инициации репликации в каждом сайте за один цикл деления клетки, называются контролем копийности. В бактериальных клетках помимо хромосомной ДНК часто содержатся плазмиды, которые представляют собой отдельные репликоны. У плазмид существуют свои механизмы контроля копийности: они могут обеспечивать синтез как всего одной копии плазмиды за клеточный цикл, так и тысяч копий.
Репликация начинается в сайте инициации репликации с расплетания двойной спирали ДНК, при этом формируется репликационная вилка — место непосредственной репликации ДНК. В каждом сайте может формироваться одна или две репликационные вилки в зависимости от того, является ли репликация одно- или двунаправленной. Более распространена двунаправленная репликация. Через некоторое время после начала репликации в электронный микроскоп можно наблюдать репликационный глазок — участок хромосомы, где ДНК уже реплицирована, окруженный более протяженными участками нереплицированной ДНК.
В репликационной вилке ДНК копирует крупный белковый комплекс (реплисома), ключевым ферментом которого является ДНК-полимераза. Репликационная вилка движется со скоростью порядка 100 000 пар нуклеотидов в минуту у прокариот и 500—5000 — у эукариот.
Молекулярный механизм репликации
Ферменты (хеликаза, топоизомераза) и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и активностью ДНК-полимеразы, способной распознать и исправить ошибку. Репликация у эукариот осуществляется несколькими разными ДНК-полимеразами. Далее происходит закручивание синтезированных молекул по принципу суперспирализации и дальнейшей компактизации ДНК. Синтез энергозатратный.
Цепи молекулы ДНК расходятся, образуют репликационную вилку, и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются две новые двуспиральные молекулы ДНК, идентичные родительской молекуле.
Характеристики процесса репликации
матричный — последовательность синтезируемой цепи ДНК однозначно определяется последовательностью материнской цепи в соответствии с принципом комплементарности;
полуконсервативный — одна цепь молекулы ДНК, образовавшейся в результате репликации, является вновь синтезированной, а вторая — материнской;
идёт в направлении от 5’-конца новой молекулы к 3’-концу;
полунепрерывный — одна из цепей ДНК синтезируется непрерывно, а вторая — в виде набора отдельных коротких фрагментов (фрагментов Оказаки);
начинается с определённых участков ДНК, которые называются сайтами инициации репликации (англ. origin).
Механизм действия стероидных гормонов.
Гормоны стероидной природы представлены жирорастворимыми гормонами коркового вещества надпочечников (кортикостероиды), половыми гормонами (эстрогены и андрогены), а также гормональной формой витамина D.
Стероидные гормоны обладают двумя путями действия на клетки: 1) классическим геномным или медленным и 2) быстрым негеномным.
Геномный механизм действия на клетки-мишени начинается трансмембранным переносом молекул стероидных гормонов в клетку (благодаря их растворимости в липидном бислое клеточной мембраны), с последующим связыванием гормона с цитоплазменным белком-рецептором. Эта связь с рецепторным белком необходима для поступления стероидного гормона в ядро, где происходит его взаимодействие с ядерным рецептором. Последующее взаимодействие комплекса гормон—ядерный рецептор с хромати-новым акцептором, специфическим кислым белком и ДНК влечет за собой: активацию транскрипции специфических мРНК, синтез транспортных и рибосомных РНК, процессинг первичных РНК-транскриптов и транспорт мРНК в цитоплазму, трансляцию мРНК при достаточном уровне транспортных РНК с синтезом белков и ферментов в рибосомах. Все эти явления требуют длительного (часы, сутки) присутствия гормон-рецепторного комплекса в ядре.
Быстрый негеномный
Эффекты стероидных гормонов проявляются не только спустя несколько часов, что требуется для ядерного влияния, часть из них проявляется очень быстро, в течение нескольких минут. Это такие эффекты, как повышение проницаемости мембран, усиление транспорта глюкозы и аминокислот, освобождение лизосомальных ферментов, сдвиги энергетики митохондрий. К числу быстрых негеномных эффектов стероидных гормонов относятся, например, увеличение в течение 5 мин после введения человеку альдосте-рона общего периферического сосудистого сопротивления и артериального давления, изменение транспорта натрия через мембрану эритроцитов (вообще лишенных ядра) под влиянием альдостерона в опытах in vitro, быстрый вход Са2+ в клетки эндометрия под влиянием эстрогенов и др. Механизм негеномного действия стероидных гормонов заключается в связывании на плазматической мембране клетки со специфическими рецепторами и активации каскадных реакций систем вторичных посредников, например фосфолипазы С, инозитол-3-фосфата, ионизированного Са2+, протеинки-назы С. Под влиянием стероидных гормонов в клетке может увеличиваться содержание цАМФ и цГМФ.
Негеномный эффект стероидных гормонов может быть реализован и после их связывания с цитоплазматическими рецепторами. Часть негеномных эффектов стероидных гормонов осуществляется благодаря их взаимодействию с рецепторами, связанными с воротным механизмом ионных каналов мембран нервных клеток, являясь тем самым модуляторами, например, глицин-, серотонин- или гамма-аминобу-тиратергических нейронов. Наконец, растворяясь в липидном бислое мембраны, стероидные гормоны могут менять физические свойства мембраны, такие как ее текучесть или проницаемость для гидрофильных молекул, что также является негеномным эффектом.
Таким образом, механизмы действия гормонов разной химической структуры имеют не только различия, но и общие черты. Как и стероиды, пептидные гормоны обладают способностью избирательно влиять на транскрипцию генов в ядре клетки. Этот эффект пептидных гормонов может быть реализован не только с поверхности клетки при образовании вторичных посредников, но и путем поступления пептидных гормонов внутрь клетки за счет интернализации гормон-рецепторного комплекса.
Мембранно-опосредованный механизм действия пептидных и белковых гормонов.
Мембранный механизм.
Гормон связывается с клеточной мембраной и в месте связывания изменяет её проницаемость для глюкозы, аминокислот и некоторых ионов. В этом случае гормон выступает как эффектор транспортных средств мембраны. Такое действие оказывает инсулин, изменяя транспорт глюкозы. Но этот тип транспорта гормонов редко встречается в изолированном виде. Инсулин, например, обладает как мембранным, так и мембранно-внутриклеточным механизмом действия.
Мембранно-внутриклеточный механизм (или мембранно-опосредованый). По мембранно-внутриклеточному типу действуют гормоны, которые не проникают в клетку и поэтому влияют на обмен веществ через внутриклеточного химического посредника. К ним относят белково-пептидные гормоны (гормоны гипоталамуса, гипофиза, поджелудочной и паращитовидной желез, тиреокальцитонин щитовидной железы); производные аминокислот (гормоны мозгового слоя надпочечников - адреналин и норадреналин, щитовидной железы - тироксин, трийодтиронин).
Функции внутриклеточных химических посредников гормонов выполняют циклические нуклеотиды - циклический 3׳,5׳ - аденозинмонофосфат (цАМФ) и циклический 3׳,5׳ - гуанозинмонофосфат (цГМФ), ионы кальция.
Гормоны влияют на образование циклических нуклеотидов: цАМФ - через аденилатциклазу, цГМФ - через гуанилатциклазу.
Аденилатциклаза встроена в мембрану клетки и состоит из 3-х взаимосвязанных частей: рецепторной (R), представленной набором мембранных рецепторов, находящихся снаружи мембраны, сопрягающей (N), представленной особым N -белком, расположенным в липидном слое мембраны, и каталитической (C), являющейся ферментным белком, то есть собственно аденилатциклазой, которая превращает АТФ (аденозинтрифосфат) в цАМФ.
Аденилатциклаза работает по слудующей схеме. Как только гормон связывается с рецептором (R) и образуется комплекс гормон- рецептор, происходит образовагние комплекса N - белок - ГТФ (гуанозинтрифосфат), который активирует каталитическую (С) часть аденилатцеклазы. Активация аденилатциклазы приводит к образованию цАМФ внутри клетки на внутренней поверхности мембраны из АТФ.
Даже одна молекула гормона, связавшегося с рецептором, заставляет работать аденилатцеклазу. При этом на одну молекулу связавшегося гормона образуется 10-100 молекул цАМФ внутри клетки. В активном состоянии аденилатциклаза находится до тех пор, пока существует комплекс гормон - рецептор. Аналогичным образом работает и гуанилатциклаза.
В цитоплазме клетки находятся неактивные протеинкиназы. Циклические нуклеотиды- цАМФ ицГМФ- активируют пртеинкиназы. Существуют цАМФ- зависимые и цГМФ - зависимые протеинкиназы, которые активируются своим циклическим нуклеотидом. В зависимости от мембранного рецептора, связывающего определенный гормон, включается или аденилатцеклаза, или гуанилатцеклазаи соответственно происходит образование или цАМФ, или цГМФ.
Назовите а-кетокислоты, образующиеся из аминокислот (аспартата, аланина) в реакциях трансаминирования с а-кетоглутаратом. Опишите механизм трансаминирования.
(????????)
Назовите пути образования и распада аминокислот. Декарбоксилирование аминокислот. Физиологическая роль продуктов этого процесса.
Декарбоксилирование аминокислот- процесс отщепления карбоксильной группы аминокислот в виде CO2 получил название декарбоксилирования. Несмотря на ограниченный круг аминокислот и их производных, подвергающихся декарбоксилированию в животных тканях, образующиеся продукты реакции – биогенные амины (т.н. "Трупные яды") - оказывают сильное фармакологическое действие на множество физиологических функций человека и животных. В животных тканях установлено декарбоксилирование следующих аминокислот и их производных: тирозина, триптофана, 5-окситриптофана, валина, серина, гистидина, глутаминовой и γ-оксиглутаминовой кислот, 3,4-диоксифенилаланина, цистеина, аргинина, орнитина, S-аденозилметионина и α-аминомалоновой кислоты. Помимо этого, у микроорганизмов и растений открыто декарбоксилирование ряда других аминокислот.
В живых организмах открыты 4 типа декарбоксилирования аминокислот:
1. α-Декарбоксилирование, характерное для тканей животных, при котором от аминокислот отщепляется карбоксильная группа, стоящая по соседству с α-углеродным атомом. Продуктами реакции являются CO2 и биогенные амины:
2. ω-Декарбоксилирование, свойственное микроорганизмам. Например, из аспарагиновой кислоты этим путём образуется α-аланин:
3. Декарбоксилирование, связанное с реакцией трансаминирования:
В этой реакции образуются альдегид и новая аминокислота, соответствующая исходной кетокислоте.
4. Декарбоксилирование, связанное с реакцией конденсации двух молекул:
Эта реакция в тканях животных осуществляется при синтезе δ-амино-левулиновой кислоты из глицина и сукцинил-КоА и при синтезе сфинголипидов, а также у растений при синтезе биотина.
Реакции декарбоксилирования в отличие от других процессов промежуточного обмена аминокислот являются необратимыми. Они катализируются специфическими ферментами – декарбоксилазами аминокислот, отличающимися от декарбоксилаз α-кетокислот как белковым компонентом, так и природой кофермента. Декарбоксилазы аминокислот состоят из белковой части, обеспечивающей специфичность действия, и простетической группы, представленной пиридоксальфосфатом (ПФ), как и у трансаминаз.
Таким образом, в двух совершенно различных процессах обмена аминокислот участвует один и тот же кофермент. Исключение составляют две декарбоксилазы: гистидиндекарбоксилаза Micrococcus и Lactobacilus и аденозилметионин-декарбоксилаза Е. coli, содержащие вместо ПФ остаток пировиноградной кислоты.
Механизм реакции декарбоксилирования аминокислот в соответствии с общей теорией пиридоксалевого катализа сводится к образованию ПФ-субстратного комплекса, представленного, как и в реакциях трансаминирования, шиффовым основанием ПФ и аминокислоты:
Образование подобного комплекса в сочетании с некоторым оттягиванием электронов белковой частью молекулы фермента сопровождается лабилизацией одной из трех связей при α-углеродном атоме, благодаря чему аминокислота способна вступать в реакции трансаминирования (а), декарбоксилирования (b) и альдольного расщепления (с).
Далее представлены отдельные примеры декарбоксилирования аминокислот, в частности тех, продукты реакции которых оказывают сильное фармакологическое действие. Одним из хорошо изученных ферментов является декарбоксилаза ароматических аминокислот. Она не обладает строгой субстратной специфичностью и катализирует декарбок-силирование L-изомеров триптофана, 5-окситриптофана и 3,4-диоксифе-нилаланина (ДОФА); продуктами реакций, помимо CO2, являются соответственно триптамин, серотонин и диоксифенилэтиламин (дофамин).
Декарбоксилаза ароматических аминокислот получена в чистом виде (мол. масса 112000), кофермент – ПФ. В больших количествах она содержится в надпочечниках и ЦНС, играет важную роль в регуляции содержания биогенных аминов. Образующийся из 5-окситриптофана серо-тонин оказался высокоактивным биогенным амином сосудосуживающего действия. Серотонин регулирует артериальное давление, температуру тела, дыхание, почечную фильтрацию и является медиатором нервных процессов в ЦНС. Некоторые авторы считают серотонин причастным к развитию аллергии, демпинг-синдрома, токсикоза беременных, карциноидного синдрома и геморрагических диатезов.
Продукт декарбоксилазной реакции дофамин является предшественником катехоламинов (норадреналина и адреналина). Источником ДОФА в организме является тирозин, который под действием специфической гидроксилазы превращается в 3,4-диоксифенилаланин. Тиро-зин-3-монооксигеназа открыта в надпочечниках, ткани мозга и периферической нервной системы. Простетической группой тирозин-моноокси-геназы, как и дофамин-монооксигеназы (последняя катализирует превращение дофамина в норадреналин) является тетрагидробиоптерин, имеющий следующее строение:
Физиологическая роль тирозин-3-монооксигеназы чрезвычайно велика, поскольку катализируемая этим ферментом реакция определяет скорость биосинтеза катехоламинов, регулирующих деятельность сердечно-сосудистой системы. В медицинской практике широко используются ингибиторы декарбоксилазы ароматических аминокислот, в частности α-метилдофа (альдомет), вызывающий снижение артериального давления.
В клинической практике широко используется, кроме того, продукт α-декарбоксилирования глутаминовой кислоты – γ-аминомасляная кислота (ГАМК). Фермент, катализирующий эту реакцию (глутаматдекарбокси-лаза), является высокоспецифичным.
Назовите а-кетокислоты, образующиеся из аминокислот (аспартата, аланина) в реакциях трансаминирования с а-кетоглутаратом.(?????)
27. Фосфоролиз
Ферментативная реакция расщепления химических связей в некоторых биологически важных соединениях с участием фосфорной кислоты; сопровождается включением фосфорильной группы (–H2PO3) в образующиеся продукты. Ферменты, катализирующие Ф., называются фосфорилазами (См. Фосфорилазы). Ф. широко распространён в процессах обмена веществ у животных, растений и микроорганизмов. Фосфоролитическому расщеплению под действием ферментов могут подвергаться гликозидные (в гликогене), тиоэфирные (в ферментсубстратном комплексе, образующемся при окислении 3-фосфоглицеринового альдегида), углерод-углеродные (в ксилулозо-5-фосфате, в пировиноградной кислоте), фосфодиэфирные (в нуклеиновых кислотах) и углерод-азотные (в цитруллине) связи. Ф. играет важную роль в энергетике живых систем, т.к. фосфорильная группа, включенная в продукты реакции, под действием различных ферментов в конечном счёте переносится на аденозиндифосфорную кислоту с образованием аденозинтрифосфорной кислоты (АТФ) – основного энергетического ресурса клеток.
28. Переваривание жиров. Этапы переваривания жиров в кишечнике
Несомненно, что в повседневной пище из жиров доминируют нейтральные жиры, известные как триглицериды, каждая молекула которых включает глицериновое ядро и боковые цепи, состоящие из трех жирных кислот. Нейтральные жиры — основной компонент животной пищи, а в растительной пище их содержится крайне мало.
В обычной пище имеется небольшое количество фосфолипидов, холестерола и эфиров холестерола. Фосфолипиды и эфиры холестерола содержат жирные кислоты и, следовательно, могут рассматриваться как жиры. Впрочем, холестерол является представителем стеринов и не содержит жирных кислот, но проявляет некоторые физические и химические свойства жиров; к тому же он производится из жиров и легко превращается в них. Следовательно, с диетологической точки зрения, холестерол рассматривается как жир.
Переваривание жиров в кишечнике. Небольшое количество триглицеридов переваривается в желудке под действием лингвальной липазы, которая секретируется железами языка в ротовой полости и проглатывается вместе со слюной. Количество перевариваемых таким образом жиров составляет менее 10%, а потому не существенно. Основное переваривание жиров происходит в тонком кишечнике, о чем сказано далее.
Эмульгирование жиров желчными кислотами и лецитином. Первый этап переваривания жиров заключается в физическом разрушении капель жира на мелкие частицы, поскольку водорастворимые ферменты могут действовать только на поверхности капли. Этот процесс называют эмульгированием жиров, он начинается в желудке с перемешивания жиров с другими продуктами переваривания желудочного содержимого.
Далее основной этап эмульгирования происходит в двенадцатиперстной кишке под влиянием желчи, секрета печени, который не содержит пищеварительных ферментов. Однако желчь содержит большое количество желчных солей, а также фосфолипид — лецитин. Эти компоненты, в особенности лецитин, чрезвычайно важны для эмульгирования жиров. Полярные частицы (места, в которых происходит ионизация воды) желчных солей и молекул лецитина хорошо растворимы в воде, тогда как большая оставшаяся часть этих молекул хорошо растворима в жирах.
Таким образом, жирорастворимые порции секрета печени растворяются в поверхностном слое жировых капель вместе с выступающей полярной частью. В свою очередь, выступающая полярная часть растворима в окружающей водной фазе, что значительно снижает поверхностное натяжение жиров и делает их также растворимыми.
Когда поверхностное натяжение капли нерастворимой жидкости низкое, нерастворимая в воде жидкость во время перемещения значительно легче разрушается на множество мелких частиц, чем при более высоком поверхностном натяжении. Следовательно, основная функция желчных солей и лецитина — делать капли жира способными к легкому размельчению при перемешивании с водой в тонком кишечнике. Это действие аналогично действию синтетических моющих средств, широко используемых в домашнем хозяйстве для устранения жира.
Каждый раз в результате перемешивания в тонком кишечнике диаметр жировых капель существенно уменьшается, поэтому общая жировая поверхность многократно увеличивается. Из-за того, что средний диаметр частичек жира в кишечнике после эмульгирования оказывается менее 1 мкм, общая площадь жировой поверхности, образованная в результате процесса эмульгирования, увеличивается в 1000 раз.
Фермент липаза является водорастворимым и может воздействовать только на поверхности жировых капель. Отсюда понятно, насколько значительна детергентная роль лецитина и желчных солей в переваривании жиров.
29. Расщепление нуклеиновых кислот под влиянием специфических ферментов - эндо - и экзонуклеаз - сопровождается разрывом фосфо-диэфирной связи и образованием продуктов различной величины, которые могут быть разделены методами электрофореза и хроматографии. Это широко используется при анализе последовательности нук-леотидов в молекулах РНК и ДНК. Особое значение при развитии генной инженерии получило расщепление ДНК специфическими эндо-нуклеазами ( рестриктазами), позволяющее получать отрезки ДНК определенной длины и нуклеотидного состава. [1]
Характер продуктов расщепления нуклеиновых кислот при их облучении указывает на разрыв водородных связей, что можно представить как окисление. То же можно сказать и об облучении биологически важных соединений, содержащих сульфгидрильные группы, в которых SH-группы переходят в S - S-гругаш. [2]
Собственно хроматографическому анализу предшествует расщепление нуклеиновых кислот, которое можно проводить химическим или ферментативным путем. Продуктами расщепления являются: а) пуриновые и иири-мидиновые основания, б) мононуклеотиды, в) нуклеозиды. [3]
НУКЛЕАЗЫ, ферменты, вызывающие в организме расщепление нуклеиновых кислот на мононуклеотиды, отщепление от монопуклеотидов фосфорной к-ты с образованием нуклеозидов и расщепление нуклеозидов на пеитозу, пуриновые и пиримиднновые основания.
Применяемые для определения нуклеотидной последовательности РНК методы сводятся к контролируемому расщеплению нуклеиновых кислот различными ферментами и последующему разделению продуктов гидролиза. [5]
30. Строение рибосом
Рибосома представляет собой элементарную клеточную машину синтеза любых белков клетки. Все они построены в клетке одинаково, имеют одинаковую молекулярную композицию, выполняют одинаковую функцию - синтез белка - поэтому их можно так же считать клеточными органоидами. В отличие от других органоидов цитоплазмы (пластид, митохондрий, клеточного центра, мембранной вакуолярной системы и др.) они представлены в клетке огромным числом: за клеточный цикл их образуется 1 х 107 штук. Поэтому основная масса клеточной РНК представляет собой именно рибосомную РНК. РНК рибосом относительно стабильна, рибосомы могут существовать в клетках культуры ткани в течение нескольких клеточных циклов. В печеночных клетках время полужизни рибосом составляет 50-120 часов.
Рибосомы - это сложные рибонуклеопротеидные частицы, в состав которых входит множество молекул индивидуальных (неповторенных) белков и несколько молекул РНК, Рибосомы прокариот и эукариот по своим размерам и молекулярным характеристикам отличаются, хотя и обладают общими принципами организации и функционирования. К настоящему времени методом рентгеноструктурного анализа высокого разрешения полностью расшифрована структура рибосом.
Полная, работающая рибосома, состоит из двух неравных субъединиц, которые легко обратимо диссоциируют на большую субъединицу и малую. Размер полной прокариотической рибосомы составляет 20 х 17 х 17 нм, эукариотической - 25 х 20 х 20. Полная прокариотическая рибосома имеет коэффициент седиментации 70S и диссоциирует на две субъединицы: 50S и 30S. Полная эукариотическая рибосома, 80S рибосома, диссоциирует на 60S и 40S субъединицы. Форма и детальные очертания рибосом из разнообразных организмов и клеток, включая как прокариотические, так и эукариотические, поразительно похожи, хотя и отличаются рядом деталей. Малая рибосомная субъединица имеет палочковидную форму с несколькими небольшими выступами, ее длина составляет около 23 нм, а ширина - 12 нм. Большая субъединица похожа на полусферу с тремя торчащими выступами. При ассоциации в полную 70S рибосому малая субчастица ложится одним концом на один из выступов 50S частицы, а другим в ее желобок. В состав малых субъединиц входит по одной молекуле РНК, а в состав большой - несколько: у прокариот - две, а у эукариот - 3 молекулы
ЧАСТЬ С
1. Обмен клетки направлен на поддержание и умножение веществ, из которых состоит клетка. Для этого необходимы энергия и строительные блоки. И то и другое получается в результате переработки поступающих извне веществ внутри клетки. Энергетический обмен, или катаболизм, ведет к получению энергии, а конструктивный обмен, или анаболизм — к образованию строительных блоков. Они составляют серии последовательных реакций, образующих пути метаболизма и объединенных в метаболическую сеть. Катаболизм и анаболизм в той или иной степени сопряжены, и общую их часть иногда называют амфиболизмом или центральными метаболическими реакциями. Следует заметить, что специальные пути катаболизма свойственны литотрофным микроорганизмам, окисляющим неорганические вещества. В этом случае промежуточные метаболиты не обязательно принадлежат амфиболическим реакциям.
Метаболизм бактерий определяет «типы обмена» или «трофии» — основу группирования их по «физиологическим группам» (см. часть 2). Каждый тип питания характеризуется определенными путями обмена и соответствующими ключевыми ферментами. Пути метаболизма составляют существенную часть биохимии. Здесь нет возможности излагать их, и придется ограничиться лишь кратким перечислением и ролью в системе. На русский язык переведены краткие учебники Г. Шлегеля «Общая микробиология» и Г. Готтшалка «Метаболизм бактерий», сконцентрированные на обмене бактерий, где этот аспект изложен достаточно подробно и ясно для неспециалиста.
Среди процессов катаболизма, осуществляемых микроорганизмами, первостепенное значение имеет разложение углеводов (глюкозы). Наиболее распространенным является путь Эмбдена-Мейергофа, или гликолиз (рис. 9), в котором при окислении одной молекулы глюкозы образуются в конечном итоге две молекулы пирувата, выигрываются две молекулы АТФ и две молекулы восстановителя (НАДН). Такой путь характерен для многих анаэробных органотрофных сахаролитических организмов, осуществляющих брожение углеводов. Проблему для анаэробов представляет удаление избытка восстановителя и регенерация НАД как окислителя. Обычно акцепторами служат органические соединения, в результате их восстановления образуются летучие жирные кислоты (ЛЖК), спирты и Н2. По набору образуемых продуктов различают спиртовое, маслянокислое, молочнокислое, пропионовое, смешанное брожения (рис. 10) и соответствующие функциональные группы анаэробов.
Многие бактерии окисляют глюкозу и по другому пути, Энт-нера-Дудорова, с промежуточным образованием 2-кето-З-дезокси-6-фосфоглюконовой кислоты. Итогом являются 2 молекулы пирувата, 2 молекулы НАДН и АТФ.
Наконец, разложение глюкозы через пентозофосфатныи окислительный цикл включает ряд реакций окисления и декарбокси-лирования глюкозо-6-фосфата с образованием рибулозо-5-фосфата и СО2. Далее следует серия превращений фосфатов Сахаров и регенерация глюкозо-6-фосфата. Одним из интермедиатов пути является 3-фосфоглицерат, который легко может быть превращен в пируват. Пентозофосфатныи цикл имеет ряд сходных ступеней с циклом Кальвина — циклическим путем фиксации СО2 у большинства автотрофов, а также с путем ассимиляции формальдегида у метилотрофов. Это обеспечивается полной обратимостью реакций между фосфосахарами, находящихся в равновесии между собой, благодаря действию ферментов трансальдолазы и транскетолазы.
2. Первой реакцией гликолиза является фосфорилирование молекулы глюкозы, происходящее при участии тканеспецифичного фермента гексокиназы с затратой энергии 1 молекулы АТФ; образуется активная форма глюкозы — глюкозо-6-фосфат (Г-6-Ф):
Для протекания реакции необходимо наличие в среде ионов Mg2+, с которым комплексно связывается молекула АТФ. Эта реакция необратима и является первой ключевой реакцией гликолиза.
Фосфорилирование глюкозы преследует две цели: во-первых, из-за того что плазматическая мембрана, проницаемая для нейтральной молекулы глюкозы, не пропускает отрицательно заряженные молекулы Г-6-Ф, фосфорилированная глюкоза оказывается запертой внутри клетки. Во-вторых, при фосфорилировании глюкоза переводится в активную форму, способную участвовать в биохимических реакциях и включаться в метаболические циклы. Фосфорилирование глюкозы — это единственная реакция в организме, в которой глюкоза участвует как таковая.
Печёночный изофермент гексокиназы — глюкокиназа — имеет важное значение в регуляции уровня глюкозы в крови.
В следующей реакции (2) ферментом фосфоглюкоизомеразой Г-6-Ф превращается во фруктозо-6-фосфат (Ф-6-Ф):
Энергия для этой реакции не требуется, и реакция является полностью обратимой. На данном этапе в процесс гликолиза может также включаться путём фосфорилирования и фруктоза.
Далее почти сразу друг за другом следуют две реакции: необратимое фосфорилирование фруктозо-6-фосфата (3) и обратимое альдольное расщепление образовавшегося фруктозо-1,6-бифосфата (Ф-1,6-бФ) на две триозы (4).
Фосфорилирование Ф-6-Ф осуществляется фосфофруктокиназой с затратой энергии ещё одной молекулы АТФ; это вторая ключевая реакция гликолиза, её регуляция определяет интенсивность гликолиза в целом.
Альдольное расщепление Ф-1,6-бФ происходит под действием альдолазы фруктозо-1,6-бифосфата:
В результате четвёртой реакции образуются дигидроксиацетонфосфат и глицеральдегид-3-фосфат, причём первый почти сразу под действием фосфотриозоизомеразы переходит во второй (5), который и участвует в дальнейших превращениях:
Каждая молекула глицеральдегидфосфата окисляется НАД+ в присутствии дегидрогеназы глицеральдегидфосфата до 1,3-дифосфоглицерата (6):
Далее с 1,3-дифосфоглицерата, содержащего макроэргическую связь в 1 положении, ферментом фосфоглицераткиназой на молекулу АДФ переносится остаток фосфорной кислоты (реакция 7) — образуется молекула АТФ:
Это первая реакция субстратного фосфорилирования. С этого момента процесс расщепления глюкозы перестаёт быть убыточным в энергетическом плане, так как энергетические затраты первого этапа оказываются компенсированными: синтезируются 2 молекулы АТФ (по одной на каждый 1,3-дифосфоглицерат) вместо двух потраченных в реакциях 1 и 3. Для протекания данной реакции требуется присутствие в цитозоле АДФ, то есть при избытке в клетке АТФ (и недостатке АДФ) её скорость снижается. Поскольку АТФ, не подвергающийся метаболизму, в клетке не депонируется а просто разрушается, то эта реакция является важным регулятором гликолиза.
Затем последовательно: фосфоглицеролмутаза образует 2-фосфоглицерат (8):
Енолаза образует фосфоенолпируват (9):
И наконец происходит вторая реакция субстратного фосфорилирования АДФ с образованием енольной формы пирувата и АТФ (10):
Реакция протекает под действием пируваткиназы. Это последняя ключевая реакция гликолиза. Изомеризация енольной формы пирувата в пируват происходит неферментативно.
С момента образования Ф-1,6-бФ с выделением энергии протекают только реакции 7 и 10, в которых и происходит к субстратное фосфорилирование АДФ.
3. Гликогенолиз и глюконеогенез
Существует всего три способа поступления глюкозы в кровь. Первый (нефизиологический путь) заключается во введении глюкозы либо внутривенным, либо каким-то иным парентеральным способом. Такой путь может приобретать особое значение в диагностике и лечении больных, а также при некоторых экспериментальных подходах. Второй (и привычный) путь – это всасывание глюкозы в пищеварительном тракте. В обычных условиях человек и животное обладают двумя основными источниками калорий: жировой тканью и содержимым пищеварительного тракта. Легко убедиться, что нарушение всасывания глюкозы или других углеводов нарушает систему поддержания постоянной концентрации глюкозы в крови, чем характеризуются некоторые заболевания человека. Третий путь поступления глюкозы в кровь открыт Клодом Бернаром и заключается во «внутренней секреции» глюкозы печенью. Было установлено, что даже в отсутствие явных гормональных эффектов печень обладает определенной степенью автономии в отношении количества сахара, выделяемого в печеночные вены или задерживаемого в ткани. Однако этот автономный контроль моделируется множеством гормональных сигналов. При необходимости в ответ на соответствующие гормональные сигналы печень может поддерживать постоянную концентрацию глюкозы в крови даже в отсутствие всасывания глюкозы в пищеварительном тракте.
Способность печени секретировать глюкозу в кровь обусловлена в основном двумя процессами. Первый из них – гликогенолиз – протекает в течение минут, тогда как второй – глюконеогенез – в течение часов и суток.
Гликогенолиз
Гликогенолиз – это просто распад накопившегося в печени гликогена до глюкозо-6-фосфата. Последний под действием специфической фосфатазы (содержащейся в печени, но не в мышцах) расщепляется с высвобождением в кровь свободной глюкозы.
Это высвобождение глюкозы из печени, по-видимому, служит адекватным средством адаптации к острым потребностям, но постоянная концентрация глюкозы в крови не могла бы сохраняться длительное время, даже если бы весь гликоген из печени поступил в кровь в виде глюкозы. Масса печени человека – около 1500 г; если на долю гликогена приходится даже 4% массы органа, то полный гликогенолиз мог бы обеспечить поступление в кровь не более 60 г глюкозы (или всего 240 калорий) в условиях длительного лишения глюкозы (при голодании) или в отсутствие реабсорбции глюкозы в почках (при флоридзиновом диабете), для эффективного поддержания постоянной концентрации глюкозы в крови необходимо участие какого-то иного механизма. Этот другой, длительный процесс, происходящий в печени, называется глюконеогенез.
4. Гликолиз. Гликолитический путь окисления. Путь Эмбдена-Мейерхофа-Парнаса.
Гликолитический путь (гликолиз, путь Эмбдена-Мейерхофа-Парнаса, фруктозо-1,6-дифосфатный путь) доминирует у большинства аэробных и анаэробных микроорганизмов — кишечной палочки, бацилл, пенициллов, дрожжей, стрептомицетов и многих других. Характерная реакция гликолиза — расщепление фруктозо-1,6-дифосфата альдолазой, в результате чего образуется смесь триозофосфатов, состоящая из дигидрооксиацетонфосфата и глицероальдегидтрифосфата, которые затем превращаются в пируват (рис. 4-8). Фермент обратной связи, лимитирующий скорость процесса, — фруктозо-6-фосфат дегидрогеназа (фосфофруктокиназа). В этом процессе образуется 2 моля АТФ и 2 моля восстановленного НАД (НАДН+ на 1 моль глюкозы.
В результате изомеризации фруктозо-6-фосфата в глюкозо-6-фосфат и конденсации двух молекул триозо-3-фосфата в гексозофосфат все перечисленные реакции замыкаются в цикл, при одном обороте которого из 3 молекул глюкозо-6-фосфата образуются 2 молекулы фруктозо-6-фосфата, одна молекула триозо-3-фосфата, 3 молекулы С02 и трижды по 2 восстановленных НАДФ (НАДФН+). Последовательность подобных превращений глюкозы у бактерий идентична таковой у высших организмов. Этот путь катаболизма глюкозы следует рассматривать как второстепенный, но имеющий специальное назначение — источник пентоз для синтеза нуклеиновых кислот, а также источник восстановительных эквивалентов НАДФН+.
5. ПЕНТОЗОФОСФАТНЫЙ ПУТЬ ОКИСЛЕНИЯ УГЛЕВОДОВ
Открытие пути прямого окисления углеводов, или, как его называют, пентозофосфатного цикла, принадлежит О. Варбургу, Ф. Липману, Ф. Ди-кенсу и В.А. Энгельгарду. Расхождение путей окисления углеводов – классического (цикл трикарбоновых кислот, или цикл Кребса) и пентозофос-фатного – начинается со стадии образования гексозомонофосфата. Если глюкозо-6-фосфат изомеризуется во фруктозо-6-фосфат, который фосфо-рилируется второй раз и превращается во фруктозо-1,6-бисфосфат, то в этом случае дальнейший распад углеводов происходит по обычному гликолитическому пути с образованием пировиноградной кислоты, которая, окисляясь до ацетил-КоА, затем «сгорает» в цикле Кребса.
Если второго фосфорилирования гексозо-6-монофосфата не происходит, то фосфорилированная глюкоза может подвергаться прямому окислению до фосфопентоз. В норме доля пентозофосфатного пути в количественном превращении глюкозы обычно невелика, варьирует у разных организмов и зависит от типа ткани и ее функционального состояния.
У млекопитающих активность пентозофосфатного цикла относительно высока в печени, надпочечниках, эмбриональной ткани и молочной железе в период лактации. Значение этого пути в обмене веществ велико. Он поставляет восстановленный НАДФН, необходимый для биосинтеза жирных кислот, холестерина и т.д. За счет пентозофосфатного цикла примерно на 50% покрывается потребность организма в НАДФН.
Другая функция пентозофосфатного цикла заключается в том, что он поставляет пентозофосфаты для синтеза нуклеиновых кислот и многих коферментов. При ряде патологических состояний удельный вес пенто-зофосфатного пути окисления глюкозы возрастает. Механизм реакций пентозофосфатного цикла достаточно расшифрован.
Рис. 10.12. Пентозофосфатный путь окисления углеводов.
Пентозофосфатный цикл начинается с окисления глюкозо-6-фосфата и последующего окислительного декарбоксилирования продукта (в результате от гексозофосфата отщепляется первый атом углерода). Это первая, так называемая окислительная, стадия пентозофосфатного цикла. Вторая стадия включает неокислительные превращения пентозофосфатов с образованием исходного глюкозо-6-фосфата (рис. 10.12). Реакции пен-тозофосфатного цикла протекают в цитозоле клетки.
Первая реакция – дегидрирование глюкозо-6-фосфата при участии фермента глюкозо-6-фосфатдегидрогеназы и кофермента НАДФ+. Образовавшийся в ходе реакции 6-фосфоглюконо-δ-лактон – соединение нестабильное и с большой скоростью гидролизуется либо спонтанно, либо с помощью фермента 6-фосфоглюконолактоназы с образованием 6-фос-фоглюконовой кислоты (6-фосфоглюконат):
Во второй – окислительной – реакции, катализируемой 6-фосфоглюко-натдегидрогеназой (декарбоксилирующей), 6-фосфоглюконат дегидрируется и декарбоксилируется. В результате образуется фосфорилированная кетопентоза – D-рибулозо-5-фосфат и еще 1 молекула НАДФН:
Под действием соответствующей эпимеразы из рибулозо-5-фосфата может образоваться другая фосфопентоза – ксилулозо-5-фосфат. Кроме того, рибулозо-5-фосфат под влиянием особой изомеразы легко превращается в рибозо-5-фосфат. Между этими формами пентозофосфатов устанавливается состояние подвижного равновесия:
При определенных условиях пентозофосфатный путь на этом этапе может быть завершен. Однако при других условиях наступает так называемый неокислительный этап (стадия) пентозофосфатного цикла. Реакции этого этапа не связаны с использованием кислорода и протекают в анаэробных условиях. При этом образуются вещества, характерные для первой стадии гликолиза (фруктозо-6-фосфат, фруктозо-1,6-бисфосфат, фосфотрио-зы), а другие – специфические для пентозофосфатного пути (седогептуло-зо-7-фосфат, пентозо-5-фосфаты, эритрозо-4-фосфат).
Основными реакциями неокислительной стадии пентозофосфатного цикла являются транскетолазная и трансальдолазная. Эти реакции катализируют превращение изомерных пентозо-5-фосфатов:
Коферментом в транскетолазной реакции служит ТПФ, играющий роль промежуточного переносчика гликольальдегидной группы от ксилулозо-5-фосфата к рибозо-5-фосфату. В результате образуется семиуглеродный моносахарид седогептулозо-7-фосфат и глицеральдегид-3-фосфат.
Транскетолазная реакция в пентозном цикле встречается дважды, второй раз – при образовании фруктозо-6-фосфата и триозофосфата в результате взаимодействия второй молекулы ксилулозо-5-фосфата с эритро-зо-4-фосфатом:
Фермент трансальдолаза катализирует перенос остатка диоксиацетона (но не свободного диоксиацетона) от седогептулозо-7-фосфата на гли-церальдегид-3-фосфат:
Шесть молекул глюкозо-6-фосфата, вступая в пентозофосфатный цикл, образуют 6 молекул рибулозо-5-фосфата и 6 молекул СО2, после чего из 6 молекул рибулозо-5-фосфата снова регенерируется 5 молекул глюко-зо-6-фосфата (см. рис. 10.12). Однако это не означает, что молекула глюкозо-6-фосфата, вступающая в цикл, полностью окисляется. Все 6 молекул СО2 образуются из С-1-атомов 6 молекул глюкозо-6-фосфата.
Валовое уравнение окислительной и неокислительной стадий пенто-зофосфатного цикла можно представить в следующем виде:
или
Образовавшийся НАДФН используется в цитозоле на восстановительные синтезы и, как правило, не участвует в окислительном фосфори-лировании, протекающем в митохондриях.
В последние годы появились работы, которые дают основание предполагать, что в некоторых тканях схема пентозофосфатного превращения углеводов сложнее, чем это представлено на рис. 10.12. Согласно этой более полной схеме пентозофосфатного пути, первые этапы превращения совпадают с прежней схемой, однако после первой транскетолазной реакции начинаются некоторые отклонения (рис. 10.13).
Считают, что пентозофосфатный путь и гликолиз, протекающие в ци-тозоле, взаимосвязаны и способны переключаться друг на друга в зависимости от соотношения концентраций промежуточных продуктов, образовавшихся в клетке
7. Глюконеогенез — процесс образования в печени и отчасти в корковом веществе почек (около 10 %) молекул глюкозы из молекул других органических соединений — источников энергии, например свободных аминокислот, молочной кислоты, глицерина. Свободные жирные кислоты у млекопитающих для глюконеогенеза не используются. Стадии глюконеогенеза
Стадии глюконеогенеза повторяют стадии гликолиза в обратном направлении и катализируются теми же ферментами за исключением 4 реакций:
Гликолиз и Глюконеогенез
Превращение пирувата в оксалоацетат (фермент пируваткарбоксилаза)
Превращение оксалоацетата в фосфоенолпируват (фермент фосфоенолпируваткарбоксикиназа)
Превращение фруктозо-1,6-дифосфата в фруктозо-6-фосфат (фермент фруктозо-1,6-дифосфатаза)
Превращение глюкозо-6-фосфата в глюкозу (фермент глюкозо-6-фосфатаза)
Суммарное уравнение глюконеогенеза: 2 CH3COCOOH + 4ATP + 2GTP + 2NADH.H+ + 6 H2O = C6H12O6 + 2NAD + 4ADP + 2GDP + 6Pn[1]. Роль в организме
При голодании в организме человека активно используются запасы питательных веществ (гликоген, жирные кислоты). Они расщепляются до аминокислот, кетокислот и других неуглеводных соединений. Большая часть этих соединений не выводится из организма, а подвергаются реутилизации. Вещества транспортируются кровью в печень из других тканей, и используются в глюконеогенезе для синтеза глюкозы — основного источника энергии в организме. Таким образом при истощении запасов организма, глюконеогенез является основным поставщиком энергетических субстратов.
8) Окисление пирувата до ацетил-КоА происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название «пируватдегидрогеназный комплекс».На I стадии этого процесса пируват (рис. 10.8) теряет свою карбоксильную группу в результате взаимодействия с тиаминпирофосфатом (ТПФ) в составе активного центра фермента пируватдегидрогеназы (E1). На II стадии оксиэтильная группа комплекса E1–ТПФ–СНОН–СН3 окисляется с образованием ацетильной группы, которая одновременно переносится на амид липоевой кислоты (кофермент), связанной с ферментом дигидроли-поилацетилтрансферазой (Е2). Этот фермент катализирует III стадию – перенос ацетильной группы на коэнзим КоА (HS-KoA) с образованием конечного продукта ацетил-КоА, который является высокоэнергетическим (макроэргическим) соединением.
На IV стадии регенерируется окисленная форма липоамида из восстановленного комплекса дигидролипоамид–Е2. При участии фермента дигидролипоилдегидрогеназы (Е3) осуществляется перенос атомов водорода от восстановленных сульфгидрильных групп дигидролипоамида на ФАД, который выполняет роль простетической группы данного фермента и прочно с ним связан. На V стадии восстановленный ФАДН2 дигидро-липоилдегидрогеназы передает водород на кофермент НАД с образованием НАДН + Н+.Процесс окислительного декарбоксилирования пирувата происходит в матриксе митохондрий. В нем принимают участие (в составе сложного мультиферментного комплекса) 3 фермента (пируватдегидрогеназа,
дигидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа) и 5 кофер-ментов (ТПФ, амид липоевой кислоты, коэнзим А, ФАД и НАД), из которых три относительно прочно связаны с ферментами (ТПФ-E1, ли-поамид-Е2 и ФАД-Е3), а два – легко диссоциируют (HS-KoA и НАД).

Е1 - пируватдегидрогеназа; Е2 - ди-гидролипоилацетилтрансфсраза; Е3 -дигидролипоилдегидрогеназа; цифры в кружках обозначают стадии процесса.
Все эти ферменты, имеющие субъединичное строение, и коферменты организованы в единый комплекс. Поэтому промежуточные продукты способны быстро взаимодействовать друг с другом. Показано, что составляющие комплекс полипептидные цепи субъединиц дигидролипоил-ацетилтрансферазы составляют как бы ядро комплекса, вокруг которого расположены пируватдегидрогеназа и дигидролипоилдегидрогеназа. Принято считать, что нативный ферментный комплекс образуется путем самосборки.
Суммарную реакцию, катализируемую пируватдегидрогеназным комплексом, можно представить следующим образом:
Пируват + НАД+ + HS-KoA –> Ацетил-КоА + НАДН + Н+ + СO2.
Реакция сопровождается значительным уменьшением стандартной свободной энергии и практически необратима.Образовавшийся в процессе окислительного декарбоксилирования аце-тил-КоА подвергается дальнейшему окислению с образованием СО2 и Н2О. Полное окисление ацетил-КоА происходит в цикле трикарбоновых кислот (цикл Кребса). Этот процесс, так же как окислительное декарбо-ксилирование пирувата, происходит в митохондриях клеток.
При окислении глюкозы, глицерина и аминокислоты аланина образуется пировиноградная кислота (пируват). Пируват далее может быть окислен до СО2 в общем пути катаболизма. Реакции промежуточн 727d32eh ого обмена протекают в цитоплазме, а окисление пирувата - в митохондриях, поэтому пируват транспортируется в митохондрии специальным переносчиком через мембрану. Первый шаг катаболизма пирувата - окислительное декарбоксилирование.
В этом процессе участвуют ферменты, работающие в определенной последовательности и объединенные в мультиферментный пируватдегидрогеназный комплекс (ПДГК).
В состав этого комплекса входит 3 фермента и 5 коферментов.
Ферменты:
Пируватдегидрогеназа (Е1)
Дигидролипоилацетилтрансфераза (Е2)
Дигидролипоилдегидрогеназа (Е3).
Коферменты:
Тиаминдифосфат (ТДФ) - производное витамина В1
Никотинамидадениндинуклеотид (НАД+) - производное витамина никотинамида РР
Флавинадениндинуклеотид (ФАД) - производное витамина В2
Кофермент А (КоА-SH)
9) амфиболические; метаболиты этих процессов могут превращаться как в катаболических,
так и в анаболических процессах. Примером является цикл трикарбоновых кислот.
ЦИКЛ КРЕБСА
Это высокоорганизованная циклическая система взаимопревращений ди- и трикарбоновых кислот, катализируемых мультиферментным комплексом Он составляет основу клеточного метаболизма Несмотря на то, что этот метаболический путь является замкнутым его началом считается цитратсинтазная реакция в ходе которой конденсация Ацетия-КоА и оксалоацитата дает цитрат Далее следует реакция отщепление воды катализируемая ферментом аконитазой продуктом реакции является цис- Этот же фермент (аконитаза или аконитатгитротаза) катализирует реакцию гидротации в итоге образуется изомер -изолимонная кислота Окислительная реакция кот катализируется ферментом изоцитратдегидрогиназа (она еще и карбоксилирующая) а-кетоггаотаровая кислота В ходе реакции отщепляется СО2, энергия окислительного превращения аккумулируется в восстановленном НАД а-кетоглютаровая кислота под действием а-кетоглюторатдегидрогиназного комплекса (3 типа ферментов, 2 кофермента НАД, КоА и 3 простетических группы тиаминпирофосфат, липоевую кислоту и Флавинадениндннуклеотид) Это тоже реакция окисления идущая путем дегидрирования В данном случае часть энергии запасается еще и в макроэргической связи Образуется сукценил-КоА Сукцешга-КоА Фермент катализирует очень интересную реакцию в ходе которой из ГДФ и фосфорной кислоты образуется ГТФ Отщепляется КоА фермент - сукценил-КоА-синтаза (сукцинаттиокиназа) В итоге образуется янтарная кислота - сукцинат Эта реакция субстратно-окислительного фосфорноирования Сукцинат далее вступает вновь в реакцию окисления с участием фермент» сукцинатдегидрогиназы Это ФАД зависимый фермент остальные дегидрогиназы НАД зависимые ФАД восстанавливается а сукцинат окисляется с образованием фумаровой кислоты Происходит немедленное присоединение воды с участием фермента фумаразы и образуется малат (яблочная кислота) Малат, с участием малатдегидрогиназы содержащий НАД, окисляется в итоге образуется ЩУК т е происходит регенерация первого продукта ЩУК может снова вступать в реакцию конденсации с ацетил-КоА с образованием лимонной
Что происходит в цикле Кребса? СНЗ-С + ЗНАД + ФАД + ГДФ + НЗРО4 + 2Н2О -> 2СОг + ЗНАДН+Н* + ФАДН2 + ГТФ + HSKoA
Как регулируется цикл Кребса? Термодинамический контроль работы цикла
Пусковой реакцией цикла является цитратсиктазная реакция она сопровождается большой потерей энергии Д G = - 7,7 ккал/моль В условиях клетки эта реакция практически необратима Большой потерей энергии сопровождается <х-кетоглюторатдегидрогиназная реакция ДО = -8 ккал/моль В условиях клетки эта реакция абсолютна необратима За счет пусковой стадии и особенно а-кетст люторатдегидрогиназной реакции движение метаболитов возможно лишь в одном направлении т е в направлении деградации (ращипления) ацетип-КоА т е здесь мы имеем дело с двойным термодинамическим контролем направления процесса Кинетический контроль скорости потока метаболитов. Обеспеченность клетки энергией можно характеризовать величиной энергетического ее заряда Он равен отношению концентрации АТФ и половины концентрации АДФ к концентрация АДФ, АТФ и АМФ Когда клетка хорошо обеспечена энергией большая часть ее адениловых нуклеотидов находится в форме АТФ и величина энергетического заряда приближается к 1 Высокая концентрация АДФ и АМФ признак нехватки энергии Отсюда работа цикла Кребса должна тормозиться высокой концентрацией АТФ Цикл Кребса должен стимулироваться высокой концентрацией АДФ и АМФ, энергии не хватает - пора включать цикл Аналогичную роль может играть отношение восстановленно! о НАД к окисленному Скорость работы цикла контролируется прежде всего на пусковой стадии, которая определяется в значительной мере концентрацией ЩУК в клетке Кроме того цитратсинтаза аллостерически угентается высокими концентрациями АТФ, восстав НАД, и сукцинил-КоА Угнетение активности ферментов высокими концентрациями АТФ вполне понятно - клетка обеспеченна энергией и цикл Кребса выключается Второе регуляторное звено - изоцитратдегидрогиназная реакция Изоцитратдегндрогиназа аллостерически ингибируется высокими концентрациями АТФ и восстановленного НАДВ то же время АДФ выступает в качестве аллостерического активатора Аллостерическим активатором по некоторым данным является так же изоцитрат
Третьим уровнем регуляции является а-кетоглюторатдегидропшазная реакция Здесь восстановленный НАД и АТФ явл аллостер ингибиторами работы этого комплекса Активатором выступает АМФ Высокие концентрации сукцинил-КоА угнетает работу комплекса
Активность фермента сукцинатдегидрогиназы ингибируется по конкурентному механизму высокими концентрациями ЩУК и малоновой кислоты.В матриксе митохондрий. В энергетическом обмене играет роль сопров.выходом вост.метаболитом(НАД как вост.НАДН,ФАД
передают……3мол.НАД и 1мол.ФАД.
10) Суммарное уравнение цикла трикарбоновых кислот:
СН3СО~SСоА + 3НАД+ + ФАД + ГДФ + Н3РО4 + 2Н2О ® 2СО2 + 3НАДН + 2Н+ + ФАДН2 + ГТФ + НSСоА.

образование янтарной кислоты из сукцинил-КоA, катализируемое сукцинилтиокиназой, в результате которого энергия, освобождающаяся при разрыве тиоэфирной связи, запасается в фосфатной связи ГТФ . ГТФ затем отдает свою фосфатную группу молекуле АДФ , что приводит к образованию АТФ . Следовательно, на данном этапе ЦТК имеет место субстратное фосфорилирование .
Необратимые=1,цис-аконитат-изоцетрат до сукцината(5)
11) Пируваты (соли пировиноградной кислоты) — важные химические соединения в биохимии. Они является конечным продуктом метаболизма глюкозы в процессе гликолиза. Одна молекула глюкозы превращается при этом в две молекулы пировиноградной кислоты. Дальнейший метаболизм пировиноградной кислоты возможен двумя путями — аэробным и анаэробным.
В условиях достаточного поступления кислорода, пировиноградная кислота превращается в ацетил-кофермент А, являющийся основным субстратом для серии реакций, известных как цикл Кребса, или дыхательный цикл, цикл трикарбоновых кислот. Пируват также может быть превращён в анаплеротической реакции в оксалоацетат. Оксалоацетат затем окисляется до углекислого газа и воды. Эти реакции названы по имени Ханса Адольфа Кребса, биохимика, получившего вместе с Фрицем Липманном Нобелевскую премию по физиологии в 1953 году за исследования биохимических процессов клетки. Цикл Кребса называют также циклом лимонной кислоты, поскольку лимонная кислота является одним из промежуточных продуктов цепи реакций цикла Кребса.Если кислорода недостаточно, пировиноградная кислота подвергается анаэробному расщеплению с образованием молочной кислоты у животных и этанола у растений.При анаэробном дыхании в клетках пируват, полученный при гликолизе, преобразуется в лактат при помощи фермента лактатдегидрогеназы и NADP в процессе лактатной ферментации, либо в ацетальдегид и затем в этанол в процессе алкогольной ферментации.Пировиноградная кислота является «точкой пересечения» многих метаболических путей. Пируват может быть превращён обратно в глюкозу в процессе глюконеогенеза, или в жирные кислоты или энергию через ацетил-КоА, в аминокислоту аланин, или в этанол. Таким образом пируват объединяет несколько ключевых метаболических процессов клетки и является универсальной «метаболической валютой» живой клетки.
12) Анаэробный гликолиз – сложный ферментативный процесс распада глюкозы, протекающий в тканях человека и животных без потребления кислорода. Конечным продуктом гликолиза является молочная кислота. В процессе гликолиза образуется АТФ. Суммарное уравнение гликолиза можно представить следующим образом:
![]()
В анаэробных условиях гликолиз – единственный процесс в животном организме, поставляющий энергию. Именно благодаря гликолизу организм человека и животных определенный период может осуществлять ряд физиологических функций в условиях недостаточности кислорода. В тех случаях, когда гликолиз протекает в присутствии кислорода, говорят об аэробном гликолизе.
Анаэробный гликолиз –брожение
Процесс преобразования глюкозы С6Н12О6 без участия кислорода О2 с образованием пировиноградной кислоты С3Н4О3 и 2 молекул АТФ.
С6Н12О6 + 2АДФ + 2 Н3РО4 + 2НАД = 2С3Н4О3 + 2НАД·2Н2 + 2АТФ + 2Н2О
Процесс анаэробного гликолиза протекает в цитоплазме вне митохондрий.
Аэробный гликолиз –дыхание
Процесс преобразования пировиноградной кислоты С3Н4О3 при участия кислорода с образованием 36 молекул АТФ.
Аэробный гликолиз протекает на мембранах крист митохондрий с обязательным участием комплекса ферментов.
2С3Н4О3 + 6О2 + 36АДФ + 36Н3РО4 + 2НАДФ·Н2 = 6СО2 + 42Н2О + 36АТФ+2НАД
13)Окисление биологиче¬ское,совокупность реакций окисления, протекающих во всех живых клетках. Основная функция О. б. — обеспечение организма энергией в доступной для использования форме. Реакции О. б. в клетках катализируют ферменты, объе-диняемые в класс оксидоредуктаз. О. б. в клетках связано с передачей т. н. вос¬станавливающих эквивалентов (ВЭ) — атомов водорода или электронов — от одного соединения — донора, к дру¬гому — акцептору. У аэробов — боль¬шинства животных, растений и многих микроорганизмов — конечным акцеп¬тором ВЭ служит кислород. Поставщи¬ками ВЭ могут быть как органические, так и неорганические вещества (см. таблицу). Основной путь использования энергии, освобождающейся при О. б., — накопление её в молекулах адено¬зинтрифосфорной кислоты (АТФ) и др. макроэргических соединений.О. б., со-провождающееся синтезом АТФ из аденозиндифосфорной кислоты (АДФ) и неорганического фосфата, происходит при гликолизе, окислении a-кетоглута¬ровой кислоты и при переносе ВЭ в цепи окислительных (дыхательных)ферментов, обычно называют окисли¬тельным фосфорилированием (см. схему). В процессе дыхания углеводы, жиры и белки подвергаются многосту¬пенчатому окислению, которое приво¬дит к восстановлению основных по¬ставщиков ВЭ для дыхательных флави¬нов, никотинамидадениндинуклеотида (НАД), никотинамидадениндинуклео¬тидфосфата (НАДФ) и липоевой ки¬слоты. Восстановление этих соедине¬ний в значительной мере осуществля¬ется втрикарбоновых кислот цикле, которым завершаются основные пути окислительного расщепления углеводов (оно начинается с гликолиза), жиров и аминокислот. Помимо цикла трикарбо¬новых кислот, некоторое количество восстановленных коферментов — ФАД (флавинадениндинуклеотида) и НАД — образуется при окислении жирных ки¬слот, а также при окислительном деза¬минировании глутаминовой кислоты (НАД) и в пентозофосфатном цикле (восстановленный НАДФ). Одновре¬менно развивалось направление, где в основу классификации ферментов был положен тип реакции, подвергающейся каталитическому воздейсвию. Наряду с ферментами, ускоряющими реакции гидролиза (гидролазы), были изучены ферменты, участвующие в реакциях переноса атомов и атомных групп (фе¬разы), в изомеризации (изомеразы), расщеплении (лиазы), различных синте¬зах (синтетазы) и т. д. Это направление в классификации ферментов оказалось наибо-лее плодотворным, так как объе-диняло ферменты в группы не по наду¬манным, формальным признакам, а по типу важнейших биохимических про¬цессов, лежащих в основе жизнедея¬тельности любого организма. По этому принципу все ферменты делят на 6 классов.1. Оксидоредуктазы — уско¬ряют реакции окисления — восстанов-ления. 2. Трансферазы — ускоряют ре¬акции переноса функцио-нальных групп и молекулярных остатков. 3. Гидролазы — уско-ряют реакции гидролитического распада. 4. Лиазы — ускоряют негидро¬литическое отщепление от субстратов определенных групп атомов с образова¬нием двойной связи (или присоединяют группы атомов по двойной связи). 5. Изомеразы — ускоряют пространствен¬ные или структурные перестройки в пределах одной молекулы. 6. Лигазы — ускоряют реакции синтеза, сопряжен¬ные с распадом богатых энергией свя¬зей.
ФУНКЦИИ БИОЛОГИЧЕСКОГО ОКИСЛЕНИЯ
1 Важнейшей функцией биологического окисления является несомненно высвобождение энергии которая в дальнейшем используется в эндоорганических процессах.2 В ходе окисления питательных веществ образуется ряд низкомолекулярных соединений, которые клетка использует потом для биосинтеза Назыв - пластическая функция Например синтез аминокислот из продуктов окисления глюкозы или жиров используется в биосинтетических реакциях Генерация восстановительных эквивалентов (потенциалов).4.Окислительные процессы несут защитную роль Многие ксенобиотики обезвреживаются путем окисления в том числе многие лекарственные препараты 5. Огромная роль в поддержании температуры тела. Таким образом существование живых существ невозможно без окислительных процессов.
фосфорилирование окислительноесинтез молекул АТФ из АДФ и неорганического фосфата за счет энергии окисления молекул органических и неорганических веществ. Ф. о. сопряжено с переносом электронов по дыхательной цепи, элементы которой встроены у эукариот во внутреннюю мембрану митохондрий, а у прокариот монтируются на внутрицитоплазматических мембранах. Перенос электронов по дыхательной цепи митохондрий завершается восстановлением О2, синтез АТФ происходит в трех пунктах энергетического сопряжения. Субстратное фосфорилирование (биохимическое)-синтез богатых энергией фосфорных соединений за счёт энергии окислительно-восстановительных реакций Гликолиза (катализируемых фосфоглицеральдегиддегидрогеназой и енолазой) и при окислении α-кетоглутаровой кислоты в Трикарбоновых кислот цикле (под действием α-кетоглутаратдегидрогеназы и сукцинаттиокиназы). Для бактерий описаны случаи С. ф. при окислении пировиноградной кислоты. С. ф., в отличие от фосфорилирования в цепи переноса электронов , не ингибируется «разобщающими» ядами (например, динитрофенолом) и не связано с фиксацией ферментов в мембранах митохондрий. Вклад С. ф. в клеточный фонд АТФ в аэробных условиях значительно меньше, чем вклад фосфорилирования в цепи переноса электронов.
14)Свободное окисление, не сопряженное с фосфорилированием АДФ, не сопровождающееся трансформацией энергии, выделяющейся при окислении, в энергию макроэргических связей. При свободном окислении высвобождающаяся при сопряженном с окислением распаде химических связей энергия переходит в тепловую и рассеивается.По типу свободного окисления идут все без исключения оксигеназные реакции, все окислительные реакции, ускоряемые пероксидазами или сопровождающиеся образованием Н202, многие реакции, катализируемые оксидазами.
Процессы свободного окисления сосредоточены в цитозоле, в мембранах эндоплазматической сети клетки, в мембранах лизосом, пероксисом и аппарата Гольджи, на внешних мембранах митохондрий и хлоропластов. Они идут также в ядерном аппарате клетки.
Реакции свободного окисления органических соединений в живой природе, равно как и ферментные системы, ускоряющие их, многообразны, и многие из них рассмотрены в предьщущих главах и в начальном разделе этой главы. Этим путем непосредственно окисляются не только многочисленные природные и неприродные субстраты, но и восстановленные коферменты (НАДН, НАДФН, ФАД • Н2 и др.), образовавшиеся при действии первичных и вторичных дегидрогеназ.Хотя реакции свободного окисления идут и в цитозоле, и на мембранах различных субклеточных структур, средоточием их являются мембраны эндоплазматической сети клетки. Так как последние при гомогенизации клеток и фракционировании субклеточных частиц гомогената дают фракцию микро-сом, которая может быть получена в виде препарата, то сейчас активно изучаются организация и функции микросомальной дыхательной цепи. Ее первая особенность сводится к тому, что, несмотря на наличие ферментов цепи переноса электронов, ни в одном пункте этой цепи не происходит сопряжения с фосфорилированием АДФ. Вторая особенность заключается в своеобразии структуры и функциональной активности цитохромов Ъ 5 и Р-450), входящих в ее состав. В частности, цитохром Р-450 (А/«50000, гемопротеин, первичная структура более десятка его форм расшифрована) обладает множеством (сотни, а может быть, и тысячи) форм, возникающих в ответ на введение (или попадание) в организм того или иного класса ксенобиотиков, подобно тому, как антитела синтезируются в ответ на присутствие антигенов; поэтому . цитохром Р-450 считают своего рода «мембранным иммуноглобулином».
Наконец, третья особенность состоит в высоком сродстве терминальной оксидазы микросомальных цепей к кислороду, позволяющая ей конкурировать за кислород с митохондриальной цитохромоксидазой. Поэтому, например, в клетках печени доля микросомального окисления эндогенных субстратов составляет 40%, а митохондриального—60%.
Наиболее своеобразными и почти не затронутыми в предыдущих главах являются реакции свободного окисления, идущие при участии оксигеназ. Этотподкласс оксидоредуктаз содержит ферменты, ускоряющие включение в окисляемый субстрат либо двух (диокситеназы), либо одного (монооксигеназы) атома молекулярного кислорода.Свободное окисление при участии диоксигеназ. Одной из наиболее изученных диоксигеназ является пирокатехаза (катехол: кислород-1,2-оксидоредуктаза дециклизующая); ее М = 85000. Концентрированные растворы пирокатехазы красного цвета, так как она содержит в активном центре два прочно связанных атома Fe, которые, согласно О. Хайаиши, соединяются с молекулярным кислородом в комплекс, где кислород далее активируется: Fe2+-f-02-*Fe2+02-» -Fe3+02.атем в активном центре пирокатехазы после присоединения субстрата возникает тройной комплекс, преобразование которого приводит к включению молекулярного кислорода в пирокатехин:
Пирокатехин Муконовая кислота
Аналогично действует катехол-2,3-оксйгеназа (М = 140 ООО, 1 атом Fe), 3,4-диоксигеназа протокатеховой кислоты (М = 700 ООО, 8x90000), образующая р-карбоксимуконовую кислоту, триптофаноксигеназа (М = 1231000), присоединяющая молекулярный кислород по пиррольному кольцу индолильного радикала, и другие диоксигеназы. Во всех случаях молекулярный кислород активируется за счет присоединения электрона, теряемого двухвалентным железом активного центра, а возникший анионный свободный радикал кислорода (02~) атакует и оксигенирует субстрат. В результате расшифровки механизма действия диоксигеназ стал ясен один из способов активирования молекулярного кислорода терминальными оксидазами.
Одной из биологически важных диоксигеназных реакций является превращение Р-каротина в витамин А:
Ретиналь (витамин А)
Прямое присоединение 1802 по 15,15'-связи в р-каротине впервые доказано Б. Б. Вартапетяном и сотр. (1966).Свободное окисление при участии монооксигеназ. Ввиду высокой лабильности монооксигеназ, как, впрочем, и диоксигеназ, выделение их сопряжено с большими трудностями. Тем не менее некоторые из них получены в кристал-лическом состоянии. Монооксигеназы характеризуются молекулярными массами от 65000 до 200000, отсутствием, как правило, в их составе ионов тяжелых металлов и наличием коферментов флавиновой природы. Кроме того, многие из них требуют участия в реакции монооксигенирования так называемого парного донора, от которого поступают атомы водорода на один из атомов молекулярного кислорода (второй внедряется в окисляемый субстрат).
Простейший представитель монооксигеназ—фенолгидроксилаза (фенол-2-монооксигеназа):
НАДФ+ + н,о
Фешм Пнрсптеош
0гон+°>+нлдфн+н* ОС
Монооксигеназы принимают большое участие в окислении аминокислот (лизин-, аргинин- и триптофанмонооксигеназы, фенил аланин- и тирозингид-роксилаза), оксикислот (салицилатгидроксилаза), полиизопреноидных соединений (сквален-эпоксидаза, см. с. 404).
Механизм действия монооксигеназ выяснен недостаточно. Предполагают, что активной формой кислорода может быть связанный с ферментом перок-сид водорода или его эквивалент. Между тем показано, что некоторые из монооксигеназ (фенолаза из грибов, фёниланин-4-гидроксилаза) содержат Си+ в своем составе. В этих случаях возможен такой механизм активирования кислорода:
Монооксигенаэа
'Си"1
'Си"1
+ субстрат
гидроксилированный
+ НАД+ + Н20
Здесь, как и в случае диоксигеназ, решающую роль в активировании молекулярного кислорода играет передача на него электронов с металла, входящего в состав фермента. Не исключено, что этот способ вовлечения молекулярного кислорода в процессы оксидоредукции в клетке является общим для оксидаз, участвующих как в свободном, так и в сопряженном с фосфорилированием биологическом окислении.
Главным источником АФК в клетках являются митохондрии. Обычно примерно 98% всего кислорода, поступающего в клетки, используется для окисления субстратов с образованием АТФ и выделением тепла, и лишь 2% используется в реакциях образования АФК, которое может значительно возрастать при усиленном поступлении кислорода в клетки или нарушении работы электронно–транспортной цепи митохондрий.
АКТИВНЫЕ ФОРМЫ КИСЛОРОДА (АФК)
О2° - супероксидный анион радикал
НО2° – гидропероксидный радикал
НО° – гидроксил-радикал
Н2О2 – пероксид водорода
НОСl – гипохлорная кислота
Н2О2 и НОСl генерируют свободные радикалы в присутствии Fe2+ и О2°:
Н2О2 + Fe2+ → НО° + ОН° + Fe3+ (реакция Фентон)
Н2О2 + О2° → НО° + ОН° + О2 (реакция Хабера-Вайса)
НОСl + О2° → НО° + СIО2
В свою очередь, О2° способен превращаться в НО2° и Н2О2 согласно реакциям:
О2° + Н+ → НО2°
НО2° + НО2° → Н2О2 + О2
Кроме того, к АФК относят окись азота (NO°), образующую с О2° пероксинитрит:
NO° + О2° → ONOO° => ONOOH >НО° + NO2°
Образование активных форм кислорода может происходить:
в процессе переноса электронов в митохондриальной дыхательной цепи;
в реакциях, которые катализируются оксидазами (образуется перекись водорода), в том числе в свободнорадикальных процессах, совершающихся в фагоцитах;
в реакциях микросомального окисления при обезвреживании веществ с участием цитохрома Р-450;
в реакциях самопроизвольного (неферментативного) окисления веществ (гемоглобина, ферредоксинов, адреналина и др.);
в биологических системах с наличием ионов металлов с переменной валентностью и, прежде всего, железа (свободных атомов, так называемых внегемовых)
15) Организация и функционирование дыхательной цепи. В клетках эукариот дыхательная цепь расположена во внутренней мембране митохондрий, у дышащих бактерий – в цитоплазматической мембране и специализированных структурах – мезосомах, или тилакоидах. Компоненты дыхательной цепи митохондрий в порядке убывания окислительно-восстановительного потенциала можно расположить, как показано.

Молярные соотношения компонентов дыхательной цепи являются постоянными, ее компоненты встроены в митохондриальную мембрану в виде 4 белково-липидных комплексов: НАДН-КоQН2-редуктаза (комплекс I), сукцинат-КоQ-редуктаза (комплекс II), КоQН2-цитохром c-редуктаза (комплекс III) и цитохром а-цитохромокси-даза (комплекс IV).
Если субстратом окисления служат α-кетокислоты, в переносе электронов на НАД+ участвуют липоатсодержащие дегидрогеназы. В случае окисления пролина, глутамата, изоцитрата и других субстратов перенос электронов происходит непосредственно на НАД+. Восстановленный НАД в дыхательной цепи окисляется НАДН-дегидрогеназой, содержащей железосерный белок (FeS) и ФМН и прочно связанной с дыхательной цепью.KoQ (убихинон), необходимый компонент дыхательной цепи, является производным бензохинона с боковой цепью, которая у млекопитающих чаще всего представлена 10 изопреноидными единицами (см. главу 7). Как любой хинон, KoQ способен находиться и в восстановленном, и окисленном состоянии. Это свойство определяет его роль в дыхательной цепи - служить коллектором восстановительных эквивалентов, поставляемых в дыхательную цепь через флавиновые дегидрогеназы. Содержание его значительно превосходит содержание других компонентов дыхательной цепи.Дополнительным участником дыхательной цепи является железосерный белок FeS (негемовое железо). Он участвует в окислительно-восстановительном процессе, протекающем по одноэлектронному типу. Первый участок локализации FeS находится между ФМН и KoQ, второй - между цитохромами b и c1. Это соответствует тому факту, что со стадии ФМН путь протонов и электронов разделяется: первые накапливаются в митохондриальном матриксе, а вторые идут на гидрофобные переносчики - KoQ и цитохромы.Цитохромы в дыхательной цепи выстроены в порядке возрастания окислительно-восстановительного потенциала. Они представляют собой гемопротеины, в которых простетическая геминовая группа близка к гему гемоглобина (у цитохрома b идентична). Ионы железа в составе гема при получении и отдаче электронов обратимо изменяют свою валентность.В процессах тканевого дыхания наиболее важную роль играют цитохро-мы b, с1, с, а и а3. Цитохром а3 представляет собой терминальный участок дыхательной цепи – цитохромоксидазу, которая осуществляет окисление цитохрома с и образование воды. Элементарный акт представляет собой двухэлектронное восстановление одного атома кислорода, т.е. каждая молекула кислорода одновременно взаимодействует с двумя электрон-транспортными цепями. При транспорте каждой пары электронов во внутримитохондриальном пространстве может накапливаться до 6 протонов .Строение дыхательной цепи интенсивно исследуется. В числе последних достижений молекулярной биохимии – установление тонкой структуры дыхательных ферментов с помощью рентгеноструктурного анализа. С помощью электронного микроскопа с наивысшим доступным в настоящее время разрешением можно «увидеть» структуру цитохромоксидазы.Если акцепторы электронов обр.при окисл.органич.субстр.явл.орган.соед,то анаэробный процесс,если конечн.акцептр электр явл.О2,то аэробный процесс.
16)Ферменты цикла Кребса, кроме СДГ, локализованной во внутренней митохондриальной мембране, находятся во внутримитохондриальном матриксе. В цитоплазме представлена активность отдельных энзимов цикла, но там они участвуют в иных метаболических реакциях. Кроме цикла Кребса, работа митохондрий подразумевает окислительное фосфорилирование (см. также т.1, стр. 168174 и данный том выше, раздел «Энергетический метаболизм и его нарушения»). Ниже даётся лишь краткое рабочее описание этого заключительного этапа метаболического пути взятых из углеводов (или других энергетических эквивалентов) электронов
Последовательность реакций переноса водорода и электронов (дыхательная цепь) локализована на внутренней стороне внутренней мембраны митохондрий. Эта цепь включает:
НАДН — дегидрогеназы
ФАДН — дегидрогеназы (флавопротеиды)
негемовые железосодержащие белки
кофермент Q
цитохромы в, си с, а, а3
и, наконец, финальный акцептор электронов — кислород.
Легко заметить, что дыхательная цепь зависит от функционирования нескольких витаминов и микроэлементов, а гиповитаминозы и дефицит железа и других металлов в организме будут оборачиваться тканевой гипоксией (см. ниже разделы «Патофизиология витаминного обмена», «Патофизиология обмена микроэлементов»).Окислительное фосфорилирование — процесс, при котором выделение энергии Насчет окисления субстратов сопряженно с синтезом АТФ.Первым пунктом сопряжения является участок, где два электрона и два водорода от восстановленного НАД переносятся на флавопротеиды.Восстановленная форма цитохрома в действует в качестве восстановителя гемацитохромов с, что представляет собой второй пункт сопряжения окисления и фосфорилирования.Третьим пунктом сопряжения является заключительный участок дыхательной цепи, осуществляющий передачу электронов от цитохрома а3на кислород.
Механизм сопряжения окисления и фосфорилирования
Каким же образом осуществляется сопряжение этих двух процессов? Наиболее обоснованный ответ на этот вопрос даёт хемиосмотическая теория Митчелла, предложенная им в 1961 г. Основные положения были подтверждены и разработаны детально совместными усилиями многих исследователей в последующие годы.
1. Протонный градиент и электрохимический потенциал
Перенос электронов по дыхательной цепи от NADH к кислороду сопровождается выкачиванием протонов из матрикса митохондрий через внутреннюю мембрану в межмембранное пространство. На эту работу затрачивается часть энергии электронов, переносимых по ЦПЭ.
Протоны, перенесённые из матрикса в межмембранное пространство, не могут вернуться обратно в матрикс, так как внутренняя мембрана непроницаема для протонов. Таким образом, создаётся протонный градиент, при котором концентрация протонов в межмембранном пространстве больше, а рН меньше, чем в матриксе. Кроме того, каждый протон несёт положительный заряд, и вследствие этого появляется разность потенциалов по обе стороны мембраны: отрицательный заряд на внутренней стороне и положительный - на внешней. В совокупности электрический и концентрационный градиенты составляют электрохимический потенциал ΔμН+ - источник энергии для синтеза АТФ. Так как наиболее активный транспорт протонов в межмембранное пространство, необходимый для образования ΔμН+, происходит на участках ЦПЭ, соответствующих расположению комплексов I, III и IV, эти участки называют пунктами сопряжения дыхания и фосфорилирования .Механизм транспорта протонов через мито-хондриальную мембрану в пунктах сопряжения недостаточно ясен. Однако установлено, что важную роль в этом процессе играет KoQ. Наиболее детально механизм переноса протонов при участии KoQ изучен на уровне комплекса III .KoQ переносит электроны от комплекса I к комплексу III и протоны из матрикса в межмембранное пространство, совершая своеобразные циклические превращения, называемые Q-циклами. Донором электронов для комплекса III служит восстановленный убихинон (QH2), а акцептором - цитохром с. Цитохром с находится с внешней стороны внутренней мембраны митохондрий; там же располагается активный центр цитохрома с1 с которого электроны переносятся на цитохром с.В мембране существует стационарный общий фонд Q/QH2, из которого каждая молекула QH2 в одном цикле обеспечивает перенос протонов из матрикса в межмембранное пространство и электронов, которые в конечном итоге поступают на кислород. На работу, совершаемую при выкачивании протонов, расходуется часть свободной энергии, которая освобождается при переносе электронов по градиенту редокс-потенциала. Энергия электрохимического потенциала (∆μH+) используется для синтеза АТФ, если протоны возвращаются в матрикс через ионные каналы АТФ-синтазы.
2. Строение АТФ-синтазы и синтез АТФ
АТФ-синтаза (Н+-АТФ-аза) - интегральный белок внутренней мембраны митохондрий. Он расположен в непосредственной близости к дыхательной цепи. АТФ-синтаза состоит из 2 белковых комплексов, обозначаемых как F0 и F1.Гидрофобный комплекс F0 погружён в мембрану. Он служит основанием, которое фиксирует АТФ-синтазу в мембране. Комплекс F0 состоит из нескольких субъединиц, образующих канал, по которому протоны переносятся в матрикс.Комплекс F1 выступает в митоховдриальный матрикс. Он состоит из 9 субъединиц (Зα, 3β, γ, ε, δ). Субъединицы аир уложены попарно, образуя "головку"; между α- и β-субъединицами располагаются 3 активных центра, в которых происходит синтез АТФ; γ-, ε-, δ- субъединицы связывают комплекс F1 с F0.Повышение концентрации протонов в межмембранном пространстве активирует АТФ-синтазу. Электрохимический потенциал ΔμH+ заставляет протоны двигаться по каналу АТФ-синтазы в матрикс. Параллельно под действием ΔμH+ происходят конформационные изменения в парах α, β-субъединиц белка F1, в результате чего из АДФ и неорганического фосфата образуется АТФ. Электрохимический потенциал, генерируемый в каждом из 3 пунктов сопряжения в ЦПЭ, используют для синтеза одной молекулы АТФ.
3.Коэффициент окислительногофосфорилирования
Окисление молекулы NADH в ЦПЭ сопровождается образованием 3 молекул АТФ; электроны от FAD-зависимых дегидрогеназ поступают в ЦПЭ на KoQ, минуя первый пункт сопряжения. Поэтому образуются только 2 молекулы АТФ. Отношение количества фосфорной кислоты (Р), использованной на фосфорилирование АДФ, к атому кислорода (О), поглощённого в процессе дыхания, называют коэффициентом окислительного фосфорилирования и обозначают Р/О. Следовательно, для NADH Р/О = 3, для сукцината Р/О - 2. Эти величины отражают теоретический максимум синтеза АТФ, фактически эта величина меньше.
4.Дыхательный контроль
Окисление субстратов и фосфорилирование АДФ в митохондриях прочно сопряжены. Скорость использования АТФ регулирует скорость потока электронов в ЦПЭ. Если АТФ не используется и его концентрация в клетках возрастает, то прекращается и поток электронов к кислороду. С другой стороны, расход АТФ и превращение его в АДФ увеличивает окисление субстратов и поглощение кислорода. Зависимость интенсивности дыхания митохондрий от концентрации АДФ называют дыхательным контролем. Механизм дыхательного контроля характеризуется высокой точностью и имеет важное значение, так как в результате его действия скорость синтеза АТФ соответствует потребностям клетки в энергии. Запасов АТФ в клетке не существует. Относительные концентрации АТФ/АДФ в тканях изменяются в узких пределах, в то время как потребление энергии клеткой, т.е. частота оборотов цикла АТФ и АДФ, может меняться в десятки раз.
Общее содержание АТФ в организме 30-50 г, но каждая молекула АТФ в клетке "живёт" меньше минуты. В сутки у человека синтезируется 40-60 кг АТФ и столько же распадается. Увеличение концентрации АДФ немедленно приводит к ускорению дыхания и фосфорилирования.
17)Установлено, что окисление жирных кислот протекает в печени, почках, скелетных и сердечной мышцах, в жировой ткани. В мозговой ткани скорость окисления жирных кислот весьма незначительна; основным источником энергии в мозговой ткани служит глюкоза.
В 1904 г. Ф. Кнооп (F.Knoop) выдвинул гипотезу Р-окисления жирных кислот на основании опытов по скармливанию собакам различных жирных кислот, в которых один атом водорода в концевой метильной группе (ш-углеродного атома) был замещен радикалом (С6Н5—).
Ф. Кнооп высказал предположение, что окисление молекулы жирной кислоты в тканях организма происходит в Р-положении. В результате от молекулы жирной кислоты последовательно отщепляются двууглеродные фрагменты со стороны карбоксильной группы.
Жирные кислоты, входящие в состав естественных жиров животных и растений, имеют четное число углеродных атомов. Любая такая кислота, от которой отщепляется по паре углеродных атомов, в конце концов проходит через стадию масляной кислоты. После очередного Р-окисления масляная кислота становится ацетоуксусной. Последняя затем гидроли-зуется до двух молекул уксусной кислоты. Теория Р-окисления жирных кислот, предложенная Ф. Кноопом, в значительной мере послужила основой современных представлений о механизме окисления жирных кислот.
Доставка жирных кислот к месту их окисления—к митохондриям — происходит сложным путем: при участии альбумина осуществляется транспорт жирной кислоты в клетку; при участии специальных белков (fatty acid binding proteins, FABP) — транспорт в пределах цитозоля; при участии карнитина — транспорт жирной кислоты из цитозоля в митохондрии.
Процесс окисления жирных кислот складывается из следующих основных этапов.
Активация жирных кислот. Свободная жирная кислота независимо от длины углеводородной цепи является метаболически инертной и не может подвергаться никаким биохимическим превращениям, в том числе окислению, пока не будет активирована. Активация жирной кислоты протекает на наружной поверхности мембраны митохондрий при участии АТФ, коэнзима A (HS-KoA) и ионов Mg2+. Реакция катализируется ферментом ацил-КоА-синтетазой:
![]()
В результате реакции образуется ацил-КоА, являющийся активной формой жирной кислоты.
Считают, что активация жирной кислоты протекает в 2 этапа. Сначала жирная кислота реагирует с АТФ с образованием ациладенилата, представляющим собой эфир жирной кислоты и АМФ. Далее сульфгидрильная группа КоА действует на прочно связанный с ферментом ациладенилат с образованием ацил-КоА и АМФ.Транспорт жирных кислот внутрь митохондрий. Коэнзимная форма жирной кислоты, в равной мере как и свободные жирные кислоты, не обладает способностью проникать внутрь митохондрий, где, собственно, и протекает их окисление. Переносчиком активированных жирных кислотРеакция протекает при участии специфического цитоплазматического фермента карнитин-ацилтрансферазы. Уже на той стороне мембраны, которая обращена к матриксу, ацильная группа переносится обратно на КоА, что термодинамически выгодно, поскольку О-ацильная связь в кар-нитине обладает высоким потенциалом переноса группы. Иными словами, после прохождения ацилкарнитина через мембрану митохондрий происходит обратная реакция—расщепление ацилкарнитина при участии HS-KoA и митохондриальной карнитин-ацилтрансферазы:
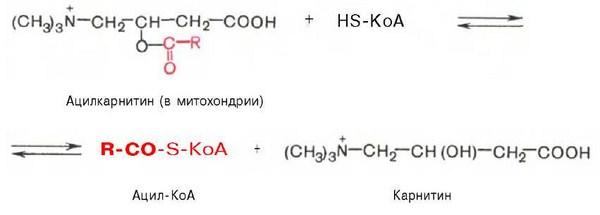
Внутримитохондриальное окисление жирных кислот. Процесс окисления жирной кислоты в митохондриях клетки включает несколько последовательных энзиматических реакций.
Первая стадия дегидрирования. Ацил-КоА в митохондриях прежде всего подвергается ферментативному дегидрированию, при этом ацил-КоА теряет 2 атома водорода в а – и в-положениях, превращаясь в КоА-эфир ненасыщенной кислоты. Таким образом, первой реакцией в каждом цикле распада ацил-КоА является его окисление ацил-КоА-де-гидрогеназой, приводящее к образованию еноил-КоА с двойной связью между С-2 и С-3:
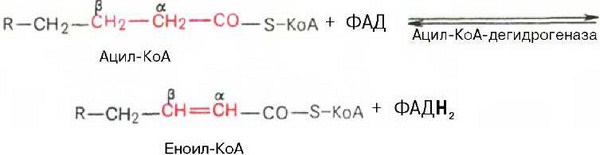
Существует несколько ФАД-содержащих ацил-КоА-дегидрогеназ, каждая из которых обладает специфичностью по отношению к ацил-КоА с определенной длиной углеродной цепи.
Стадия гидратации. Ненасыщенный ацил-КоА (еноил-КоА) при участии фермента еноил-КоА-гидратазы присоединяет молекулу воды. В результате образуется в-оксиацил-КоА (или 3-гидроксиацил-КоА):
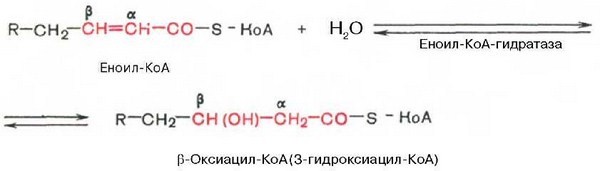
Заметим, что гидратация еноил-КоА стереоспецифична, подобно гидратации фумарата и аконитата (см. с. 348). В результате гидратации транс-А2-двойной связи образуется только L-изомер 3-гидроксиацил-КоА.Вторая стадия дегидрирования. Образовавшийся в-оксиацил-КоА (3-гидроксиацил-КоА) затем дегидрируется. Эту реакцию катализируют НАД+-зависимые дегидрогеназы:
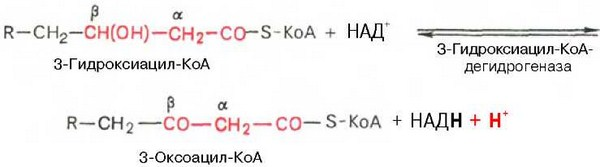
Тиолазная реакция. В ходе предыдущих реакций происходило окисление метиленовой группы при С-3 в оксогруппу. Тиолазная реакция представляет собой расщепление 3-оксоацил-КоА с помощью тиоловой группы второй молекулы КоА. В результате образуется укороченный на два углеродных атома ацил-КоА и двууглеродный фрагмент в виде ацетил-КоА. Данная реакция катализируется ацетил-КоА-ацилтрансферазой (в-ке-тотиолазой):
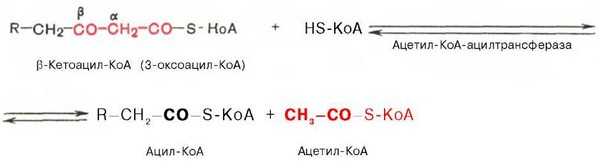
Образовавшийся ацетил-КоА подвергается окислению в цикле трикар-боновых кислот, а ацил-КоА, укоротившийся на два углеродных атома, снова многократно проходит весь путь в-окисления вплоть до образования бутирил-КоА (4-углеродное соединение), который в свою очередь окисляется до 2 молекул ацетил-КоА (рис. 11.2). Например, при окислении пальмитиновой кислоты (С16) повторяется 7 циклов в-окисления. Запомним, что при окислении жирной кислоты, содержащей п углеродных атомов, происходит n/2—1 цикл в-окисления (т. е. на один цикл меньше, чем n/2, так как при окислении бутирил-КоА сразу происходит образование 2 молекул ацетил-КоА) и всего получится п/2 молекул ацетил-КоА.
18) С17Н35СООН + [O] = 18CO2 +18H2O
При метаболизме жир ных кислот выход энергии составляет 8 молекул АТФ на 1 атом углерода. Например, при окислении 1 молекулы стеариновой кислоты образуется 104 атома Н и 148 молекул АТФ.Стеариновая кислота относится к насыщенным жирным кислотам CH3-(CH2)16-СООН
1)активация(Ацил-КоА-синтетаза)
CH3-(CH2)16-СООН+HS-KoA+АТФCH3-(CH2)16-СО-S-КоА+АМФ+ФФн
2. Ацил-S-КоА не способен проходить через митохондриальную мембрану, поэтому существует способ его переноса в комплексе с витаминоподобным веществом карнитином. На наружной мембране митохондрий имеется фермент карнитин-ацилтрансфераза I.
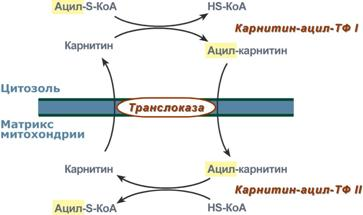
3. После связывания с карнитином жирная кислота переносится через мембрану транслоказой. Здесь на внутренней стороне мембраны фермент карнитин-ацилтрансфераза II вновь образует ацил-S-КоА который вступает на путь β-окисления.
4. Процесс собственно β-окисления состоит из 4-х реакций, повторяющихся циклически. В них последовательно происходит окисление (ацил-SКоА-дегидрогеназа), гидратирование (еноил-SКоА-гидратаза) и вновь окисление 3-го атома углерода (гидроксиацил-SКоА-дегидрогеназа). В последней, трансферазной, реакции от жирной кислоты отщепляется ацетил-SКоА. К оставшейся (укороченной на два углерода) жирной кислоте присоединяется HS-КоА, и она возвращается к первой реакции. Все повторяется до тех пор, пока в последнем цикле не образуются два ацетил-SКоА.
Реакции бета-окисления жирных кислот
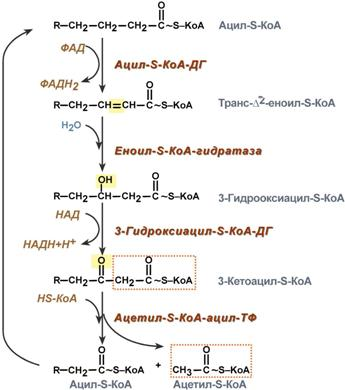
Окисление стеариновой кислоты
так как имеется 18 атомов углерода, то при β-окислении образуется 9 молекул ацетил-SКоА. Последний поступает в ЦТК, при его окислении в одном обороте цикла образуется 3 молекулы НАДН, 1 молекула ФАДН2 и 1 молекула ГТФ, что эквивалентно 12 молекулам АТФ. Итак, 9 молекул ацетил-S-КоА обеспечат образование 9×12=108 молекул АТФ.
для стеариновой кислоты число циклов β-окисления равно 8. В каждом цикле образуется 1 молекула ФАДН2 и 1 молекула НАДН. Поступая в дыхательную цепь, в сумме они "дадут" 5 молекул АТФ. Таким образом, в 8 циклах образуется 8×5=40 молекул АТФ.
двойных связей в стеариновой кислоте нет.
∑148 АТФ
на активацию жирной кислоты идет 1 молекула АТФ, которая, однако, гидролизуется до АМФ, то есть тратятся 2 макроэргические связи или две АТФ.
148-2(или 1 надо уточнить)=146(147)
С17Н35СООН + 26 О2 = 18 СО2 + 18 Н2О.